
Abstract
Background
Cluster headache is characterized by recurrent unilateral headache attacks of severe intensity. One of the main features in a majority of patients is a striking rhythmicity of attacks. The CLOCK (Circadian Locomotor Output Cycles Kaput) gene encodes a transcription factor that serves as a basic driving force for circadian rhythm in humans and is therefore particularly interesting as a candidate gene for cluster headache.
Methods
We performed an association study on a large Swedish cluster headache case-control sample (449 patients and 677 controls) screening for three single nucleotide polymorphisms (SNPs) in the CLOCK gene implicated in diurnal preference (rs1801260) or sleep duration (rs11932595 and rs12649507), respectively. We further wanted to investigate the effect of identified associated SNPs on CLOCK gene expression.
Results
We found a significant association with rs12649507 and cluster headache (p = 0.0069) and this data was strengthened when stratifying for reported diurnal rhythmicity of attacks (p = 0.0009). We investigated the effect of rs12649507 on CLOCK gene expression in human primary fibroblast cultures and identified a significant increase in CLOCK mRNA expression (p = 0.0232).
Conclusions
Our results strengthen the hypothesis of the involvement of circadian rhythm in cluster headache.
Introduction
The pathophysiology of cluster headache (CH) is largely unknown, but one of the main features is a striking rhythmicity of attacks. The CLOCK (Circadian Locomotor Output Cycles Kaput) gene is located on chromosome 4q12 and encodes a transcription factor that serves as a basic driving force for circadian rhythm in humans together with BMAL1 (Brain and muscle arnt-like protein 1) (1). Its key role in the circadian clock makes CLOCK a particularly interesting candidate gene for CH pathophysiology, considering the often-featured periodical occurrence of CH attacks. A number of studies show a genetic contribution to CH and 7–20% of the patients report a positive family history, defined as having at least one first, second, or third-degree relative diagnosed with CH (2,3). Previous studies have investigated candidate genes with a possible link to CH, such as the genes HCRTR2 (4–8), ADH4 (7,9,10), CACNA1A (11), NOS (12), and PER3 (13). For HCRTR2 and ADH4, conflicting results have been reported. Only three smaller case-control studies (≤107 CH patients and ≤210 controls) have investigated a possible causative role of one genetic variant (rs1801260) in CLOCK in relation to CH. All of these studies were done in Italian cohorts and reported a lack of association between the single nucleotide polymorphism (SNP) rs1801260 and CH (7,14,15). Rs1801260 has previously been linked to modifying diurnal preference (16). We wanted to investigate the frequency of rs1801260 in the largest Swedish CH case-control material (449 patients and 677 controls) collected to date. We further aimed to screen two additional CLOCK SNPs, rs11932595 and rs12649507, previously reported to be implicated in sleep duration (17). The SNP rs1801260 is located in the 3′ UTR, rs11932595 in intron 10–12, depending on splice variant, and rs12649507 in intron 1 of the CLOCK gene.
Moreover, we were interested in investigating whether general CLOCK gene expression differs between patients and controls, and if disease-associated CLOCK SNPs lead to changes in CLOCK gene expression.
Materials and methods
Demographic characterization.

Based on 398 individuals for whom detailed information was available. CH = cluster headache; n/a = not available.
Genotyping
DNA was extracted from whole blood using standard protocols. Genotyping was performed using quantitative Real-Time PCR (qPCR). We used pre-designed TaqMan® SNP genotyping assays (rs1801260: C___8746719_20; rs11932595: C____296556_10; rs12649507: C___1836992_10) and TaqMan® Genotyping Master Mix (Applied Biosystems, Carlsbad, CA, USA). The ABI 7500 Fast Real-Time PCR instrument (Applied Biosystems, Foster City, CA, USA) was programmed according to the manufacturer’s instructions with an increased cycle number.
Cell culture and gene expression
For gene expression experiments, cutaneous biopsies were obtained at Karolinska University Hospital. Biopsies were taken in a subsample of the study cohort; 11 CH patients and 11 healthy control individuals. From the biopsies, fibroblast cell cultures were established according to the following protocol: Biopsies were minced and placed in 35 mm dishes (Corning Inc., Corning, NY, USA) under a sterile glass coverslip and cultured in DMEM GlutaMAX™ (supplemented with 25 mM D-Glucose and 1 mM sodium pyruvate), 10 mM HEPES, 1 x Antibiotic-Antimycotic (supplemented with 10,000 U/mL penicillin, 10,000 µg/mL streptomycin, and 25 µg/mL Amphotericin B), and 15% fetal calf serum (all from Invitrogen, Paisley, UK), in a humidified 37℃, 5% CO2 incubator . Fibroblasts were harvested when growing confluent around the explant and passaged two times (20). Cells were reseeded into 75 cm2 culture flasks (Corning Inc., Corning, NY, USA); at confluence, two to three days later, treatments were performed. Cell cultures from patients and controls were furthermore given a high-serum treatment defined as zeitgeber (ZT) + 0 h in order to synchronize their circadian clocks according to the protocol of Johansson et al. (21). CLOCK has previously been reported to be rhythmically expressed in human fibroblasts with a peak in the evening (22–24). Fibroblasts were therefore harvested 12 hours after serum shock treatment (ZT + 12 h) and frozen at −80℃. RNA and cDNA were prepared from frozen cells using the QIAGEN RNeasy Mini Kit and the QuantiTect Reverse Transcription Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s protocols.
The qPCR was performed by using CLOCK primers, designed with the web-based software Primer3 (25) (F: 5′-CCTGAGACAGCTGCTGACAA–3′; R: 5′-TGGTTGGTGTTGAGGAAGGG–3′) (Thermo Fisher Scientific, Waltham, MA, USA), and Power SYBR® Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA). A pool of all 11 control samples comprised the reference sample to which the study samples were compared. As endogenous control for normalisation of CLOCK mRNA expression, the housekeeping gene GAPDH (Glyceraldehyde 3-phosphate dehydrogenase) with self-designed primers (F: 5′-CACATGGCCTCCAAGGAGTAA–3′; R: 5′-TGAGGGTCTCTCTCTTCCTCTTGT–3′) was chosen. The ABI 7500 Fast Real-Time PCR instrument was programmed for quantitation by a relative standard curve according to the manufacturer’s recommendations, with increased cycle number.
Statistical analyses
Genotype association was evaluated with a chi-square (χ2) test, and allele association was analyzed with Fisher’s exact test, or a χ2 test for a very large sample size (n > 2000), using GraphPad Prism v5.04 (GraphPad Softwares Inc., La Jolla, CA, USA). Additionally, genotypic association was tested for via logistic regression with sex as a covariate using the PLINK whole genome association analysis toolset v1.07 (26). For stratified analysis, the patient group was subdivided by their reporting of whether CH attacks occurred with diurnal rhythmicity (n = 262) or not (n = 191). Haplotype analysis was performed in HaploView v.4.2 (27). For power analysis, the power and sample size software PS v3.022 (28) was used. With our sample size and an estimated minor allele frequency of 0.26 in individuals with European descent (http://www.ncbi.nlm.nih.gov/SNP), we have 80% power to detect true odds ratios (OR) for CH below 0.662 or above 1.451, respectively. The observed frequencies of controls were in agreement with the Hardy-Weinberg equilibrium, which was tested for using the web-based Online Encyclopedia for Genetic Epidemiology studies software (29). Gene expression data were analysed using qBase v1.3.5 (30). Relative gene expression is reported as log2-transformed values in order to pass normality tests. To compare mean expression between groups, one-way ANOVA was used in Graph Pad Prism v5.04. The difference in frequency of diurnal rhythmicity between men and women was tested for using Student’s t-test. All statistical tests were performed with a significance level of 0.05. Bonferroni correction for multiple testing was applied when relevant for the number of studied SNPs in all single marker analyses. A permutation test with 10,000 permutations was performed for the haplotype analysis.
Results
Genotype and allelic analysis
Genotype and Allele frequencies for three CLOCK SNPs in CH patients and controls.
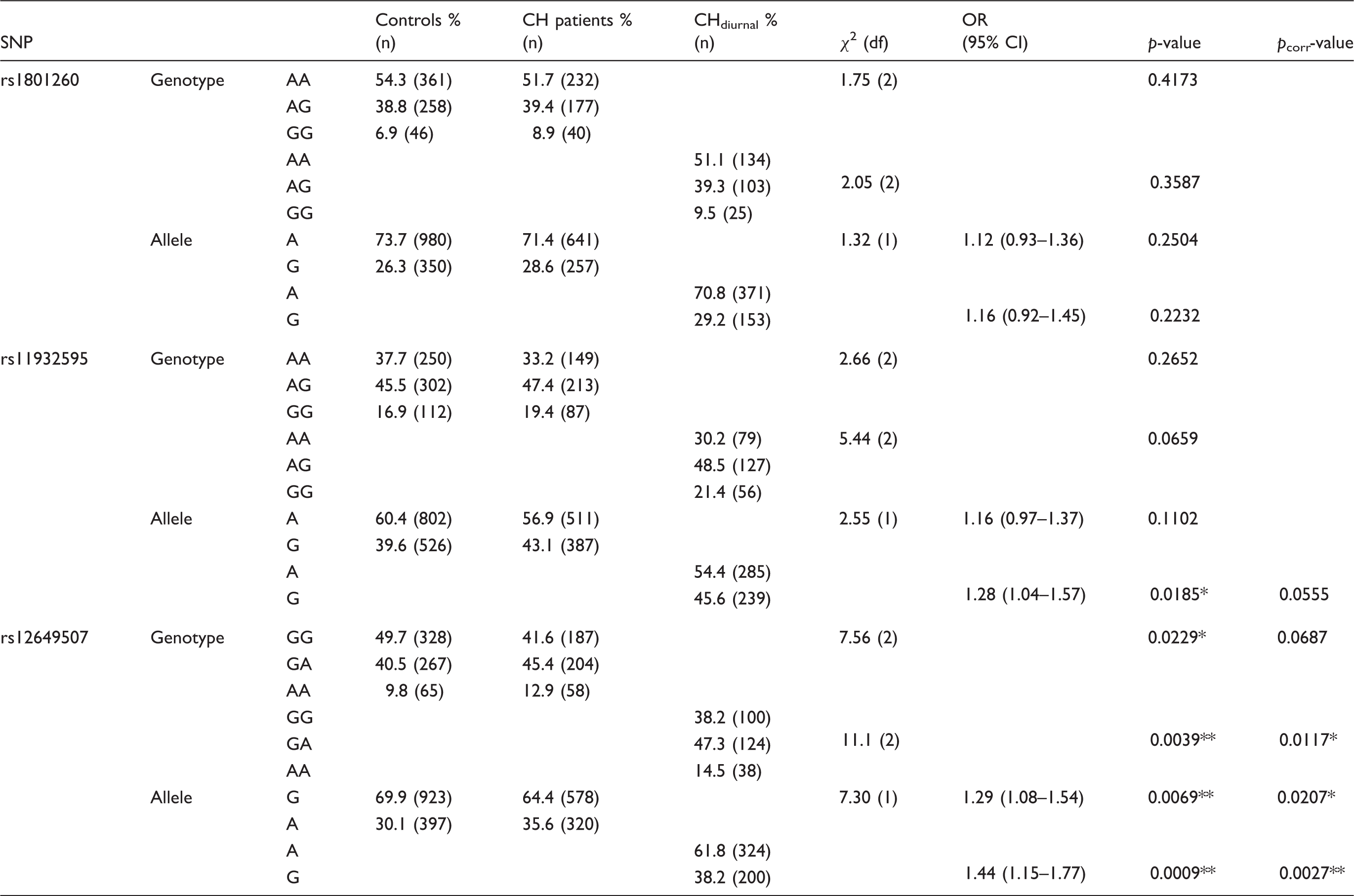
CH = cluster headache; CHdiurnal = cluster headache patients reporting attacks occurring with diurnal rhythmicity; χ2 = chi-square; CI = confidence interval; df = degrees of freedom; OR = odds ratio; pcorr-value = p-value corrected for multiple testing; SNP = single nucleotide polymorphism. *p < 0.05; **p < 0.01.
Logistic regression with sex as a covariate was run in order to exclude any bias introduced by the skewed sex ratio in the patient group. This analysis confirmed the results obtained with a χ2 test: For rs12649507, there was an association with CH (p = 0.0295) that became even stronger when only comparing the patient group with diurnal rhythmicity to controls (p = 0.0072). We could not find a genotypic association with CH for either rs1801260 (p = 0.2636) or rs11932595 (p = 0.1887) when applying sex-corrected logistic regression.
With regard to possible gender differences, we also compared how many men and women with CH reported diurnal rhythmicity. In total, 63.3% of the men and 77.2% of the women who filled out the questionnaire had diurnally recurring attacks: this difference is significant (p = 0.0061).
Haplotype analysis
Haplotype analysis for three CLOCK SNPs in CH patients and controls.

Reference haplotype (A-A-G) corresponds to wild-type allele of each CLOCK single nucleotide polymorphism (SNP) where rs1801260 is in position 1, rs11932595 is in position 2, and rs12649507 is in position 3.
CH = cluster headache; χ2 = chi-square; df = degrees of freedom; pcorr-value = p-value after test with 10,000 permutations. *p < 0.05; **p < 0.01; ***p < 0.001.
Gene expression analysis
Relative CLOCK gene expression levels were compared between CH patients (mean log2 expression ± standard deviation = −0.435 ± 0.742) and controls (−0.292 ± 0.979), see Figure 1a and supplementary table 1 for specific CLOCK gene expression information for each individual. No significant difference in CLOCK gene expression was detected between patients and controls (p = 0.7095).
CLOCK mRNA expression in correlation to disease status/genotype. (a) Quantification of CLOCK mRNA levels in human fibroblasts from controls (n = 11) and cluster headache (CH) patients (n = 11), normalized to the housekeeping gene GAPDH; (b) CLOCK mRNA expression levels in the same individuals grouped by the three different rs12649507 genotypes GG (n = 7), GA (n = 13), and AA (n = 2), compared using one-way ANOVA.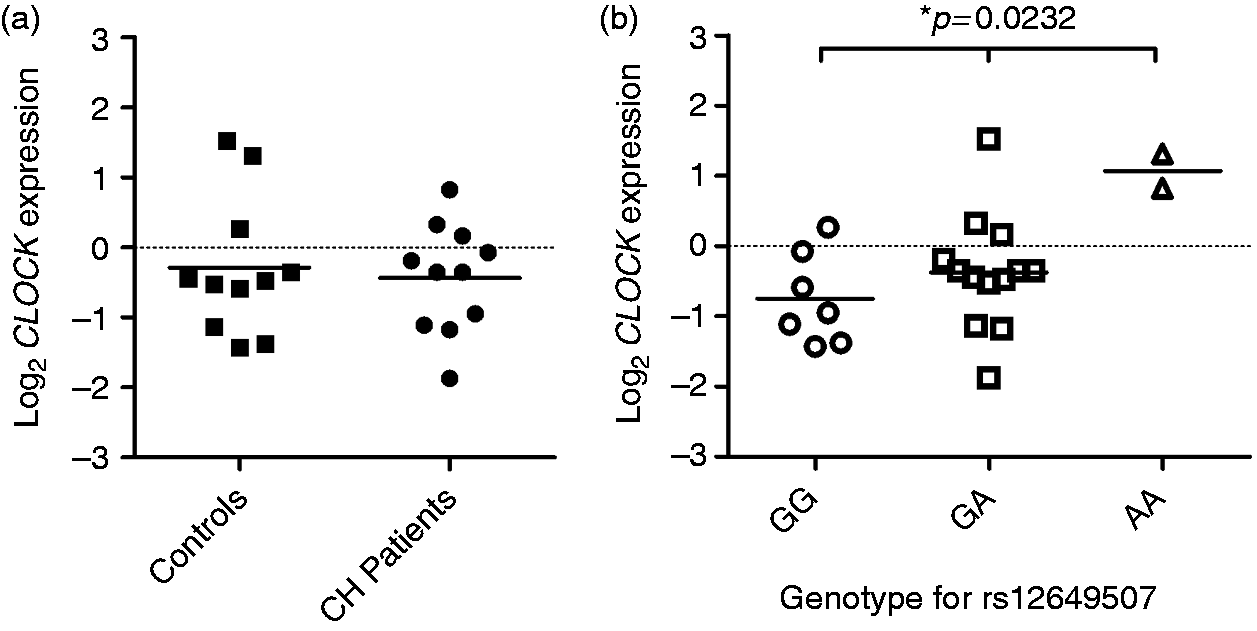
To investigate whether CLOCK mRNA expression can be linked to rs12649507, the 22 samples, independent from disease phenotype, were grouped by the rs12649507 genotype that each individual exhibited (Figure 1b). By doing this, we found that mean CLOCK gene expression is significantly different between the three groups (p = 0.0232). Mean CLOCK expression is lowest in individuals with the wild-type genotype GG (−0.752 ± 0.651) and increases with each addition of the less prevalent A-allele (GA: −0.374 ± 0.782; AA: 1.068 ± 0.343).
Discussion
We found a significant association between the CLOCK gene variant rs12649507 and CH. The less common A-allele of this SNP was consistently more prominent in CH patients than controls throughout genotype, allele and haplotype analysis. Interestingly, rs12649507 has previously been implicated in sleep duration (17). In two different populations, it was shown that individuals with the rs12649507 AA genotype exhibited significantly shorter sleep duration compared to individuals with the GG or GA genotype. Sleep patterns have not yet been investigated in our CH cohort, but are subject to future studies. However these studies need to carefully consider the reduced sleep quality reported in patients in bout compared with controls; in addition to that, sleep quality improves between clusters (31).
No association could be found with rs1801260 and CH in this large Swedish case-control study, which is in agreement with the smaller Italian studies (7,14,15). One limitation of our association study could be the control subjects, where a majority were anonymous blood donors with no information about age etc. However, we still consider this an excellent control group representing a general, healthy Swedish adult population between 18–60 years of age (https://geblod.nu/english-engelska/).
To characterize whether CLOCK gene activity is altered in CH, CLOCK mRNA expression levels were compared between CH patients and controls. We did not find a difference between the two groups regarding CLOCK expression. However, when grouping all samples by rs12649507 genotype, there was a clear link between differential CLOCK expression and the CH-associated SNP rs12649507. This suggests that the AA genotype specifically leads to increased CLOCK gene expression. This is in accordance with the eQTL (expression quantitative trait locus) findings reported on the GTEx portal website (www.gtexportal.org), where rs12649507 could be linked to a change in CLOCK expression in several tissues with the addition of the minor A-allele, for example sun-exposed skin cells (p = 7.3 × 10−7). From these results, we hypothesize an increased expression of CLOCK in CH patients, as the AA genotype is more prevalent in this group. However, our biopsy material is relatively small (CH = 11 vs. controls = 11) and there were only two individuals with the AA genotype, one in the patient group and one control individual.
CLOCK is an important modulator of circadian rhythms and is strongly expressed in the brain region responsible for controlling circadian rhythms, the suprachiasmatic nucleus, located in the hypothalamus (1,32). Furthermore, it has been shown that the hypothalamus is specifically activated during CH attacks (33). The underlying mechanism of how CLOCK could modulate CH via the hypothalamus is yet to be determined, but one hypothesis is that altered CLOCK activity, caused by, for example, a genetic variant, may affect the phase of the internal clock. This may lead to a mismatch between the internal clock and external time cues, causing a circadian misalignment that has been reported to lead to different health problems over time (13,34,35).
In conclusion, we identified the first association with a genetic variant in CLOCK, the A-allele of rs12649507, and cluster headache, and could further show a functional consequence of the rs12649507 on CLOCK gene expression. Our data support circadian rhythm involvement in the disease, which could lead to new treatment strategies. However, the functional consequence of increased CLOCK mRNA activity needs to be further investigated.
Article highlights
Significant association between rs12649507 in CLOCK and cluster headache was observed in Sweden (p = 0.0069). No difference in CLOCK mRNA expression between cluster headache patients and controls. CLOCK mRNA expression is dependent on the rs12649507 genotype.
Footnotes
Acknowledgements
We thank Ann-Christin Karlsson for help with the recruitment of patients and Fengqing Xiang for technical assistance.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Swedish Brain Foundation [FO2013-0213], Swedish Research Foundation [2012-02211], Åke Wiberg Foundation [ID:756194137K2012], Swedish Migraine Society, and Karolinska Institutet Research Funds [2014fobi41360].
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.