
Abstract
Objective
To review and discuss the putative role of light, sleep, and the biological clock in cluster headache.
Discussion
Cluster headache attacks are believed to be modulated in the hypothalamus; moreover, the severe pain and typical autonomic cranial features associated with cluster headache are caused by abnormal activity of the trigeminal-autonomic reflex. The temporal pattern of cluster headache attacks suggests involvement of the biological clock, and the seasonal pattern is influenced by the number of daylight hours. Although sleep is often reported as a trigger for cluster headache attacks, to date no clear correlation has been established between these attacks and sleep stage.
Conclusions
We hypothesize that light, sleep, and the biological clock can change the brain’s state, thereby lowering the threshold for activating the trigeminal-autonomic reflex, resulting in a cluster headache attack. Understanding the mechanisms that contribute to the daily and seasonal fluctuations in cluster headache attacks may provide new therapeutic targets.
Introduction
Cluster headache is characterized by agonizing headache attacks with strictly side-locked, excruciating temporal or intraorbital/periorbital pain lasting anywhere from 15 minutes to 3 hours; these attacks are often accompanied by ipsilateral autonomic cranial features such as lacrimation, ptosis, miosis, nasal congestion, and/or rhinorrhoea (1). Cluster headache is one of the so-called trigeminal autonomic cephalalgias, a class of headache disorders characterized by brief, side-locked headache attacks accompanied by cranial autonomic symptoms (1). The prevalence of cluster headache in the general population is 1 in 1000 and it is generally more common among men than among women, with a ratio of 2.5–3.5 to 1 (2). A fascinating aspect of cluster headache is the currently unexplained temporal pattern of the attacks. Attacks typically occur at specific times during the day, and a seasonal rhythm is also common, with attacks occurring more frequently and/or severely in specific months of the year (3–5). These temporal patterns have led to the question of whether the circadian system plays a causal role in the pathogenesis of cluster headache.
The circadian system is controlled by the suprachiasmatic nuclei (SCN), a bilateral structure located at the base of the anterior hypothalamus (6). Interestingly, cluster headache is believed to originate in and/or be modulated by the hypothalamus and has been treated using hypothalamic stimulation in patients with severe forms of cluster headache (7–11). In this review, we evaluate the putative role of the circadian system in the onset and frequency of cluster headache attacks.
In episodic cluster headache – the most common form of the disease – the attacks occur in episodes with an interval of weeks or months, with remission periods lasting several months to years (1). However, approximately 14% of cluster headache patients do not have a remission period and therefore suffer from the chronic form of the disease (12,13). Cluster headache attacks are associated with autonomic cranial features and a sense of restlessness and/or agitation (1), symptoms that can be explained by hyperactivation of the trigeminal-autonomic reflex (14), a reflex arch formed by the facial/greater superficial petrosal nerve dilator pathway as the efferent and the trigeminal nerve as the afferent pathway (15). This reflex is activated by stimulation of the trigeminal ganglion, which results in vasodilation of the internal carotid arteries through the release of vasodilator peptides such as calcitonin gene-related peptide (CGRP), substance P and neurokinin A (16).
During a cluster headache attack, the levels of plasma CGRP and vasoactive intestinal peptide (VIP) increase, and dilation of the ipsilateral internal carotid artery and the resulting increased blood flow have been observed using magnetic resonance angiography (MRA) and positron emission tomography (PET), supporting activation of the trigeminal-autonomic reflex (7,8,17–19). Furthermore, injecting capsaicin, which is known to trigger the trigeminal-autonomic reflex (20,21), into the forehead leads to symptoms similar to a cluster headache attack (8). It has also been hypothesized that this reflex underlies the escalation of pain during a cluster headache attack (22). Interestingly, however, other autonomic processes such as heart rate, blood pressure, and autonomic cardiovascular responses do not differ between patients with cluster headache and control subjects (23).
There is currently no doubt that cluster headache originates at the level of the hypothalamus, in contrast to many other forms of headache. However, there is no consensus on the area within the hypothalamus, but most studies point to the posterior hypothalamus. Specifically, positron emission tomography (PET) and functional magnetic resonance imaging (fMRI) have revealed that the ipsilateral inferior posterior hypothalamic gray area is activated during cluster headache attacks (8–10); importantly, this activity is not present in cluster headache patients during remission periods (7,8). Activity in other brain areas associated with the processing of pain, including the thalamus, cingulate cortex, and insular cortex, has been observed during a cluster headache attack, as well as during other types of headache (7,8). In addition, extensive studies of possible structural differences between cluster headache patients and control subjects have been performed, and one study found that the posterior hypothalamus is enlarged in patients (24). Another study found evidence of enlargement of the anterior hypothalamus in patients with a form of trigeminal autonomic cephalalgia (25).The structural abnormalities in the anterior hypothalamus of patients are particularly interesting, given that this is the location of the SCN (i.e. immediately above the optic chiasm), which generates circadian rhythms (25,26). Two additional studies found changes in a variety of brain structures connected to the hypothalamus and in brain areas related to emotion handling and nociception; however, neither of these studies found evidence of structural abnormalities in the hypothalamus (27,28). There are discrepancies between the hypothalamic imaging studies that prohibit a clear conclusion at this point. One possibility is that fMRI/PET do not have sufficient special resolution, resulting in slightly different outcomes, and/or the precise location within the hypothalamus that leads to cluster headache is different among the patient groups. Additionally, there is no exactly known relation between an enlarged hypothalamus and its function. However, the consensus is that the hypothalamus is an area of great interest in cluster headache, even if the exact location (i.e. anterior vs. posterior) remains unclear.
Further evidence supporting a role of the hypothalamus in cluster headache comes from a recent report that deep brain stimulation of the posterior hypothalamus improved symptoms in patients with drug-resistant chronic cluster headache (11). Moreover, lithium is often effective in treating cluster headache. It is important to note that lithium was one of the first substances found to affect clock function by lengthening the circadian period (29). The working mechanism of lithium for cluster headache remains unknown. Changes in the hypothalamus, combined with the temporal pattern of cluster headache attacks, suggest that the central clock of the SCN plays an important role in cluster headache.
Neurons in the SCN have the autonomous ability to produce circadian rhythms (26). This rhythmicity at the single-cell level arises via a negative-feedback loop between circadian clock genes and their protein products (30). These protein products activate currently unidentified pathways in order to change the cell’s membrane potential and electrical frequency (for review, see (31)). In both nocturnal species (e.g. rodents) and diurnal species (e.g. humans), electrical activity in the SCN is relatively high during the day and low during the night (31), creating a waveform of electrical activity that is directly correlated with the animal’s temporal pattern of behavioral activity (32).
The rhythm of the SCN is synchronized to the external light-dark cycle (33). External light is sensed in the retina by rod and cone photoreceptors and by intrinsically photosensitive retinal ganglion cells (ipRGCs); ipRGCs are directly photosensitive due to the presence of the photopigment melanopsin (33). The rod and cone photoreceptors transmit light information through the ipRGCs to the ventral SCN via a monosynaptic pathway known as the retinohypothalamic tract (RHT) (33). The release of glutamate and pituitary adenylate cyclase-activating peptide (PACAP) at RHT terminals triggers an increase in intracellular calcium in the SCN, which changes the rhythm of circadian clock genes (26).
This direct connection between light and the SCN is interesting, given that hypersensitivity to light – that is, photophobia – is a symptom that often accompanies primary headaches (1). Although typically associated with migraine, photophobia has also been reported to occur in 45–91% (34,35) of patients with cluster headache. The fact that external light can aggravate headache pain suggests a link between the nociceptive and optic pathways.
Interestingly, photophobia can still occur in migraine patients who are visually blind due to a loss of rod cone and cone cells, but whose retinas can still detect light (36). In contrast, patients with blindness due to damage to the optic nerves or the complete loss of both eyes do not experience photophobia (36). Additionally, these patients do not exhibit photoentrainment and, in the case of patients with optic nerve damage but intact eyes, also do not exhibit pupillary responses (36). The fact that only visual blind patients experience photophobia suggests that photophobia involves the non-image-forming pathway mediated by ipRGCs and melanopsin. This hypothesis is supported by the finding that light evokes melanopsin-dependent neuronal patterns associated with an experience of aversion in neonatal mice (37). The precise neuronal circuit that underlies photophobia is currently unknown; however, three pathways that can transmit information from the ipRGCs in the retina to the brain have been identified.
The first pathway was identified by tracing the axons of ipRGCs to trigeminal vascular dura-sensitive neurons in the posterior thalamus (36). These neurons are modulated by stimulation with light, thereby representing a retino-thalamo-cortical pathway by which light can modulate the neuronal activity of nociception in mice (36). Both fMRI and diffusion weighted imaging studies indicate that humans also have a retino-thalamo-cortical tract (38,39). A second pathway involves the caudal trigeminal complex and is activated by retinal stimulation (40). Finally, a third pathway is formed by ipRGCs and/or ipRGC-like melanopsin-containing neurons, which are present in the iris and retina and project to trigeminal afferents (41,42). Given the presence of these three – and possibly more – pathways, understanding the transmission of information from ipRGCs to the brain may help identify and characterize the mechanism underlying photophobia.
In migraine patients, the severity of photophobia increases with increasing light intensity; however, whether this is also the case in cluster headache patients is currently unknown (36). Color is also a determinant for photophobia; visual blind migraineurs have observed that blue light worsens their photophobia (43), and green light has significantly less effect on migraine compared to other colors such as white, amber, blue, and red (44); at low intensities, green light is even considered soothing (44). Interestingly, the dura-photosensitive trigeminovascular neurons in the posterior thalamus have the highest response to blue light and the lowest response to green light (44); this is particularly relevant given that melanopsin, the photopigment in ipRGCs, is most sensitive to blue/green light (45–47). The fact that photophobia is wavelength-dependent in migraineurs raises the possibility that the same anatomical pathway might underlie the photophobia reported in some patients with cluster headache.
Factors that influence the risk of cluster headache attacks
Cluster headache is characterized by temporal patterns of unexplained origin. As illustrated in Figure 1, the attacks can occur in clusters of weeks to months with a seasonal rhythm, or they can occur more frequently and can be more severe in certain months of the year (3–5). Within a cluster episode, the attacks typically occur at certain times of the day and/or night. This rhythmicity of attacks may be due to the effects of the biological clock and/or sleep.
Circadian and seasonal rhythms affect the frequency of cluster headache attacks. (a) The 24-hour day-night rhythm (with day shown in yellow and night shown in gray) leads to a synchronized 24-hour rhythm in the frequency of cluster headache attacks. (b) Although the seasonal rhythm (with summer shown in green and winter shown in brown) is less pronounced, many studies indicate that the pattern of cluster headache attacks is also synchronized to this rhythm.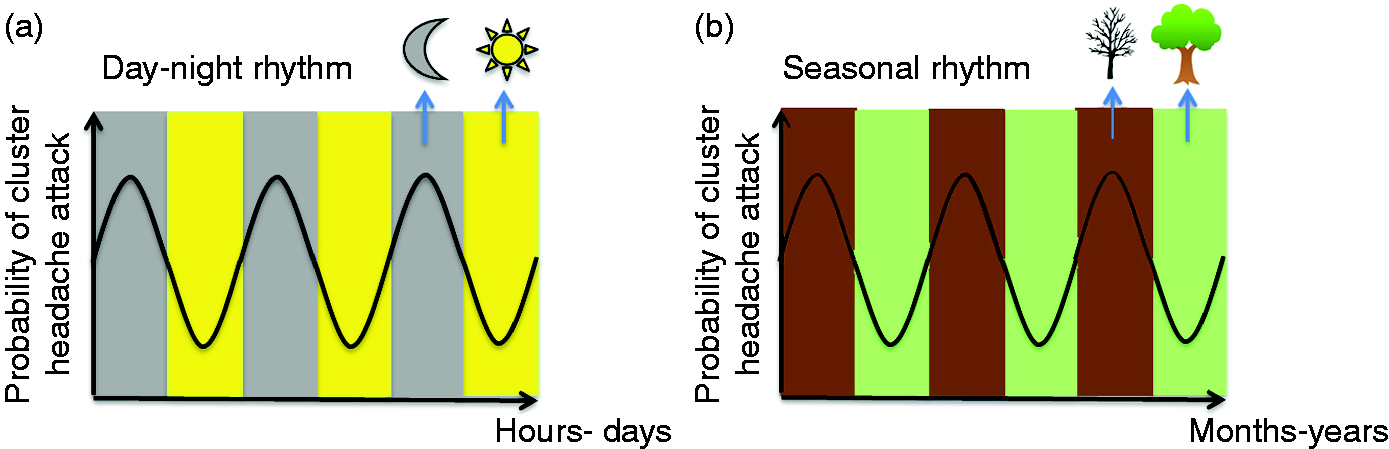
The day-night cycle
Cluster headache attacks typically occur at predictable times (3), and approximately 80% of cluster headache patients report that their attacks generally follow a 24-hour cyclical pattern (3,5,48). Overall, 75% of attacks occur between 9 pm and 10 am (49), with peak incidence at 2 am (3,5,49,50). The mechanisms that underlie this rhythmicity are currently unknown. Interestingly, this diurnal cycle of attacks has been reported to shift by approximately one hour in women compared to men; however, other features of diurnal rhythmicity are similar between sexes (48). A recent genetics study found a correlation between cluster headache and the rs12649507 single-nucleotide polymorphism (SNP) in the CLOCK (Circadian locomotor output cycles kaput) gene (51). In contrast, no correlation was found between cluster headache and a polymorphism in the PER3 gene or the T-> C polymorphism in the CLOCK gene (52,53).
Sleep and cluster headache
Although nocturnal sleep is frequently reported as the most common trigger of cluster headache attacks (23), one-third of patients also report that napping during the daytime can trigger attacks (5). Interestingly, most cluster headache attacks occur within 90 minutes of falling asleep (54). In addition, many patients anecdotally report that they fear going to bed, actively postpone sleep, and/or use voluntary sleep deprivation to avoid experiencing an attack. Moreover, cluster headache patients generally experience poor sleep quality, even during periods of remission (5). Importantly, however, both sleep quality and sleep patterns returned to normal in patients with chronic cluster headache following successful treatment with deep brain stimulation (55).
Overview of data regarding the occurrence of cluster headache attacks during specific sleep stages.
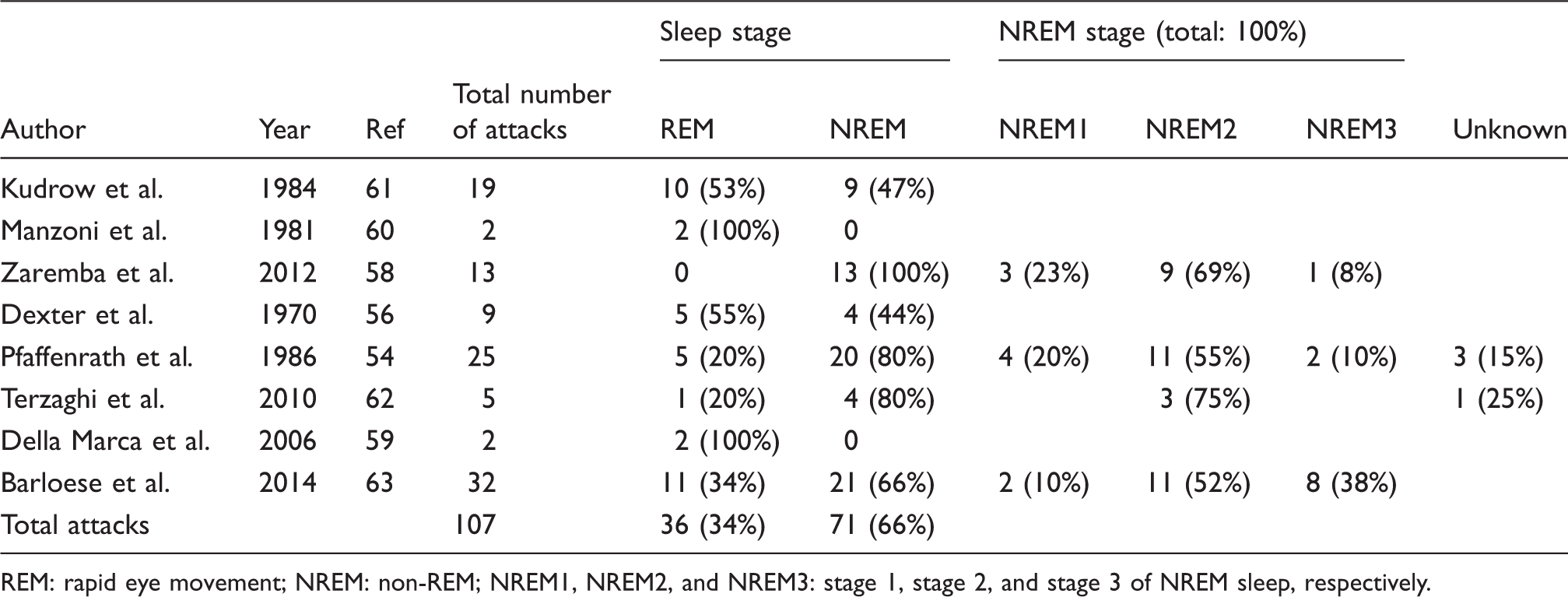
REM: rapid eye movement; NREM: non-REM; NREM1, NREM2, and NREM3: stage 1, stage 2, and stage 3 of NREM sleep, respectively.
Hormonal changes
Hormones are closely related to the biological clock, as the SCN imparts is rhythmic activity to the autonomic nervous system and the hypothalamic-pituitary-adrenal axis (65). Moreover, case reports have described pituitary tumors that were believed to cause symptomatic cluster headache attacks (66,67). Hormonal changes have been observed in cluster headache patients and include changes in growth hormone, prolactin, cortisol, follicle-stimulating hormone (FSH), luteinizing hormone (LH), testosterone, and melatonin levels (68). The release of the hormone melatonin is inhibited by light acting via the SCN. Through this influence of light, the circadian rhythm is synchronized with the external light-dark cycle and the peak in melatonin levels occurs during the night (69,70); this nocturnal peak in melatonin levels is reduced or even absent in cluster headache patients during an attack period (71–73) and returns during remission periods (71). Moreover, melatonin supplements have been reported to reduce the frequency of attacks in episodic cluster headache patients, although the therapeutic benefit was somewhat limited (74,75). In addition, cortisol concentrations are elevated during cluster headache attack periods (76–79). Finally, a nocturnal peak in prolactin levels often coincides with cluster headache attacks, whereas prolactin levels are low during remission periods (80).
Although cluster headache is generally 2.5–3.5-fold more prevalent among males than among females (2), the clinical symptoms are similar between men and women (48,81). The age of onset for cluster headache ranges from 11 to 40 years, and the prevalence generally decreases with age (48,50). The gender-biased nature and age dependency suggest that sex hormones may play a role in the pathophysiology of cluster headache, although the precise role remains poorly understood. Testosterone levels are often lower in patients during a cluster headache period (76,79,82–85); this effect may be secondary to disrupted REM sleep and/or increased cortisol levels during a cluster headache period (86,87). With the exceptions of FSH and LH, as discussed above, no other changes in sex hormones have been reported in cluster headache patients(79,84), and other factors that affect hormonal status such as oral contraceptives, menstruation, pregnancy, and menopause have only a small influence on cluster headache attacks (88).
The neuropeptides hypocretin-1 and hypocretin-2 play a key role in regulating sleep, maintaining homeostasis, controlling arousal, and processing pain (89). Interesting, the levels of HCRT-1 in the CSF are lower in cluster headache patients compared to controls (63), although these lowered levels are not associated with the self-reported rhythmicity of cluster headache attacks (5). A meta-analysis of genetic association studies revealed that the missense SNP rs2653349 in the HCRTR2 gene (which encodes HCRT receptor 2) is associated with a higher risk of cluster headache (90); however, a more recent analysis failed to reproduce this association, indicating that the previous conclusion should be interpreted with caution (91). Nevertheless, the putative role of the HCRT receptor remains interesting given the episodic nature of cluster headache.
In summary, several hormones and the neuropeptide hypocretin have been associated with cluster headache. However, whether changes in these compounds correspond with changes in their rhythmicity, thus reflecting a change in the SCN clock and/or its output, remains unknown. Alternatively, the altered sleep patterns and stress experience by these patients may explain the observed hormonal changes.
Circadian clock and cluster headache
The daily rhythmicity in the activity of the SCN follows a sinusoidal waveform due to the combined electrical activity of many SCN neurons that are synchronized but not in perfect phase with one another (92). The higher number of active neurons during the daytime creates a peak in electrical activity at midday and a trough during the night (Figure 2). This pattern of electrical activity is directly correlated with behavioral activity patterns (32).
The central pacemaker in mammals. (a) A coronal mouse brain section stained with Nissl, showing the location of the suprachiasmatic nuclei (SCN) and optic chiasm (OX). (b) Schematic diagram depicting the electrical activity of individual SCN cells (shown as blue traces) over a 24-hour period. The SCN cells oscillate slightly out of phase and give rise to the ensemble population pattern shown in black. This ensemble signal is transmitted to other areas of the brain, thereby providing rhythmicity to the central nervous system.
The biological clock can play a role in both the pathogenesis and clinical manifestation of various diseases and conditions. For example, the rhythmic output of the SCN can influence certain bodily functions, thereby altering the risk of certain diseases. In some cases, the clock can affect disease severity without playing a causal role; on the other hand, a malfunction in the clock’s activity can disrupt the synchrony of cellular/molecular rhythms in peripheral tissue, giving rise to various diseases such as metabolic disorders, cardiovascular disease, depression, and immune disorders (6). The circadian system may also play a causal role in cluster headache, particularly given the daily and seasonal pattern of the attacks and given that the SCN is located near the hypothalamus, which is believed to play a role in the onset and/or modulation of cluster headache attacks.
Seasonal rhythmicity and cluster headache
More than half of cluster headache patients report that their attacks have an annual rhythm (23). Typically, cluster headache is reported to follow an annual pattern with a peak in the spring (March–April) and a second peak in autumn (September–October) (3,4,50,81,93). However, there are some differences in the outcomes of these studies that seem to depend on the location of the observed patients. The studies set in California and in the US show the highest occurrence when days are shortening or lengthening (3) or shortest and longest (4). In Norway and Denmark, the lowest incidence systematically occurred when the days were longest (5,93). A study set in Taiwan showed an inverse correlation between daily sunlight (rather than daylight, which was not investigated) hours and incidence of cluster headache periods (94). These findings suggest that the occurrence of cluster headache attacks is related to the seasonal changes of daily sunlight. The seasonal rhythm of cluster headache attacks is more apparent in populations living further from the equator, where seasonal rhythms in daylight are stronger.
The biological clock and seasonal rhythmicity
The synchronization of neurons in the SCN appears to play an important role in encoding seasonal rhythms. For example, long days tend to reduce this synchronisation, resulting in a lower but broader ensemble peak; in contrast, shorter days increase synchronization, leading to a narrower, higher ensemble peak (Figure 3) (95). The width of the ensemble peak is therefore an internal reflection of the length of the environmental day (96), and a change in the width of the peak is the major mechanism by which the photoperiod, and thus the season, is encoded by the SCN (96). Certain diseases and conditions can also have seasonal fluctuations; for example, seasonal affective disorder is characterized by seasonal periods of depression that generally start in autumn/winter and resolve in the spring.
Seasonal synchronization of SCN activity. The electrical activity of individual SCN cells is shown in blue, and the ensemble activity of the SCN is shown in black. (a) During winter, the reduced amount of daily sunlight results in more synchronized firing, giving rise to a narrow, high-amplitude peak. (b) During summer, the increased amount daily sunlight results in less synchronized firing, giving rise to a broader peak with a lower amplitude.
Seasonal changes in hormone levels
In addition to daily rhythms, many hormones also follow a seasonal rhythm. For example, in mammals, melatonin levels – which are influenced by the SCN – encode information regarding day length by having a broader nighttime peak when days become shorter (97). In many mammals that breed in specific seasons, this information also affects the production of gonadotropic hormones such as FSH and prolactin (98). Studies in Antarctica showed that the nocturnal peak in melatonin production is delayed in the winter compared to the summer (99); in contrast, such a delay was not found in subjects living in Washington, DC; one possible explanation for this difference may be that the delay in peak melatonin levels could be suppressed by the presence of artificial lighting (100).
Lastly, analyses of the annual rhythm of melatonin revealed lowered concentrations of melatonin in the urine of cluster headache patients compared to control subjects (101). On the other hand, the concentration of melatonin was not influenced by the actual occurrence of attacks in patients (101).
The threshold hypothesis
The studies described above provide a plethora of clues suggesting that the biological clock plays a role in the occurrence of cluster headache attacks. Based on these data, we propose a threshold-based hypothesis to explain the temporal patterns of these headaches. Both chronic and episodic cluster headache appear to have a fluctuating rhythmicity, resulting in fluctuations in the probability of a cluster headache attack occurring at a given time, with high probability during a cluster and low probability outside the cluster. In patients with chronic cluster headache, this process follows a rhythmic sinusoidal pattern consisting of more active and less active periods (Figure 4, bottom). On the other hand, patients with episodic cluster headache have a similar sinusoidal pattern, with cluster headache attacks interspersed with periods of remission, with a threshold demarking the transition between these periods; when this threshold is exceeded, the brain enters a state in which the headaches can occur (Figure 4, top).
A hypothetic model to give an overview of factors influencing the disease process of cluster headache. Left: The threshold hypothesis explains the difference between episodic and chronic cluster headache. Top left: In episodic cluster headache, a fluctuating disease process causes cluster headache attacks only when the probability exceeds a predetermined threshold (depicted by the horizontal red line). Bottom left: In chronic cluster headache, the disease process always exceeds this threshold. Right: Central to this hypothesis is the notion that this threshold can be adjusted by various factors, including the circadian clock, sleep state and hormonal changes.
We hypothesize that a variety of factors, including external light, sleep, and the biological clock, can change the brain’s state, thereby lowering the threshold for activating the trigeminal-autonomic reflex. With the exception of the temporal differences in occurrence, the clinical and demographic characteristics of episodic and chronic cluster headache are strikingly similar. Thus, although episodic and chronic cluster headache are likely caused by the same pathophysiological processes, a more “permissive” brain state leads to chronic cluster headache. Based on this theoretical model, such a threshold could be regulated by factors such as the circadian clock and sleep state.
Conclusions
Cluster headache has a clearly established daily rhythm, with the frequency of attacks peaking during the night; the seasonal pattern of cluster headache is also rhythmic but is more variable. These patterns are likely driven either directly or indirectly by the biological clock. Changes in the daily rhythmicity of hormones involved in cluster headache have been reported, although the reports are not consistent. In addition, a relationship between REM sleep and cluster headache attacks has been reported, but the evidence for this relationship is not compelling and is also not consistent. One explanation for the seasonal rhythmicity of cluster headache attacks is the occurrence of the summer and winter solstice and the March and September equinoxes, as well as the amount of daily sunlight. We propose that episodic and chronic cluster headache have similar pathophysiological mechanisms, and changes in the brain’s state can reduce the threshold for activating the trigeminal-autonomic reflex, thereby giving rise to the chronic form of cluster headache. An important implication for the threshold hypothesis is that chronic cluster headache can turn into episodic cluster headache (and vice versa) in a given patient. We also suggest that various factors, including sleep, light, and particularly the biological clock, can influence the threshold for experiencing a cluster headache attack. Future studies should be designed to help elucidate the relative contributions of these putative factors to the susceptibility of cluster headache attacks. The insights gained from these studies would provide an important starting point for more mechanism-based studies of cluster headache and would likely help facilitate the development of new therapeutic strategies.
Abbreviations
Calcitonin gene-related peptide
Circadian locomotor output cycles kaput
Functional magnetic resonance imaging
Hypocretin
Hypocretin receptor 2
Intrinsically photosensitive retinal ganglion cells
Magnetic resonance angiography
Pituitary adenylate cyclase-activating peptide
Positron emission tomography
Retinohypothalamic tract
Suprachiasmatic nucleus
Sex hormone-binding globulin
Single-nucleotide polymorphism
Thyrotropin-releasing hormone
Vasoactive intestinal peptide
Article highlights
The biological clock plays a role in both the occurrence and severity of cluster headache attacks. Although sleep has been reported as a trigger for cluster headache attacks, the precise relationship between sleep stage and cluster headache is currently unknown. The seasonal pattern of cluster headache attacks may be influenced by the amount of daily sunlight and appears to vary with latitude. The precise mechanistic role that the circadian clock plays in cluster headache requires further study in order to facilitate the development of more effective therapeutic approaches. On the basis of the “threshold hypothesis”, we propose that the same mechanism accounts for episodic and chronic cluster headache.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.