
Abstract
Carnivore protoparvovirus-1 (CPPV-1) infection has been reported frequently in both domestic and wildlife species including wild carnivores. Fifty-five captive small Indian civets (Viverricula indica), farmed for perfume production in Eastern Thailand, showed clinical signs of acute bloody diarrhea, anorexia, vomiting, circling, and seizures. The disease spread within the farm and resulted in the death of 38 of the 55 civets (69% mortality) within a month. Fecal swabs were collected from the 17 surviving civets, and necropsy was performed on 7 of the dead civets. Pathologic findings were severe hemorrhagic gastroenteritis with generalized lymphadenopathy. CPPV-1 was identified in both fecal swabs and postmortem samples by species-specific polymerase chain reaction. Further whole-gene sequencing and restriction fragment length polymorphism analysis suggested feline panleukopenia virus (FPV) as the causative agent. The viral tropism and tissue distribution were confirmed by immunohistochemistry, with immunolabeling in the cytoplasm and nucleus of small intestinal crypt epithelial cells, villous enterocytes, histiocytes in lymphoid tissues, myenteric nerve plexuses, and cerebral and cerebellar neurons. Phylogenetic analysis of civet-derived CPPV-1 indicated a genetic similarity close to the FPV HH-1/86 strain detected in a jaguar (Panthera onca) in China. To our knowledge, this mass die-off of civets is the first evidence of disease associated with CPPV-1 infection in the subfamily Viverrinae. These findings support the multi-host range of parvovirus infection and raises awareness for CPPV-1 disease outbreaks in wildlife species.
Keywords
Carnivore protoparvovirus-1 (CPPV-1), belonging to the genus Protoparvovirus, family Parvoviridae, is one of the most contagious pathogens causing fatal disease in dogs, cats, and other wild carnivores worldwide. According to the International Committee on Taxonomy of Viruses report in 2019, this species contains 2 parvoviruses that cause severe hemorrhagic gastroenteritis in both cats and dogs, known as feline panleukopenia virus (FPV) and canine parvoviruses (CPVs), respectively. Previously, it was believed that FPV and CPVs infected restricted hosts. However, it has recently been reported that both viruses can cause serious illness in other carnivores. Thus, CPPV-1 was tentatively designated as a causative parvoviral agent in carnivores. 1 –3,11
CPPV-1 is a nonenveloped single-stranded DNA virus with an approximately 5 kb genome size containing 2 open reading frames encoding 2 nonstructural proteins (NS1 and NS2) and 2 capsid proteins (VP1 and VP2). Several mutations of amino acid residues in the VP2 protein play a central role in the susceptible host selection in domestic animals and expand infectivity to other wild carnivores. 1,17,18,25,27,28 Although mutations in the VP2 gene have been a basis for variant classification and understanding the potential host-dynamic range, mutations in the NS1 region of the FPV isolated from domestic cats have been recently considered as potential points for pathological insights. 6 Moreover, the NS1 of CPPV-1 is now important for molecular epidemiological investigations. 15 However, the information regarding both NS1 and VP2 mutations of CPPV-1 isolated from wild carnivores is limited.
The FPV, a member of CPPV-1, was reported in wild felids, such as American tiger cat (Felis tigrina), African tiger cat (Felis aurata), rusty tiger cat (Felis planiceps), marbled cat (Felis marmorata), caracal lynx (Felis caracal), ocelot (Felis pardalis), and leopard (Panthera pardus) in the 1930s 8 and since then has been reported in various individual wildlife carnivore species, including mink (Mustela vision), American river otter (Lontra canadensis), skunk (Mephitis mephitis), raccoon (Procyon lotor), blue fox (Alopex lagopus), grey wolf (Canis lupus), coyote (Canis lupus), dingo dog (Canis latrans), Asiatic raccoon dog (Nyctereutes procyonoides), Arctic fox (Vulpes lagopus), European red fox (Vulpes vulpes), cheetah (Acinonyx jubatus), Siberian tiger (Panthera tigris altaica), stone marten (Martes foina), and bat-eared fox (Otocyon megalotis). 24
Focusing on CPPV-1 infection in the Viverridae family of small- to medium-sized mammals, evidence of seropositivity for CPPV-1 in viverrids was initially documented in Formasan gem-faced civets (Paguma larvata taivana) in Taiwan. 10 Thereafter, periodic CPPV-1 infection was reported individually in the 2 subfamilies of viverrids of Paradoxurinae and Genettinae. 4,5 However, evidence of this CPPV-1 infection and associated in another viverrid subfamily, Viverrinae, has never been reported. This study describes a large fatal outbreak in captive small Indian civets (Viverricula indica).
Materials and Methods
Clinical History and Sample Collection
In September 2013, 2 out of 55 captive small Indian civets (Viverricula indica), farmed for perfume production in Thailand, died after a short course of acute bloody diarrhea, anorexia, vomiting, dehydration, circling, and seizures. Thereafter, the disease spread to 36 of the other 53 captive civets, resulting in their death within a month. Fecal swabs were collected from the 17 surviving civets (cases 1–17), using sterile Rayon Tipped Applicator (Puritan). The swabs were placed in 1% (v/v) sterile phosphate-buffered saline (PBS) and kept at −80 °C for molecular diagnosis. Essential data were recorded, including clinical signs, farm management, and biosecurity practices. Seven dead civets (cases 18–24) were submitted to the Department of Pathology, Faculty of Veterinary Science, Chulalongkorn University, for pathological study. All experimental protocols were approved by the Chulalongkorn University Animal Care and Use Committee (No. 11310088).
Pathological Examination
Gross lesions of the 7 necropsied civets (cases 18–24) were evaluated by a veterinary pathologist (WB). Various organs, including the heart, lung, liver, kidney, small intestine, large intestine, cecum, rectum, mesenteric and peripheral lymph nodes, thymus, genital organs, and brain, were collected and preserved in 10% (v/v) neutral buffered formalin and routinely processed for histopathological examination.
Polymerase Chain Reaction (PCR) and RFLP Analysis
Eighteen fecal swabs (cases 1–17, 24) were subjected to viral genomic extraction using a viral DNA/RNA extraction kit II (GeneAid, Taipei, Taiwan). The VP1/VP2 gene encoding for the capsid proteins of CPPV-1 was amplified using primers VPF and VPR (Supplemental Table S1) as previously described 16 with minor modifications. Briefly, the PCR thermal cycling was performed as 30 cycles of 94 °C for 30 seconds, 55 °C for 2 minutes, and 72 °C for 2 minutes. The 2246 bp PCR products were resolved by 1.5% (w/v) agarose-gel electrophoresis and visualized under UV transillumination after ethidium bromide staining, and then subjected to RFLP analysis to identify the CPPV-1 as FPV or CPV. Amplicons were digested with HincII restriction enzyme (New England BioLabs, Hitchin, UK) for differentiation between wild-type FPV and CPVs as previously described. 16 The digested fragments were resolved by 1.5% (w/v) agarose gel electrophoresis. FPV and CPV-2 isolates from fecal samples of an infected cat and an infected dog, respectively, with confirmed sequences served as the positive controls. Distilled water instead of DNA template was used as negative control.
To further differentiate between the virus in these civets and vaccine strains given to the nearby client-owned cats, separate PCR reactions and RFLP analyses were performed. Civet viral samples and the FPV-commercial vaccine strains Felocell (Zoetis) and Purevax (Merial) were PCR-amplified by primers specific to the 1755 bp of VP2 gene (P5 and P6, Supplemental Table S1) as reported, 20 with thermal cycling performed as above. The PCR products were digested with HinP1I (New England BioLabs, Hitchin, UK) and resolved by agarose gel electrophoresis as above using a positive FPV (mentioned above) and the available FPV vaccines (Purevax and Felocell) as positive controls.
The complete nonstructural NS1 and NS2 genes were amplified from the dead civet cat that had shown neurological signs of circling and seizures (case 24). The PCR amplification was performed in separate reactions using previously described primer pairs for NS gene (Supplemental Table S1). 22 Briefly, PCR thermal cycling was performed as 30 cycles of 94 °C for 30 seconds, 55 °C for 1 minute, and 72 °C for 1 minute.
Other common canine/feline viral pathogens, including canine distemper virus (CDV), canine parainfluenza (CPIV), canine herpesvirus-1 (CaHV-1), canine adenovirus-1 and -2 (CAdV-1/-2), and feline and canine enteric coronaviruses (FCoV/CCoV) were also tested using civet fecal samples by specific PCRs as described previously. 13,21,23
Genomic Sequencing, Phylogenetic and Recombination Analysis
Eight VP1/VP2 PCR-positive samples derived from fecal swabs of cases 2, 4, 8, 10, 11, 13, 14, and 24 were further sequenced for genome analysis. Multiple PCR reactions with different primer pairs (Supplemental Table S1) were used for the complete genetic characterization of the civet CPPV-1 VP gene as described. 16 Moreover, the civet cat that showed neurological signs (case 24) was additionally sequenced for the complete NS gene (as mentioned above). All PCR products were purified using NucleoSpin Extract II (Macherey-Nagel, Düren, Germany) and submitted for commercial genetic sequencing (SolGent Ltd, Daegeon, South Korea). Nucleotide alignments and deduced amino acids were analyzed and compared with published data deposited in GenBank using Bioedit Sequence Alignment Editor Version 7.2.5.
The phylograms were analyzed using the MEGA 7.0 software (http://www.megasoftware.net/). Phylogenetic trees were constructed by the maximum likelihood (ML) method and bootstrap analysis with 1000 replicates was used to assess the confidence level of the branch pattern. Bootstrap values of >70% were considered to be significant. To detect possible genetic recombination event(s) in these CPPV-1 strains, recombination analysis was conducted with CPPV-1 sequence derived from the infected civet (case 24), and CPPV-1 sequences available in GenBank served as sequence queries. A cocktail of statistical methods, including RDP, GENECONV, BootScan, MaxChi, Chimaera, SiScan, and 3Seq were used with default settings in the Recombination Detection Program (RDP) package, version 4.0, to check for potential recombination events. Any potential breakpoint signals revealed by at least 4 methods with p values <.01 were considered to gain a potentially positive recombination. 23
Immunohistochemistry (IHC)
To localize CPPV-1 antigen in formalin-fixed paraffin embedded tissue, 3 necropsied civets (cases 18, 19, and 24) were used for IHC investigation using the EnViSion system horseradish peroxidase labeled with polymer anti-mouse (EnVision polymer, Dako, Glostrup, Denmark). Sections were cut at 4-µm thickness, deparaffinized, and hydrated. They were then pretreated with trypsin for 30 minutes. Endogenous peroxidase and nonspecific reactions were blocked by incubation with 3% (w/w) hydrogen peroxide and 5% (w/v) skim milk in 1% PBS, respectively. Subsequently, sections were incubated with mouse monoclonal anti-canine parvovirus antibody (1:250 dilution in 5% skim milk) to detect the CPVs, mink enteritis virus (MEV), and FPV (ab59832, Abcam, Cambridge, UK), at 4 °C overnight, washed 3 times in 1% PBS, and then sections were incubated with the EnViSion system as per the manufacturer’s instructions before being immersed in 3,3′-diaminobenzidine and counterstained with Mayer’s hematoxylin before mounting. Sections of the intestinal tissue of confirmed FPV-infected cats were used as a positive control, while sections from the same source but omitting primary monoclonal antibody (replaced with distilled water) was applied as a negative control. Immunopositive cells were noted for interpretation of cellular tropism and viral distribution in organs. Immunoreactivity was scored, semiquantitatively analyzed, and averaged in 5 different areas under high-power field (400×) as follows: − (no immunopositive cells); + (≤25% immunopositive cells); ++ (26% to 50% immunopositive cells); +++ (51% to 75% immunopositive cells); ++++ (≥76% immunopositive cells).
Results
Clinical Findings
All captive civets in this study were small Indian civets, Viverricula indicia [Viverridae: Viverrinae]. The farm was located in a rural area of Trat province, Eastern Thailand, and close to the Thailand-Cambodia border. Farm biosecurity was rather poor, as seen by the open and unfenced border. Animals living around this farm such as domestic dogs and cats could freely access the housed civet cat area, and there was no disinfection area prior to enter the animal’s housing. Each captive civet was raised individually in a ventilated wood lathe cage. Of the 55 housed civets, 38 died (69%) within a month of initial farm infection and had clinical signs of bloody diarrhea. Some civets showed signs of circling and seizure. Clinical signs of surviving civets are shown in Supplemental Table S2.
Pathological Findings
The 7 necropsied civets (cases 18–24) were markedly emaciated with pale mucous membranes and severe dehydration. All 7 civets had remarkable macroscopic lesions of segmental hemorrhagic enteritis containing a watery dark-brown content in the lumen (Fig. 1; Supplemental Table S3). Hematopoietic organs including spleen and mesenteric lymph nodes were enlarged and congested, and there were foci of thymic hemorrhage in 2 civets. Marked congestion was observed in the brain, lung, and liver. One dead civet (case 24) was promptly tested with rapid test kits for CDV/CPV/CCoV antigen (Bionote, Gyeonggi-do, Republic of Korea) using a fecal sample, and showed a strong positive result only for CPV (data not shown).
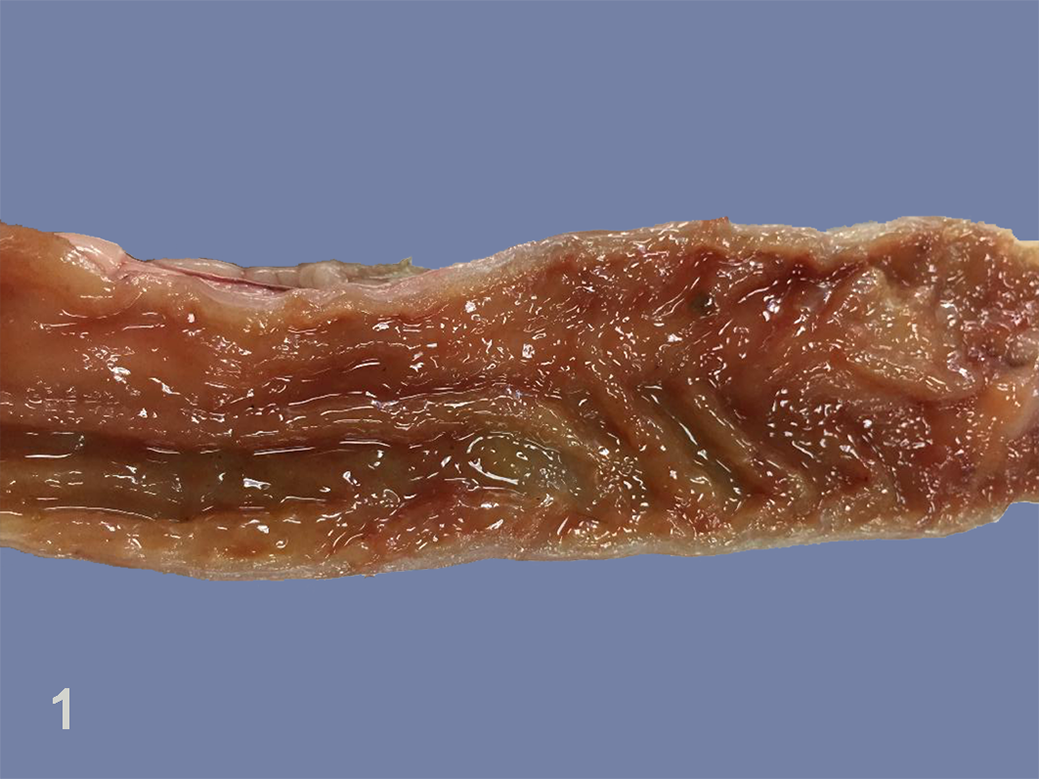
Carnivore protoparvovirus infection, jejunum, small Indian civet, case 24. There is reddening of the mucosal surface.
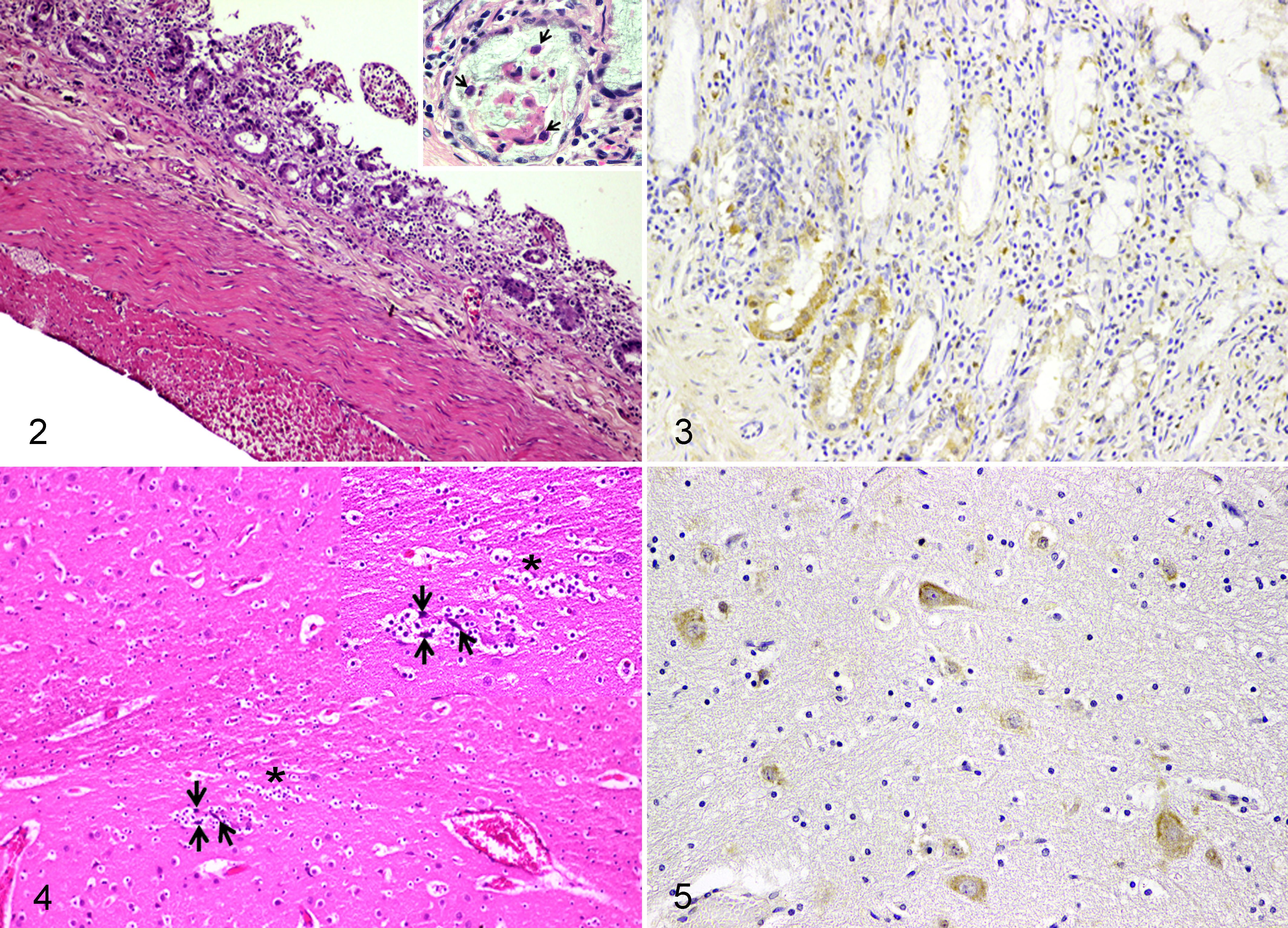
Histopathological findings in the small intestine revealed severe diffuse desquamative hemorrhagic enteritis, characterized by atrophied and fused villi, crypt necrosis, and lymphoplasmacytic infiltration in the lamina propria (Fig. 2). All 7 examined civet cats showed numerous amphophilic intranuclear inclusion bodies in small intestinal crypt epithelial cells and villous enterocytes (Fig. 2, inset; Supplemental Table S4). There was severe lymphoid depletion in the Peyer’s patches of the ileum, colonic lymphoid follicles, mesenteric lymph nodes, and spleen in all 7 examined cases. For the central nervous system, the meninges and choroid plexus were moderately congested. There was neuronal degeneration with satellitosis in the cerebral cortex while there were no lesions in white and gray matters of cerebellum (Fig. 4).
PCR and RFLP Results
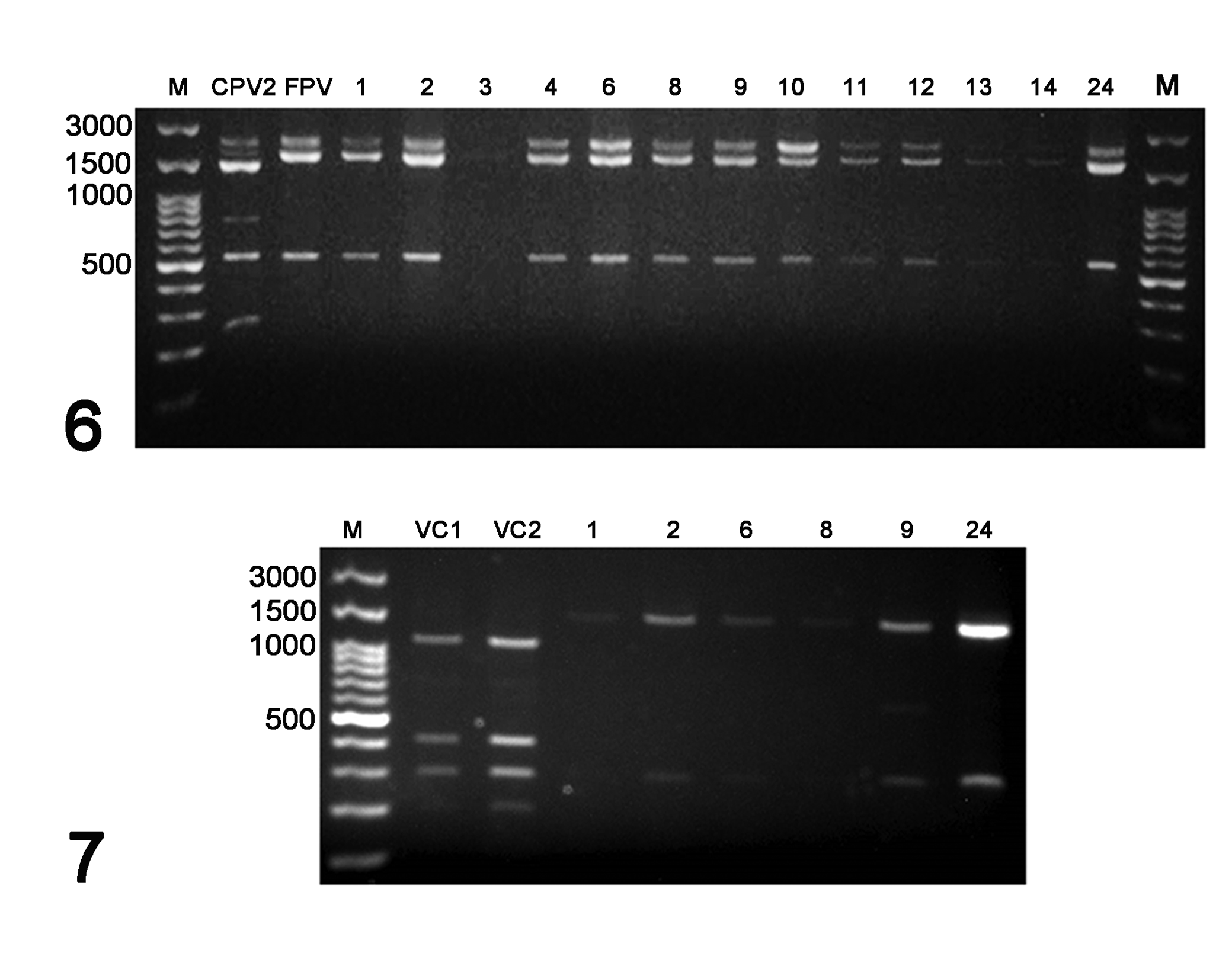
Out of 18 fecal swab samples (cases 1–17 and 24), 13 (72%) had positive 2246 bp PCR products for the VP1/VP2 gene of CPPV-1. All were negative using PCR assays specific for CDV, CPIV, CaHV-1, CAdVs, FCoV, and CCoV (Supplemental Table S2). The 2246 bp VP1/VP2 amplicons from each of the 13 CPPV-1 positive civets were subjected to RFLP analysis using the HincII restriction enzyme. This generated 535 bp and 1711 bp fragments in 12 of 13 tested civet samples (92.3%), with the same as the FPV positive control sample, whereas the CPV-positive control displayed 3 fragments (242 bp, 535 bp, and 1469 bp). One civet sample (case 3) was RFLP-negative showing no HincII restriction pattern (Fig. 6). However, this sample was not sequenced to confirm the absence of a HincII restriction site (rather than a failed digestion). Similarly, the 1755 bp amplicons of the CPPV-1 VP2 gene were digested with HinP1I, revealing 2 fragments (248 bp and 1507 bp) in the civet samples, with the same as the wild type FPV, while the vaccine strains presented 3 fragments (248 bp, 417 bp, and 1090 bp; Fig. 7).
Viral Genome Analysis
The VP2 gene was further sequenced for 7 cases that had positive VP1/VP2 amplicons (cases 2, 4, 8, 10, 11, 13, 14), while one other (case 24) was further sequenced for the full-length CPPV-1 genome. The obtained sequences have been submitted to the GenBank database with accession nos. KP019617 to KP019620 for cases 2, 8, 10, and 13, respectively; KT357491 to KT357493 for cases 4, 11, and 14, respectively; plus KP019621 for the full-length genome from case 24 (Supplemental Table S2). The civet-derived nucleotide sequences and deduced amino acids were compared to other CPPV-1-annotated sequences retrieved from the GenBank database.
The full-length genome of the civet CPPV-1 (case 24) shared 99.4% nucleotide similarity with the FPV HH-1/86 strain from a jaguar (Panthera onca) in China (accession no. KX900570). For individual gene analysis, civet CPPV-1 from this study shared 99.7%, 100%, and 99.1% nucleotide similarity to the NS1, NS2, and VP1 gene sequences of the jaguar FPV HH-1/86 isolate, respectively.
For the in silico deduced amino acid analysis, the civet CPPV-1 (case 24) revealed 99.7% and 99.4% amino acid similarity to NS1 of the jaguar FPV HH-1/86 and the FPV GX01 strain isolated from domestic cat (accession no. MG924893), respectively. For the amino acid similarity to NS2, this civet CPPV-1 demonstrated 98.8% similarity to both jaguar FPV HH-1/86 and the cat FPV GX01 (data not shown), while the VP2 showed 100% amino acid similarity to the FPV lineage at specific amino acid residues (Table 1). Interestingly, the civet CPPV-1 had a distinct amino acid mutation, G573S, in the NS1 gene, that was not observed in other FPV isolates, but this mutation was present in some CPV-2a (accession no. AY742936) and CPV-2c (accession no. MH476584) isolates (Table 1).
Host-Specific Amino Acids of the Viral NS1 and VP2 Proteins for Carnivore Protoparvovirus-1 Viruses Analyzed in This Study.
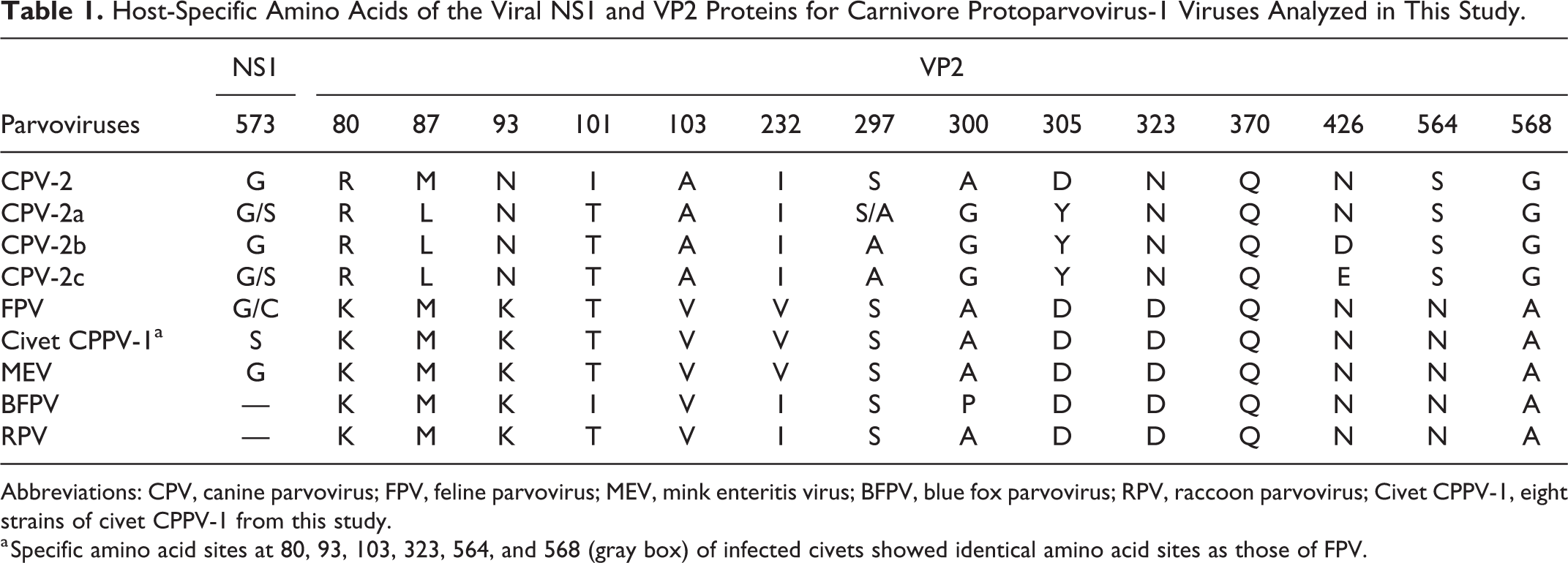
Abbreviations: CPV, canine parvovirus; FPV, feline parvovirus; MEV, mink enteritis virus; BFPV, blue fox parvovirus; RPV, raccoon parvovirus; Civet CPPV-1, eight strains of civet CPPV-1 from this study.
a Specific amino acid sites at 80, 93, 103, 323, 564, and 568 (gray box) of infected civets showed identical amino acid sites as those of FPV.
Phylogenetic analysis of NS1, VP2, and whole genome sequences of civet CPPV-1 depicted a distinctively monophyletic branch in the FPV lineage, which was separated from the other CPV (and MEV) lineages (Supplemental Figs. S1-S3). The phylogram of reported CPPV-1 infections in animals of the family Viverridae is depicted in Figure 8, including a related CPPV-1 from an Asian palm civet (subfamily Paradoxurinae) in Hungary in 2008.
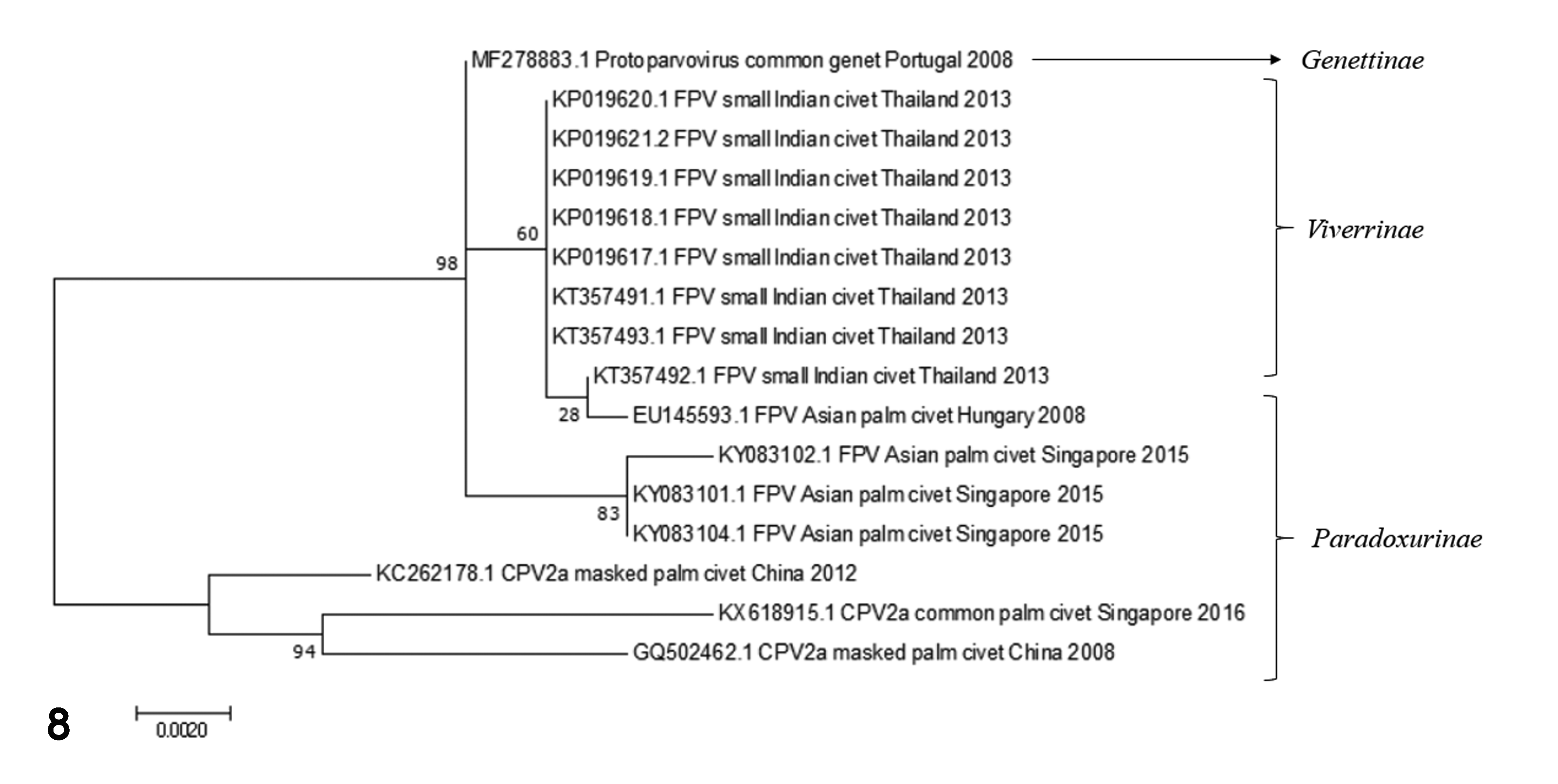
Phylogram of CPPV-1 (FPV and CPV) infection in family Viverridae (including the subfamilies Viverrinae, Paradoxurinae, and Genettinae). The phylogenetic tree was constructed in MEGA 7.0 using the ML method with 1000 bootstrap replications. GenBank accession numbers are present on the tree. Bar indicates the estimated numbers of nucleotide substitutions per site.
To investigate the possibility of genomic recombination in these CPPV-1-infected civets, the potential recombination event(s) in the CPPV-1 VP2 gene of the 8 sequences obtained from the different infected civets in this study and the other CPPV-1 sequences used for the phylogenetic analysis were analyzed using the RDP software. However, no recombination breakpoints were found in any of the civet CPPV-1 sequences (data not shown).
Cellular Tropism and Tissue Localization
Three necropsied civets (cases 18, 19, and 24) were analyzed using IHC to determine the localization of CPPV-1 antigen. Immunolabeling was observed in both the cytoplasm and nucleus of the villous enterocytes, crypt epithelial cells, and myenteric ganglionic neurons, which was compatible with the observed abundant infiltration of mononuclear inflammatory cells in the mucosa of the small intestine (Fig. 3). The infiltrated histiocytes in the spleen and mesenteric lymph node also showed intense immunolabeling. In the central nervous system, neurons in the cerebral cortex and cerebellar Purkinje cells showed strong immunoreactivity in their cytoplasm (Fig. 5). The IHC scores are shown in Supplemental Table S5.
Discussion
The fatal gastroenteritis disease caused by protoparvovirus infection in dogs and cats is common, and it is sporadically reported in wild carnivores. Unvaccinated free-roaming dogs and cats may serve as potential hosts for the outbreak. Unfortunately, samples of the free-roaming domestic and farmed animals that had access to this Indian civet cat farm were not available for evaluation of their infection status.
The CPV and FPV are members of the contagious CPPV-1 virus group in which VP2, the main structural protein, plays a major role in infection in various animals. Therefore, recent studies of CPPV-1 genome have focused on amino acid mutations in the VP2 gene. In this study, 6 mutated amino acid residues of VP2 of civet CCPV-1 (sites 80, 93, 103, 323, 564, and 568) were observed that can differentiate FPV from CPV isolates. 28 Furthermore, a deduced amino acid mutation (G573S) in the NS1 gene of civet CPPV-1 was observed, that was distinct from the amino acid sequence of FPV. Therefore, civet CPPV-1 was tentatively named as a potentially novel virus detected in this outbreak. Nevertheless, a small number of the observed sequences did not have this mutation. Thus, the relationship of CPPV-1 to FPV needs further clarification.
The obtained sequences demonstrated a high nucleotide similarity to other FPV isolates and the phylogenetic analysis revealed that civet CPPV-1 grouped with FPV in a monophyletic lineage that was separate from CPV isolates. This supports the conclusion that CPPV-1 detected in this civet cat outbreak was of FPV origin, which is also supported by the RFLP analysis. The use of RFLP is useful for distinguishing origins of CPPV-1. However, one sample (case 3) that was PCR positive for the VP2 gene revealed no digestion in the RFLP assay. Although not sequenced to confirm the presence or absence of the HincIII restriction enzyme sequences(s), this may reflect a technical failure of the digestion due to the lower yield of DNA sample, 7 or to enzyme inhibitors.
The origin of the viral carrier in this outbreak remained unclarified. It might have occurred from civet-spread parvovirus itself or been from other animal reservoirs living in the area surrounding this farm. To date CPPV-1 detection in Viverridae has been reported in the subfamilies Paradoxurinae and Genettinae, 4,5,30 while CPPV-1 isolated in this study, named as civet CPPV-1, is the first reported infection in a member of the subfamily Viverrinae.
From the pathological studies, CPPV-infected civets showed lesions similar to parvovirus infection, including severe acute segmental muco-hemorrhagic enteritis with collapsed, blunted villi, and necrosis and inflammation of intestinal crypts. Moreover, viral amphophilic intranuclear and intracytoplasmic inclusion bodies were abundant in enterocytes and cryptal epithelial cells, in accordance with previous findings in parvovirus-infected dogs and cats. 9,14,19,26
The IHC analysis also supported the parvovirus infection and indicated its localization. Interestingly, immunopositive cells were present not only in intestinal crypts but also in cerebral neuronal cells and cerebellar Purkinje cells. There are some previous reports of CPPV-1 localization in the brain tissue in companion animals. 12,29 The mechanism of neuron invasion of the FPV-infected cat was related to the L582S mutation of NS1, changing residue 582 from leucine to serine. 6 However, this specific mutation was absent in CPPV-1, even though the neuronal cells were immunolabeled for parvoviral antigen. These discrepant findings warrant further investigation, due to the different affected hosts, using a large-cohort study.
In conclusion, we report here the first identification of CPPV-1 infection in small Indian civet cats (Viverricula indica), as evidenced by pathological, immunohistochemical, and molecular investigations. These results extend the multi-host range of parvovirus and emphasize the pathological role of CPPV-1 in a new mammalian host.
Supplemental Material
Supplemental Material, Combined_supplemental_materials-Chaiyasak_et_al - Carnivore Protoparvovirus-1 Associated With an Outbreak of Hemorrhagic Gastroenteritis in Small Indian Civets
Supplemental Material, Combined_supplemental_materials-Chaiyasak_et_al for Carnivore Protoparvovirus-1 Associated With an Outbreak of Hemorrhagic Gastroenteritis in Small Indian Civets by Surangkanang Chaiyasak, Chutchai Piewbang, Wijit Banlunara and Somporn Techangamsuwan in Veterinary Pathology
Footnotes
Acknowledgements
We are thankful to Dr Piyaporn Kongmakee for excellent coordination with the Monitoring and Surveillance Center for Zoonotic Disease in Wildlife and Exotic Animals (MoZWE) of Mahidol University, Thailand, and the Zoological Park Organization under The Royal Patronage of H.M. The King, Thailand.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: S. Chaiyasak was granted by Chulalongkorn University tuition fee and Mahasarakham University domestic tuition fee. C. Piewbang was supported by Ratchadapisek Somphot Fund for Postdoctoral Fellowship, Chulalongkorn University. This study was financially supported by the 90th Anniversary of Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund) and partially funded by the Veterinary Pathogen Bank, Animal Virome and Diagnostic Development Research Group, and Diagnosis and Monitoring of Animal Pathogens Research Unit, Faculty of Veterinary Science, Chulalongkorn University.
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.