
Abstract
Background
Drugs that modulate endocannabinoid signalling are effective in reducing nociception in animal models of pain and may be of value in the treatment of migraine.
Methods
We investigated the anti-nociceptive effects of inhibition of monoacylglycerol lipase (MGL), a key enzyme in the hydrolysis of the 2-arachidonoylglycerol, in a rat model of migraine based on nitroglycerin (NTG) administration. We evaluated c-fos expression in specific brain areas and nociceptive behavior in trigeminal and extra-trigeminal body areas.
Results
URB602, a reversible MGL inhibitor, did not show any analgesic effect in the tail flick test, but it inhibited NTG-induced hyperalgesia in both the tail flick test and the formalin test applied to the hind paw or to the orofacial area. Quite unexpectedly, URB602 potentiated formalin-induced hyperalgesia in the trigeminal area when used alone. The latter result was also confirmed using a structurally distinct, irreversible MGL inhibitor, JZL184. URB602 did not induce neuronal activation in the area of interest, but significantly reduced the NTG-induced neuronal activation in the ventrolateral column of the periaqueductal grey and the nucleus trigeminalis caudalis.
Conclusions
These findings support the hypothesis that modulation of the endocannabinoid system may be a valuable approach for the treatment of migraine. The topographically segregated effect of MGL inhibition in trigeminal/extra-trigeminal areas calls for further mechanistic research.
Introduction
Cannabinoid analgesia involves effects at the supraspinal (1,2), spinal (3) and peripheral levels (4,5). Endocannabinoid administration, either systemically or directed at appropriate pain relay or modulatory sites, alters pain sensitivity and modulates the processing of nociceptive information within discrete spinal and brain pathways (6,7). Thus, considerable attention has been devoted to the endocannabinoid system as a potential therapeutic target for pain control (8).
Anandamide (AEA) and 2-arachidonoylglycerol (2-AG) are the most studied endocannabinoids. They act mainly on cannabinoid (CB) 1 and CB2 receptors to modulate normal physiology and disease states, including nociceptive processing (9). The antinociceptive action of endocannabinoids and their role in the modulation of trigeminovascular system activation suggests that the endocannabinoid system may be dysfunctional in migraine patients (10).
The availability of inhibitors of the enzymes that degrade AEA and 2-AG, fatty acid amide hydrolase (FAAH) and monoacylglycerol lipase (MGL) respectively, has provided pharmacological probes for exploring AEA and 2-AG signalling pathways in experimental models of pain. Indeed, the possibility of targeting endocannabinoid-degrading enzymes allows the actions of endocannabinoids to be prolonged (11). Several studies have demonstrated the anti-hyperalgesic effect of MGL inhibition in animal models of inflammation and neuropathic pain (12), but no study has ever specifically addressed the potential role of 2-AG modulation in migraine pain.
Nitroglycerin (NTG) administration causes spontaneous-like migraine attacks in migraine sufferers. In the rat, NTG produces an increased sensitivity to pain and neuronal activation in several brain structures involved in pain transmission (13,14). In a previous study, we have shown that exogenous AEA abolishes NTG-induced nociceptive behavior at the plantar formalin test and significantly decreases NTG-induced c-Fos expression in nucleus trigeminalis caudalis (NTC) and area postrema (15), two areas that are believed to play an important role in migraine pain. AEA modulates NTG-induced TRPV1 and nNOS expression in the C1-C2 segments, and it reduces the activation of the inflammatory pathway in the same areas (16). Additionally, AEA may exert a direct effect upon trigeminal neurons (17) to cause inhibition of CGRP release from central terminals of primary afferent fibers (18,19) and to reduce the nociceptive behavior. Alternatively, AEA might reduce the formation of NO by iNOS in macrophages located in the meninges (20) via CB1 or CB2 receptors activation (21). More recently, we demonstrated that the peripherally restricted FAAH inhibitor, URB937, attenuates NTG-induced hyperalgesia in the plantar and orofacial formalin tests. URB937 also significantly reduces NTG-induced c-Fos expression in the NTC and in the locus coeruleus (LC). Moreover, we found increased MGL activity in specific brain regions of rat following NTG administration, which is suggestive of altered 2-AG-mediated signalling in the CNS (22). To gain insights into the possible role of 2-AG in migraine pathophysiology, in the present study we evaluated the effect of systemic MGL inhibition on NTG-induced neuronal activation and NTG-induced hyperalgesia in a well-established rat model of migraine.
Material and methods
The findings presented in this study reflect experiments that were conducted partly simultaneously with a previous study (23) on URB937, an inhibitor of another enzyme that degrades endocannabinoids. Both URB937 and URB602 are best dissolved in the same vehicle Dimethylsulfoxide (DMSO), therefore we used the same control group wherever possible and reasonable in order to reduce to a minimum the number of rats exposed to painful stimuli.
Animals
Adult Sprague-Dawley rats (weight 175–200 g, Charles River, Calco, Como, Italy) were used. The IASP’s guidelines for pain research in animals were followed (24). Rats were housed in plastic boxes in groups of two with water and food available ad libitum and kept on a 12:12 hours light-dark cycle at the Centralized Animal Facility of the University of Pavia. The protocol (Document N°1032/2015-PR) was approved by the ad hoc Committee of the Ministry of Health at the National Institute of Health (Rome, Italy) and by the Institutional Animal Care and Use Committee.
Drugs
NTG (Bioindustria LIM Novi Ligure (AL), Italy) was prepared from a stock solution of 5.0 mg/1.5 ml dissolved in 27% alcohol and 73% propylene glycol. For the injections, NTG was further diluted in saline (0.9% NaCl) to reach the final concentration of alcohol 6% and propylene glycol (PG) 16%. The vehicle contained saline, alcohol 6% and PG 16% (NTG vehicle). URB602 ([1,1′-biphenyl]-3-yl-carbamic acid, cyclohexyl ester, Cayman Chemical) and JZL184 (4-Nitrophenyl 4-(dibenzo[d][1,3]dioxol-5-yl(hydroxy)methyl)piperidine-1-carboxylate, Cayman Chemical) were dissolved in 100% DMSO (25) and were injected intraperitoneally at a dose of 2 and 4 mg/kg, respectively. DMSO was administered in a volume of 1 ml/kg body weight as reported in our previous study (23). URB602, a reversible MGL inhibitor, was used for all the behavioral tests and fos expression, whereas JZL184, an irreversible MGL inhibitor, was only tested at the orofacial formalin test to confirm the results obtained with URB602. In the experimental session, rats received NTG (10 mg/kg, i.p.) or its vehicle and were treated with URB602 or JZL184 or DMSO three hours later (see experimental groups below).
Timing of evaluations
The behavioral tests and the immunohistochemical evaluations were performed four hours after NTG or its vehicle administration. Animals were assigned to one of the treatment groups according to a randomization list, whose codes were disclosed only after study completion. All researchers who performed the behavioral testing were blind to treatments. Each rat was acclimatized to the test chamber 20 minutes before testing.
Tail flick test
Experimental Groups: NTG + DMSO (n = 6); NTG vehicle + DMSO (n = 7); NTG vehicle + URB602 (n = 6); NTG + URB602 (n = 6).
The test was performed with a test instrument (Ugo Basile, model 7360, Varese, Italy) that allowed automatic recording of tail-flick latency to radiant heat with a temperature of 50℃. Latency at each evaluation was calculated as the mean of three measurements in three different parts of the tail. The movement of the tail from the window of the beam of light to hit a sensor (latency in s) was automatically registered (26). Following the test, each rat was sacrificed with a lethal dose of anesthetic.
Plantar Formalin test
Experimental Groups: NTG+DMSO (n = 6); NTG vehicle + DMSO (n = 7); NTG vehicle + URB602 (n = 6); NTG + URB602 (n = 6).
One animal at a time was placed into a plexiglas observation chamber (10 × 20 × 24 cm) with a mirror (45° angle) positioned to permit unhindered observation of the animal paws. A 100 µl volume of 1% formalin solution was injected subcutaneously into the center of the plantar surface of the right hind paw with slight restraint (27). Each rat was then replaced in the box, the clock was started, and nocifensive responses were recorded for a period of 60 min. Nocifensive behavior was quantified by counting the total number of flinches and shakes occurring for 1 min periods from 1–5 min (Phase I) and then for 1 min periods at 5 min intervals during the period from 15 to 60 min (Phase II) after formalin injection. At the end of the test, each rat was sacrificed with a lethal dose of anesthetic.
Orofacial Formalin test
Experimental Groups: NTG + DMSO (n = 7); NTG vehicle + DMSO (n = 7); NTG vehicle + URB602 (n = 6); NTG + URB602 (n = 7); NTG vehicle + JZL184 (n = 6); NTG + JZL184 (n = 6).
The observation box was a 30 × 30 × 30 cm glass chamber with mirrored sides. A camera, recording animal behaviors for off-line analysis, was located 50 cm from the box to provide a clear view of each rat. The subcutaneous injection of formalin (1.5%, 50 µl) was performed into the right upper lip. Immediately after formalin injection, each animal was placed into the observation box and its behavior recorded for a 45 min period (23). The pain–related behavior was quantified by counting the seconds the animal spent grooming the injected area with the ipsilateral fore- or hind-paw in both Phase I and Phase II of the test. The observation time was divided into 15 blocks of 3 min each for the time course analysis. The face rubbing time was quantified by counting the total seconds of face rubbing recorded during Phase I (0–6 minutes after formalin injection) and during Phase II (12–45 min after formalin injection). Following the test, each rat was sacrificed with a lethal dose of anesthetic.
c-Fos expression
The evaluation of the effect of URB602 on c-fos expression was introduced as an ancillary measure to verify that URB602, via its potentiation effect upon the endocannabinoid system, did not induce per se any metabolic activation in the areas of interest, which would suggest a possible indirect effect (e.g. sedation or interaction with central neurotransmission).
Experimental groups: NTG + DMSO (n = 8); NTG vehicle + DMSO (n = 6); NTG vehicle + URB602 (n = 6); NTG + URB602 (n = 6).
Rats in this group were anaesthetized with a lethal dose of anesthetic (chloral hydrate) and perfused transcardially with saline and 270–300 ml of ice-cold 4% paraformaldehyde 4 h after NTG or vehicle administration. Brains were removed, post-fixed for 12 h in the same fixative, and subsequently transferred in solutions of sucrose at increasing concentrations (up to 30%) during the following 72 h. All brains were cut at 50 µm on a freezing sliding microtome. c-Fos expression was measured using the free floating immunohistochemical technique with a rabbit polyclonal antiserum directed against c-Fos protein (residues 4–17 of human c-Fos). Coronal sections were incubated for 48 h at 4℃ with the c-Fos antibody (1:1000; Oncogene, Cambridge, MA, USA). After thorough rinsing in potassium phosphate buffered saline containing Triton X-100, sections were processed with the avidin–biotin technique, using a commercial kit (Vector Labs, UK). c-Fos staining was visualized with nickel-intensified 3′,3′-diaminobenzidine tetrahydrochloride (DAB).
Statistical evaluation
An a priori power analysis was conducted to determine the minimal sample size needed to obtain a statistical power of 0.80 at an alpha level of 0.05. In our previous study (23) we evaluated the difference of at least 20% in nociceptive response in the second phase of the orofacial formalin test (time of face rubbing) between rats injected with NTG and rats injected with vehicle (NTG vehicle) and we calculated a standardized effect size of 1.683 for this variable. The power analysis by GPower 3.1 estimated a sample size of at least six rats for an experimental group. The effects of treatments on the latency of the tail flick test were evaluated by means of the Wilcoxon rank-sum test (baseline vs. post-treatment). For the plantar formalin test, the total number of flinches/shakes evoked by formalin injection were counted separately for Phase I and for Phase II, while for the orofacial formalin test, the time spent (in seconds) in face rubbing was counted separately for Phase I and for Phase II. All data were tested for normality using the Kolmogorov-Smirnov normality tests and considered normal. Therefore, the differences between groups were analyzed by one-way analysis of variance (ANOVA) followed by the Newman-Keuls Multiple Comparison Test.
For c-Fos expression, cell counts of individual nuclei were made from every sixth section throughout their rostrocaudal extent for each rat. To avoid differences related to the asymmetrical sectioning of the brain, c-Fos-positive cells were counted bilaterally (three sections for each nucleus and the mean value obtained from the two sides was used for the statistical analysis). Image analysis was performed by an investigator unaware of the experimental design, using an AxioSkop 2 microscope connected to a computerized image analysis system (AxioCam MR5) equipped with dedicated software (AxioVision Rel 4.2) (Zeiss, Oberkochen, Germany). Differences between groups were analyzed by Kruskal-Wallis test followed by Dunn’s Multiple Comparison Test. A probability level of less than 5% was regarded as significant.
Results
Tail Flick test
NTG (NTG + DMSO) induced a hyperalgesic response in the tail flick test, as suggested by the significant decrease in the latency of the tail flick response four hours after NTG administration. DMSO (NTG vehicle + DMSO) did not show any significant influence on tail flick latency. URB602 in association with NTG vehicle did not exert any significant analgesic effect when compared to baseline values. By contrast, URB602 administration counteracted NTG-induced hyperalgesia, increasing baseline values of latency. Data are shown in Figure 1.
Activity of URB602 at the tail flick test in naïve rats and in rats treated with nitroglycerin (NTG). Dimethylsulfoxide (DMSO) in association with NTG vehicle (NTG vehicle + DMSO) did not show significant effect on latency when compared with baseline levels. URB602 in association with NTG vehicle (NTG vehicle + URB602) did not show significant effect on latency when compared with baseline levels. Rats treated with NTG and DMSO (NTG + DMSO) showed hyperalgesia, demonstrated by the significant reduction in the latency of tail flicking when compared with baseline levels. URB602 in association with NTG (NTG + URB602) significantly increases latency when compared with baseline levels. The latency change was evaluated four hours after NTG or NTG vehicle administration. Data are expressed as percent change (0%) from baseline ± SEM. Wilcoxon rank-sum test (post-treatment vs. baseline) *p < 0.05 vs. baseline.
Plantar formalin test
NTG (NTG + DMSO) administration significantly increased the total number of flinches/shakes in Phase II of formalin test when compared to NTG vehicle+DMSO group, thus confirming our previous findings (23,26). URB602 (NTG vehicle+URB602) inhibited nociceptive behavior only in Phase I of formalin test when compared to NTG vehicle+DMSO group. By contrast, when associated with NTG, URB602 significantly reduced the nociceptive behavior induced by NTG administration during Phase II of the test. Data are shown in Figure 2.
Activity of URB602 at the formalin test in naïve rats and in rats treated with nitroglycerin (NTG). NTG administration associated with dimethylsulfoxide (DMSO), significantly increased the total number of flinches/shakes in Phase II of formalin test, when compared to the NTG vehicle + DMSO group. URB602 in association with NTG vehicle significantly inhibited nocifensive behavior induced by formalin injection during Phase I when compared with the NTG vehicle + DMSO group. In rats treated with NTG, URB602 (NTG + URB602) significantly reduced the total number of flinches/shakes in Phase II of the test, when compared to the NTG + DMSO group. Data are expressed as mean ± SEM. ANOVA analysis followed Newman-Keuls Multiple Comparison test, *p < 0.05 vs. NTG vehicle + DMSO; °p < 0.05 vs. NTG + DMSO.
Orofacial formalin test
No differences were observed between all experimental groups regarding Phase I of the test. Regarding Phase II, NTG (NTG + DMSO) administration significantly increased nocifensive face-rubbing behavior when compared to the NTG vehicle + DMSO group. URB602 (NTG vehicle + URB602) significantly increased face rubbing time compared to the NTG vehicle + DMSO group. By contrast, when administered in association with NTG, URB602 significantly decreased NTG-induced behavior in Phase II. These results were also confirmed by administration of another MGL inhibitor, JZL184. Data are shown in Figure 3.
Effect of MGL inhibition during the orofacial formalin test. No differences were observed between all experimental groups as regards Phase I of the test. During Phase II, URB602 in association with nitroglycerin (NTG) vehicle (NTG vehicle + URB602) significantly increased the face rubbing time when compared to the NTG vehicle + DMSO group. By contrast, when administered in association with NTG, URB602 (NTG + URB602) significantly decreased NTG-induced nocifensive behavior. These results were also confirmed by administration of JZL-184. Data are expressed as mean ± SEM. ANOVA analysis followed by Newman-Keuls Multiple Comparison test **p < 0.01 vs. NTG vehicle + DMSO; ###p < 0.001 vs. NTG vehicle + DMSO; °p < 0.05 vs. NTG + DMSO; ***p < 0.001 vs. NTG + DMSO. DMSO: dimethylsulfoxide.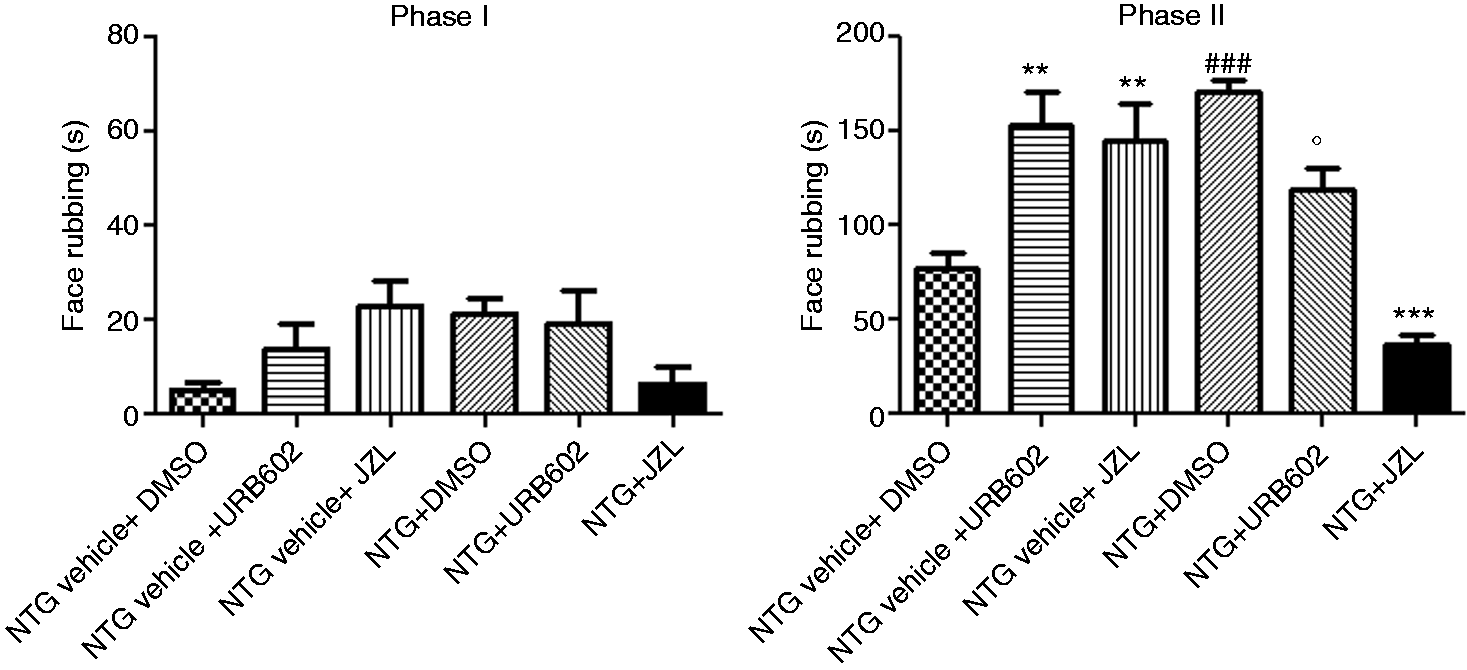
c-Fos expression
NTG (NTG + DMSO) administration induced a significant increase in c-Fos expression in the paraventricular nucleus of the hypothalamus (PVH), central nucleus of the amygdala (AMI), ventrolateral column of periaqueductal gray (PAG), LC, parabrachial nucleus (PAB), NTC and nucleus tractus solitarii (NTS), when compared to NTG vehicle + DMSO group. URB602 (URB602 + DMSO) did not change c-Fos expression in the brain nuclei under evaluation. When administered three hours after NTG, URB602 reduced NTG-induced c-Fos expression in all the cerebral areas that were examined; the reduction reached a statistical significant level in NTC and PAG. Data are shown in Figure 4.
a) Expression of c-Fos protein in specific brain areas after URB602 administration alone or in association with nitroglycerin (NTG). NTG (NTG + DMSO) administration induced a significant increase in c-Fos expression in PVH, AMI, PAG, LC, PAB, NTC and NTS compared to the NTG vehicle + DMSO group. URB602 reduced c-Fos expression in all the areas under evaluation only when administered in association with NTG (NTG + URB602); the reduction reached a statistically significant level in NTC and PAG. Data are expressed as mean ± SEM, Kruskal-Wallis test followed by Dunn’s Multiple Comparison test: *p < 0.05 vs. DMSO; °p < 0.05 vs. NTG + DMSO; b) Micrographs of representative sections of PAG and NTC of rats treated with NTG vehicle + DMSO, or NTG administered with DMSO (NTG + DMSO) or URB602, three hours after NTG (NTG + URB602).
Discussion
The present findings, together with previous results from our group (23,25), suggest that potentiation of endocannabinoid signalling has an anti-hyperalgesic effect in an animal model of migraine. The exact mechanisms involved in this effect are not fully understood, and multiple targets at different sites are likely to be involved. Endocannabinoids exert a critical control on cerebrovascular tone, by interacting with serotonergic transmission, nitric oxide (NO) production, and calcitonin gene-related peptide (CGRP) release (7). Human data support the involvement of a dysfunction in the endocannabinoid system in the pathology of chronic migraine: 2-AG and AEA levels were found to be reduced in platelets and cerebrospinal fluid of subjects with migraine (28,29) compared to control subjects. Pre-clinical studies show that the anti-nociceptive action of the endocannabinoids is related to a modulation of the trigeminovascular system, probably by the CB1 receptors localized on fibers in the spinal trigeminal tract and spinal NTC or CB1/CB2 receptors on non-neuronal cells (e.g. macrophages) (23,25,30). AEA was able to inhibit dural blood vessel dilation due either to electrical stimulation or to CGRP, capsaicin, or NO application. This effect was reversed by AM251, a CB1 receptor inverse agonist (17). The endocannabinoid 2-AG is involved in various (patho)physiological processes, it exerts numerous beneficial actions and it is the most abundant endocannabinoid in the central nervous system (CNS). The activity of this bioactive lipid depends on its endogenous levels, which are controlled by 2-AG-producing and 2-AG-hydrolyzing enzymes, and particularly by MGL, a presynaptic serine hydrolase that accounts for the majority of 2-AG degradation in brain tissue (31). The potential role of 2-AG signalling in spinal pain control has been demonstrated (9), and its enzymatic machinery is expressed in the superficial dorsal horn (32), with axonal and astrocytic localization (33). Pharmacological research has shown that selective MGL inhibitors produce analgesia in several animal models of pain and reduce inflammation in models of acute inflammatory pain (34,35). Very little is known, however, about the effects of MGL inhibitors at the trigeminal level.
In the present study, we investigated the role of 2-AG in modulating the spinal hyperalgesia induced by NTG in rats, a well-known animal model of migraine (23), by using the reversible MGL inhibitor URB602 (2). In the same model, we also evaluated the effect of URB602 on NTG-induced neuronal activation. In order to gain new insights on the role of MGL inhibition in trigeminal hyperalgesia, we evaluated the pharmacologic effects of URB602 and JZL184, an irreversible MGL inhibitor, in the NTG-induced hyperalgesia at the orofacial formalin test.
Our findings demonstrate that URB602 does not exert any analgesic effect per se in the tail flick test, but inhibits NTG-induced hyperalgesia. URB602 showed an analgesic effect in Phase I of the plantar formalin test. Conversely, its administration increased the time of face rubbing elicited by orofacial formalin injection in the second phase of the orofacial formalin test. This finding was surprising, since it suggests that a higher concentration of 2-AG may have an algesic effect at the trigeminal level. Though unexpected, the observed algesic effect of MGL inhibition in the trigeminal territory is not new. Spradley et al. (36) indeed reported differences in peripheral endocannabinoid modulation after MGL inhibition. In particular, the authors showed that an increase in peripheral levels of 2-AG was associated with a pro-nociceptive effect in the facial skin, while it had an anti-hyperalgesic effect in the hindpaw skin. It is possible that this topographically segregated effect is related to the formation of a metabolite of 2-AG occurring endogenously at the trigeminal level, thus specifically influencing the local nociceptive response (37). Taken together, these findings suggest that 2-AG levels may exert a differential effect on nociception, depending on the nature of the stimulus and on the tissue under investigation.
When analyzing NTG-induced changes in brain activity, URB602 treatment significantly reduced neuronal activation in the PAG and in the NTC, probably by inhibition of NTG-induced nuclear factor-kappa B expression (NF-kB) (38). 2-AG is an important endogenous signalling mediator that is known to protect neurons from pro-inflammatory, excitotoxic stimuli and other harmful insults (39). Several recent studies have shown that 2-AG protects CNS neurons from lipopolysaccharide-induced toxicity by suppressing the elevation of cyclooxygenase-2 (40,41). In addition, 2-AG inhibits cytokine release from both lipopolysaccharide-treated rat microglial cells and murine macrophages (42). An increased availability of 2-AG, might therefore prevent the activation of inflammatory pathways that is known to be initiated by NTG (14,38,41). In vivo, 2-AG protects against neuroinflammation in response to sulfur dioxide inhalation by attenuating the overexpression of iNOS. Furthermore, MGL inhibition causes suppression of NF-κB-p65 phosphorylation and COX-2 expression in response to pro-inflammatory insults by PPARγ receptors in mouse hippocampal neurons in vitro (40).
Regarding the tests evaluating nocifensive behavior, URB602 counteracted NTG-induced hyperalgesia in the tail flick test, as well as at the plantar and orofacial formalin tests. The anti-hyperalgesic effect of MGL inhibition was also confirmed when using JZL184 treatment. JZL184 is a more potent and selective MGL inhibitor than URB602, which displays excellent activity in vivo (43).
This finding further supports the hypothesis that one of the mechanisms underlying the anti-hyperalgesic effect of MGL in our animal model of migraine is the inhibition of the NF-kB pathway and/or cyclooxygenase-2 cascade as a consequence of the local increase in 2-AG levels (41,44). In a previous study, Nozaki et al. (30) reported that NTG-induced mechanical allodynia and neuronal activation of NTC were completely abolished in FAAH-deficient mice, but not in MGL-deficient mice, two hours after NTG or vehicle administration. Disparities between our findings and those of Nozaki et al. (30) may be due to differences in the pain model (allodynia versus formalin test or tail flick test), in species (rats versus mice), or in other methodological differences (e.g. the timing of sampling). As regards the latter issue, it must be noted that MGL knockout mice have a variable increase in 2-AG concentrations in the brain that ranges from 10 to 58-fold (45,46). We do not know the amplitude nor the exact temporal pattern of 2-AG increase in the brain in our model, which does not warrant any direct comparison of our study with the findings by Nozaki. Furthermore, genetic inactivation of MGL may impair the signalling of CB1 receptors, as suggested by Imperatore et al. (47). These considerations cannot rule out the possibility that URB602 exerts an indirect effect on 2-AG levels, which does not depend on MGL inhibition. Additional research is needed to elucidate the intriguing signalling mechanisms related to the role of 2-AG in migraine pain and in trigeminal nociception.
The anti-hyperalgesic effect of URB602 observed in this study is much stronger than its analgesic effect. This probably reflects a complex condition in which the increase of brain NO levels associated to NTG administration (48), on one side, results in a reduced synaptic availability of 2-AG as a consequence of its increased uptake from the cells (49), while, on the other, it activates inflammatory pathways (14, 20,38).
The action of URB602, which is neutral or attenuated under physiological conditions, might become amplified, and therefore detectable with our probes, in the “sensitized condition” created by NTG administration.
Similar to the present data, in a previous study we have shown that peripheral inhibition of FAAH activity by URB937 counteracted NTG-induced hyperalgesia (23). At variance with this, URB937 per se did not induce any algesic effect in the orofacial formalin test. If we compare the effects observed upon peripheral FAAH inhibition with URB937 with those obtained with the non-selective inhibition of MGL caused by URB602 in the same experimental paradigm, a ‘dissociated’ effect of the two endocannabinoids in the trigeminal district becomes apparent. A possible explanation for this discrepancy may reside in a tissue-specific metabolism. Formalin injection might indeed cause a strong increase in tissue levels of 2-AG; the maintenance of high 2-AG concentrations in either central or peripheral tissues, as a result of MGL inhibition, could contribute to the formation of pro-nociceptive metabolites (37). 2-AG may indeed represent an important source of arachidonic acid and neuroinflammatory prostanoids, such as PGE2-G metabolite, in some circumstances, thus driving pro-nociceptive mechanisms. By contrast, URB937, which does not cross the blood-brain barrier, mainly acts by maintaining higher AEA levels released by neurons localized in the injured peripheral tissues (the upper lip, in our experimental paradigm). Another possible explanation of the differential effects may be searched in the receptors that are involved in the biologic activity of AEA and 2-AG. AEA activates vanilloid type 1 receptors (TRPV1) and CB1 receptors, whereas 2-AG is a full agonist of CB1 and CB2 receptors, devoid of effect on TRPV1 (50). Thus, in our paradigm, increased availability of AEA might activate CB1 receptors or desensitize TRPV1 expressed in primary sensory neurons, thus reducing the transmission of pain to central areas (51), probably via the inhibition of NO release and/or of neuropeptides from primary afferents at the spinal (52) and possibly at the trigeminal level. However, it is known that a wide range of central FAAH inhibitors injected at variable doses may induce the opposing effect in the same model of pain; furthermore, some of them did not show a clear dose dependence in nociceptive models. It is worth noting that the activation of CB1 and TRPV1 receptors may indeed have opposing effects on nociception, thus confounding the interpretation of results (53). Altogether, our findings strongly suggest that FAAH and MGL inhibition modulates migraine pain, though other studies are needed to address more precisely the mechanism underlying their biologic effect and to characterize more precisely their profile of action at the trigeminal level.
Conclusions
MGL inhibition, and the likely increase in central or peripheral levels of 2-AG, reduces pain sensitivity during NTG-induced hyperalgesia at the nociceptive tests and it attenuates neuronal activation in specific brain areas involved in cephalic pain. The present findings show that inhibition of MGL represents a promising candidate for the development of drugs for migraine treatment. Additional research is needed to elucidate the intriguing signalling mechanisms related to the role of 2-AG at the trigeminal level; in particular, the evaluation of 2-AG levels in the tissues of interest will provide evidence to support or refute our hypothesis.
Article highlights
URB602, a reversible monoacylglycerol lipase (MGL) inhibitor, significantly reduces nitroglycerin (NTG)-induced c-Fos expression in the nucleus trigeminalis caudalis and the locus coeruleus in an animal model of migraine. URB602 is also effective in counteracting NTG-induced hyperalgesia during the tail flick test and both the plantar and orofacial formalin tests. URB602 potentiated formalin-induced hyperalgesia in the trigeminal area when used alone. The latter result was also confirmed after treatment with JZL184, an irreversible MGL inhibitor.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant of the Italian Ministry of Health to Institute C. Mondino (RC 2014-2016).