
Abstract
Cardiovascular disorders and predominantly idiopathic myocardial fibrosis are frequently associated with mortality among zoo-housed chimpanzees (Pan troglodytes). Formalin-fixed whole hearts of deceased chimpanzees housed in zoos (n = 33) and an African sanctuary (n = 2) underwent detailed macroscopic and histopathologic examination using a standardized protocol. Archived histological slides from the hearts of 23 additional African sanctuary-housed chimpanzees were also examined. Myocardial fibrosis (MF) was identified in 30 of 33 (91%) of the zoo-housed chimpanzees but none of the 25 sanctuary-housed chimpanzees. MF was shown to be characterized by both interstitial and replacement fibrosis. Immunophenotyping demonstrated that the fibrotic lesions were accompanied by the increased presence of macrophages, alpha smooth muscle actin-positive myofibroblasts, and a minimal to mild T-cell-dominant leukocyte infiltration. There was no convincing evidence of cardiotropic viral infection or suggestion that diabetes mellitus or vitamin E or selenium deficiency were associated with the presence of the lesion. However, serum vitamin D concentrations among zoo-housed chimpanzees were found to be lower in seasons of low ultraviolet light levels.
Keywords
Cardiovascular disease has been cited as a significant cause of mortality among captive chimpanzees. 23,32,37,46,47 A chronic, degenerative heart disease characterized by fibrosis of the cardiac muscle is the predominant condition diagnosed on postmortem examination and is associated with sudden death presumably due to dysrhythmia. 11,24,48 As yet, however, the cause(s) of idiopathic myocardial fibrosis (IMF) in great apes (also referred to as interstitial myocardial fibrosis 23 and fibrosing cardiomyopathy 43 ) remains unknown.
Myocardial fibrosis (MF) is a nonspecific response to cardiac insult or injury and one of the most common histologic features of the diseased heart. In humans, there are 2 histological subtypes of MF, the pathophysiology of which differ. 28,39 Replacement fibrosis (RF) is the irreversible response to cardiomyocyte damage and loss, which occurs, for example, following myocardial infarction or myocarditis. Necrotic cardiomyocytes are removed and replaced by scar tissue to preserve structural and functional integrity of the myocardium. 15,52 In contrast, interstitial fibrosis (IF) can occur in the absence of cardiomyocyte damage or death and is an adaptive response that at least initially allows cardiac structure and function to be preserved. Its formation is typically reactive, occurring in response to underlying (sometimes remote) disease processes such as renal disease, hypertension, diabetes mellitus, primary cardiomyopathies, secondary pressure/volume overloading, and inflammation. 1,4,5,7,17,21,30,34,56 IF is also well-established as a feature of age-related remodeling of the human heart. 18 Irrespective of the cause, IF is characterized by excessive deposition of extracellular matrix (ECM) proteins by cardiac myofibroblasts, which expand the interstitial (and often also perivascular) space. 52 Importantly, IF is reversible, and given that its formation can precede that of the irreversible RF subtype, its extent and severity can also be used as a marker of disease progression. 29,31,39
Trauma and infectious disease are cited as the most frequent causes of death among wild chimpanzees, with MF being identified in only 2 (out of 11) chimpanzees from Gombe National Park, Tanzania between 2004 and 2010. 51 This raises the question as to whether the high prevalence of cardiovascular disease and MF seen in zoos and research laboratories is a feature and/or consequence of life in captivity. There are differences in the life expectancy and the infectious, dietary, and environmental factors to which wild versus captive chimpanzee populations are exposed, each of which require careful consideration and investigation. One such difference is their exposure to sunlight and therefore, presumably, their serum vitamin D levels. 10,54 An inverse relationship between serum vitamin D concentrations and cardiovascular disease risk is recognized in people, 42 and vitamin D has also been shown to have antifibrotic properties in the heart and other organs. 38 Other diseases and deficiencies relating to metabolic status and micronutrient levels in zoo-housed chimpanzees might also be significant. Type 2 diabetes has been diagnosed commonly in captive chimpanzee populations 36 and in humans is associated with a cardiomyopathy, which is characterized by myocardial fibrosis and dysfunctional remodeling. 1,40 Additionally, both vitamin E and selenium, which possess anti-oxidant properties, are believed to have protective effects against myocardial ischemic damage. 6,13,44 Deficiencies in both micronutrients have been implicated in the development of myocardial necrosis in various animal species, 14,22 and vitamin E deficiency specifically has been associated with cardiomyopathy in gorillas 35 and other zoo-housed primates. 26,27 Captivity is also associated with greater human-chimpanzee interaction than would occur in the wild. This combined with our genetic similarity suggests that captive chimpanzees are at risk of becoming infected with human pathogens. Inflammation (albeit mild) has been described as a feature of MF in chimpanzees and gorillas 2,9,24,43 and is recognized as being linked to cardiac fibrosis and cardiomyopathy in people. 12,41,50 The potential that IMF in chimpanzees might occur due to infectious myocarditis or viral persistence therefore also warrants exploration.
Until recently, much of the investigative work aimed at enhancing understanding about great ape cardiovascular disease has been carried out in North American zoos (driven by the Great Ape Heart Project, based at Zoo Atlanta) or US research facilities. In 2013, however, a European taskforce was formed, bringing together specialists from the fields of human and veterinary cardiovascular medicine and pathology. The Ape Heart Project is a collaborative initiative based at Twycross Zoo (United Kingdom), which aims to promote and drive the study of great ape cardiovascular disease across European zoological collections. Its work to date has resulted in mortality reviews for each of the great ape genera, 46,47 a retrospective study of great ape cardiovascular disease and epidemiology and pathology, 48 and the generation of guidelines for the consistent examination, sampling, and reporting of great ape cardiovascular lesions. 49
This study, a part of the Ape Heart Project, describes the cardiac lesions affecting European zoo-housed chimpanzees, characterizes chimpanzee IMF, and investigates the possible associated dietary, infectious, and environmental factors.
Materials and Methods
Cardiac Samples From African Sanctuary Chimpanzees
Archived hematoxylin eosin (HE)-stained histologic sections of the heart of 23 African sanctuary chimpanzees (8–27 years; median age = 12 years; 8 male, 15 female) were examined by a board-certified veterinary pathologist (K. Baiker). These slides were produced from small, often single-site myocardial (unspecified location) samples taken at postmortem examination at a single rescue and rehabilitation center in Sierra Leone, Africa. Two entire formalin-fixed hearts from the same African sanctuary (cases C12 and C13; 11 and 13 years; male) underwent detailed examination as described below. All animals from the sanctuary were housed in a large mixed indoor/outdoor forested enclosure and fed a diet comprised of local mixed vegetables, fruits, cereals, and nuts.
Cardiac Samples From European Zoo Chimpanzees
Hearts from European Association of Zoos and Aquaria (EAZA) member zoo-housed chimpanzees (n = 33; 9–65 years; median age = 32 years; 19 male, 14 female) (cases C1–C11, C14–C35) were collected postmortem, submerged in 10% neutral buffered formalin, and sent to the Ape Heart project for further investigation. Data relating to the age, sex, weight, and circumstances of death for these animals are presented in Supplemental Table S1. All hearts were fully submerged in 10% neutral buffered formalin for at least 72 hours prior to examination and sampling.
Detailed Cardiac Examination
Formalin-fixed entire hearts underwent detailed macroscopic examination and sampling according to a protocol that is based on guidelines for the investigation of sudden cardiac death in humans 3,45 and has been published for use in great apes. 49 A minimum of 12 cardiac samples were taken from predetermined and consistent locations, including myocardium from the anterior, posterior, and lateral left and right ventricular walls and anterior and posterior portions of the interventricular septal wall and right ventricular outflow tract. Samples of the aorta, sino-atrial nodal region, and atrioventricular nodal region were also taken. Additional tissue samples of anomalies or lesions detected as part of the examination were taken as indicated. Histologic sections were prepared from formalin-fixed paraffin-embedded blocks and routinely stained with HE (n = 35) and Masson’s trichrome to highlight collagenous connective tissue fibers (n = 13; C1, C3, C4, C7, C10, C12, C13, C15, C19, C20, C23, C33, C34). The histopathology slides were evaluated by a board-certified veterinary pathologist (K. Baiker), and a subset of cases was additionally reviewed by an expert in medical (human) cardiovascular pathology (M. N. Sheppard). Pathologic diagnoses were generated for each case and lesions graded according to severity. IF was used to describe the accumulation of excessive ECM proteins expanding the interstitial space between cardiomyocytes; RF being used when the scar tissue replaced cardiomyocytes. Those cases that were affected by myocardial fibrosis (IF and/or RF) for which no cause could be identified were given a diagnosis of IMF.
Immunophenotyping
Of those hearts that had undergone detailed cardiac examination, samples from 8 zoo-housed chimpanzees (C1, C3, C4, C7, C10, C15, C19, C20) with variable degrees of IMF and 2 African sanctuary chimpanzees (C12, C13) without fibrosis were also subjected to immunohistochemical examination. Tissue sections from the anterior left ventricular wall were immunolabeled for CD3, CD20, IBA-1, and α-SMA (alpha-smooth muscle actin; Supplemental Table S2; Veterinary Laboratory Service, Liverpool, UK) using antihuman antibodies. Chimpanzee splenic tissue was used as a positive control (Supplemental Figs. S1–S3). Within cardiac slides, vascular smooth muscle cells in arterial walls were used as an internal positive control for the α-SMA antibody. The negative control consisted of substitution of the primary antibody with isotype matched murine immunoglobulin (Supplemental Fig. S4).
Detection of Viruses
Fresh frozen heart tissue samples from 6 chimpanzees (C4, C8, C9, C11, C15, C20; Supplemental Table S3) taken at postmortem examination were tested for the presence of cardiotropic viruses at the Primate Biomedical Research Centre (Netherlands; https://www.bprc.nl/en/our-services#diagnostics;AAALACaccreditation). Nucleic acids were extracted and PCR and RT-PCR carried out to amplify the sequences of encephalomyocarditis virus, human adenovirus, parvovirus virus B19, human pan-enteroviruses, human herpesvirus 6, and pan-herpesviruses. Subsequent IHC for herpesvirus antigen (human herpes simplex viruses 1 and 2) and human cytomegalovirus was carried out at an accredited medical commercial diagnostic laboratory (University College London Hospitals, HSL Advanced Diagnostics; http://www.hsl-ad.com/; UKAS to ISO 15189:2012 accreditation standards) on those samples for which a positive PCR result was obtained.
Vitamins D and E, Selenium, and Glycosylated Hemoglobin
Whole blood was collected via femoral venipuncture from 20 chimpanzees as part of the routine preventive health care program at a UK zoological collection, under the Veterinary Surgeons Act 1966. Of these 20 chimpanzees, 11 (cases AC1–AC11) were still alive, and 9 (cases C2–C4, C8, C9, C15, C20, C30, C35) subsequently died and underwent detailed cardiac examination as previously described. Due to the opportunistic nature of sample collection, time between blood sampling and death for these 9 chimpanzees varied from 0 days to 8.5 years. Serum and EDTA-preserved whole blood samples were frozen and shipped, on ice, to a United Kingdom Accreditation Service (UKAS)-accredited laboratory (Laboratory Medicine-Central Manchester University Hospitals) where vitamins D and E, selenium, and glycosylated hemoglobin (HbA1c) were measured. In the absence of established chimpanzee normal values, human reference intervals were used to interpret results: vitamin E = 11.6–34.8 umol/l, selenium = 0.8–1.5 umol/l, 25-OH-vitamin D >50 nmol/l, HbA1c < 48 mmol/mol.
Statistical Analyses
Statistical analyses were carried out using SPSS Statistics (v.24). Data were assessed for normality using a Shapiro-Wilk test. Age at death among European zoo versus African sanctuary chimpanzees and vitamin D levels during high (May to October) versus low (November to April) ultraviolet (UV) light seasons were compared using a Mann-Whitney U-test. Confidence intervals were set at 95%; P < .05 was considered statistically significant.
Results
Cardiac Samples From African Sanctuary Chimpanzees
Multiple acute myocardial infarcts, thromboemboli, and sepsis were evident in the heart section of 1 adult female sanctuary-housed chimpanzee. No evidence of interstitial or replacement fibrosis was identified in any of the remaining 22 chimpanzees for which archived histological sections were examined.
The 2 African sanctuary chimpanzees (C12 and C13; 11 years and 13 years; male) underwent a comprehensive cardiac examination and showed no gross heart lesions and no histologic evidence of IF or RF.
Cardiac Samples From European Zoo Chimpanzees
Macroscopic examinations revealed a mild to moderate left ventricular hypertrophy (concentric thickening of the left ventricular free wall and septum) and atrial dilation in 6 of 33 (18%) adult zoo-housed chimpanzees (Supplemental Table S1). Histological examination revealed mild to moderate degrees of fiber and nuclear hypertrophy within the midmyocardium of the left ventricular free wall and septum of these animals. All affected hearts also showed variable degrees of IF, perivascular fibrosis, and RF affecting multiple or all examined myocardial sections.
One case (C33) was diagnosed with hypertrophic cardiomyopathy (HCM) based on marked, random myofiber disarray. One adult female chimpanzee (C29) had a mild dilation of the left ventricle and atrium and a mildly increased mitral valve circumference with an overall enlarged heart. Histologically in this case, moderate to marked interstitial fibrosis compressed midmyocardial myofibers into thin and elongated, wavy fibers within the left ventricle and septum. Other adjacent areas nearby had fiber and nuclear hypertrophy. Multifocally, the subepicardial and subendocardial spaces contained variably sized areas of RF. The findings were interpreted as dilated cardiomyopathy (DCM). One elderly, female chimpanzee (C27) showed a mild to moderate aortic valve stenosis with a secondary mild left and right ventricular hypertrophy (likely due to chronic pressure overload and pulmonary hypertension, respectively).
Histopathological examination of consistent areas of each heart showed a diffuse, mild variation in myocardial nuclear size in all investigated chimpanzees; interpreted by the authors to be a normal finding in hominids. There were mild to severe fibrotic changes affecting 1 to all 4 chambers in 30 of 33 (91%; age range, 9–65 years) of the European zoo-housed chimpanzees investigated (Supplemental Table S1). IF was the most consistent feature; it varied from minimal to marked (Figs. 1, 2) and was identified in all 30 of the chimpanzees with MF. Perivascular fibrosis (expansion of the vascular adventitial matrix) was also identified in these 30 zoo chimpanzees; its severity closely mirrored that of the IF and therefore was interpreted as being part of the same change and/or process. IF appeared to begin in very mild cases within the midmyocardium of the left ventricular wall and interventricular septum and with increasing severity involved the entire myocardium of all 4 chambers. Minimal to severe, randomly distributed myocardial RF that occasionally extended into the subepicardial space and involved the epicardium was also identified in 29 of 30 animals with MF (Figs. 3, 4). Masson’s trichrome staining of a subset of cases (n = 13; C1, C3, C4, C7, C10, C12, C13, C15, C19, C20, C23, C33, C34) confirmed the presence and extent of the IF, RF, and perivascular fibrosis (Figs. 1, 2).
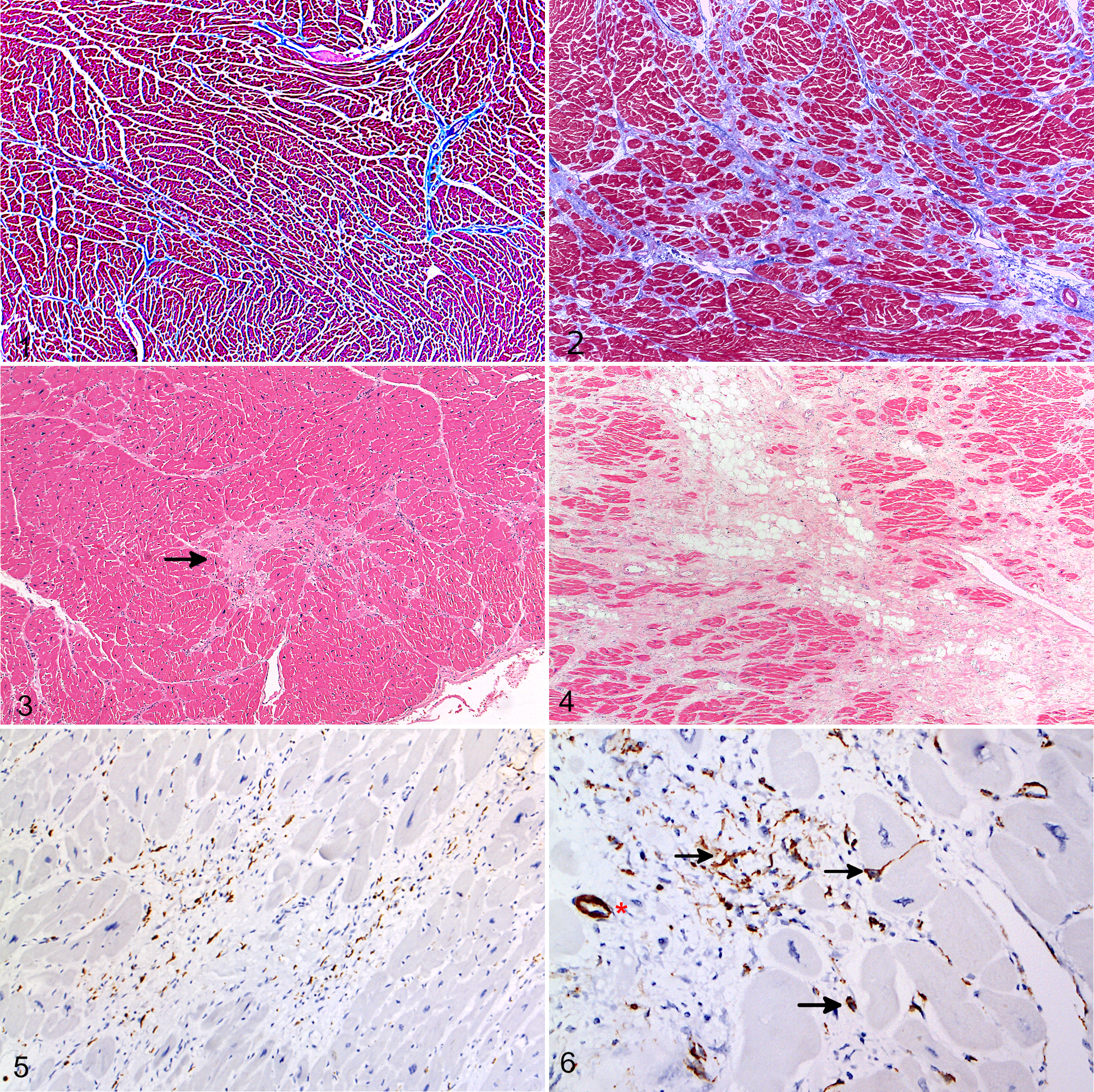
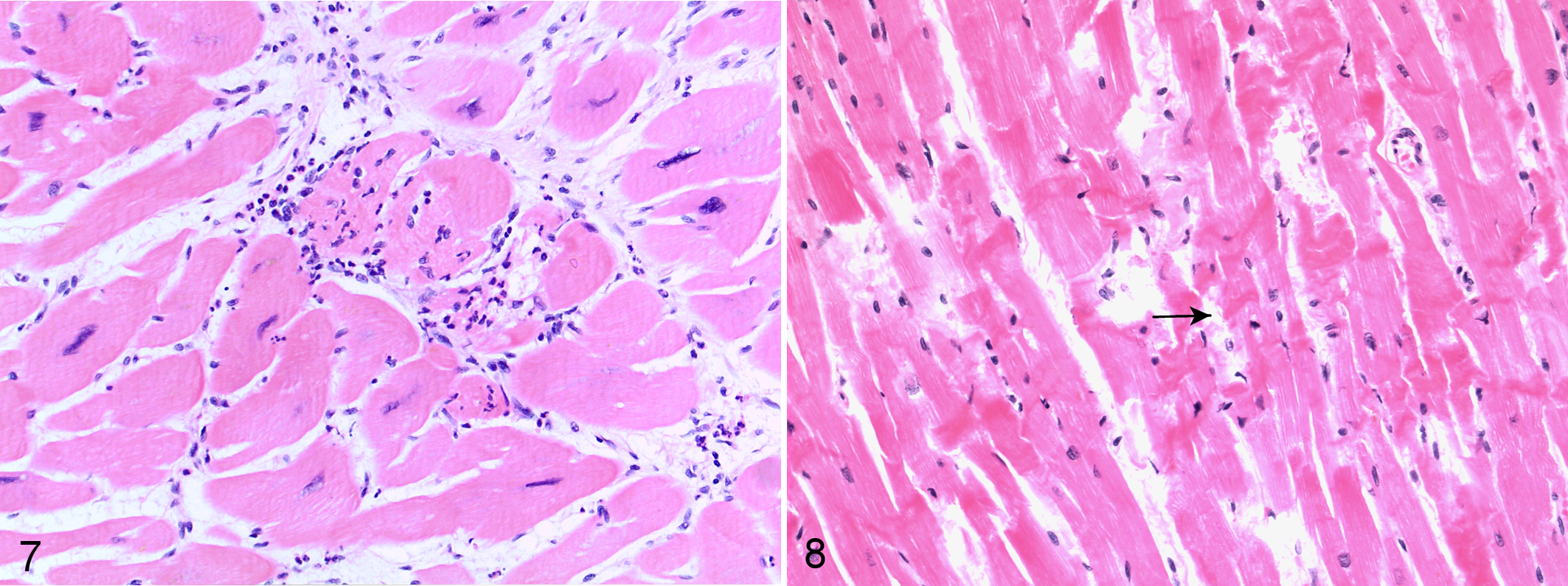
A minimal to mild, occasionally moderate, mixed leukocytic infiltration was also identified in the myocardium of chimpanzees with MF. It was consistently associated with areas of replacement fibrosis and more pronounced in granulation tissue. IHC, carried out on 10 chimpanzees, demonstrated the leukocyte population to be composed mainly of CD3-positive T lymphocytes and IBA-1-positive macrophages (Fig. 5) with fewer neutrophils, eosinophils, and CD20-positive B lymphocytes (Supplemental Figs. S5–S7). Hearts unaffected by MF showed no or low numbers of α-SMA reactive myofibroblasts (Supplemental Fig. S8). Greater numbers of scar-related α-SMA reactive myofibroblasts as well as IBA1-reactive macrophages and scar-related, T-cell dominant, minimal to mild leukocyte populations were present in all MF-affected hearts, with their numbers often seemingly mirroring the severity of myocardial changes (Fig. 6). The extent of eosinophil and neutrophil infiltration was greater in cases of sudden death and in association with foci of acute myocytolysis (Fig. 7). Additional changes in the myocardium of cases of acute cardiac failure included acute hemorrhages and variably sized areas of contraction band necrosis (Fig. 8). Areas of RF that appeared to be older due to a more organized collagen deposition and lack of vascularization and leukocytes often showed increasing numbers of mature adipocytes within the center of the lesion (so-called fibrofatty core; Fig. 4). Other chronic changes identified in 2 chimpanzees were scattered fiber calcifications indicating dystrophic calcification of previous cardiomyofiber degeneration.
Detection of Viruses
Three chimpanzees (C9, C11, C15) of the 6 tested were positive for pan-herpesviral nucleic acid on PCR (Supplemental Table S3). Subsequent IHC testing of cardiac tissue from these animals was negative for herpes simplex virus 1 and 2 and cytomegalovirus. PCR testing was negative for encephalomyocarditis virus, human adenovirus, parvovirus virus B19, human pan-enteroviruses, and human herpesvirus 6.
Vitamins D and E, Selenium, and HbA1c
Antemortem analysis of serum/blood vitamin D, vitamin E, selenium, and HbA1c levels was carried out for 20 zoo-housed chimpanzees, 9 of which had also undergone detailed postmortem cardiac examination (Table 1). The HbA1c levels failed to show any evidence of diabetes mellitus (Table 1). Two chimpanzees were shown to be vitamin E deficient (AC2, AC7), whereas levels exceeded the upper limit of the (human) reference interval in 4 animals. No deficiencies in selenium were identified, and levels exceeded the upper limit of the (human) reference interval in 5 animals. Vitamin D levels were significantly higher in samples collected during the high (n = 19) versus low (n = 12) UV-light season (105.0 ± 33.92 nmol/l [mean ± SD] in May to October vs 54.56 ± 22.47 nmol/l in November to April; P < .0001). Using human reference ranges, vitamin D levels were inadequate (25–50 nmol/l; n = 4) or deficient (<25 nmol/l; n = 2) in 6 of 12 (50%) low UV season samples but inadequate in only 1 of the 19 high UV season samples tested (Table 1). This animal was C4, a male chimpanzee that died suddenly after the subsequent winter and was diagnosed with severe IMF on postmortem examination (Supplemental Table S1).
Severity of Myocardial Fibrosis and Concentration of Serum and Blood Analytes for 20 Chimpanzees Screened for Metabolic Disturbances.a
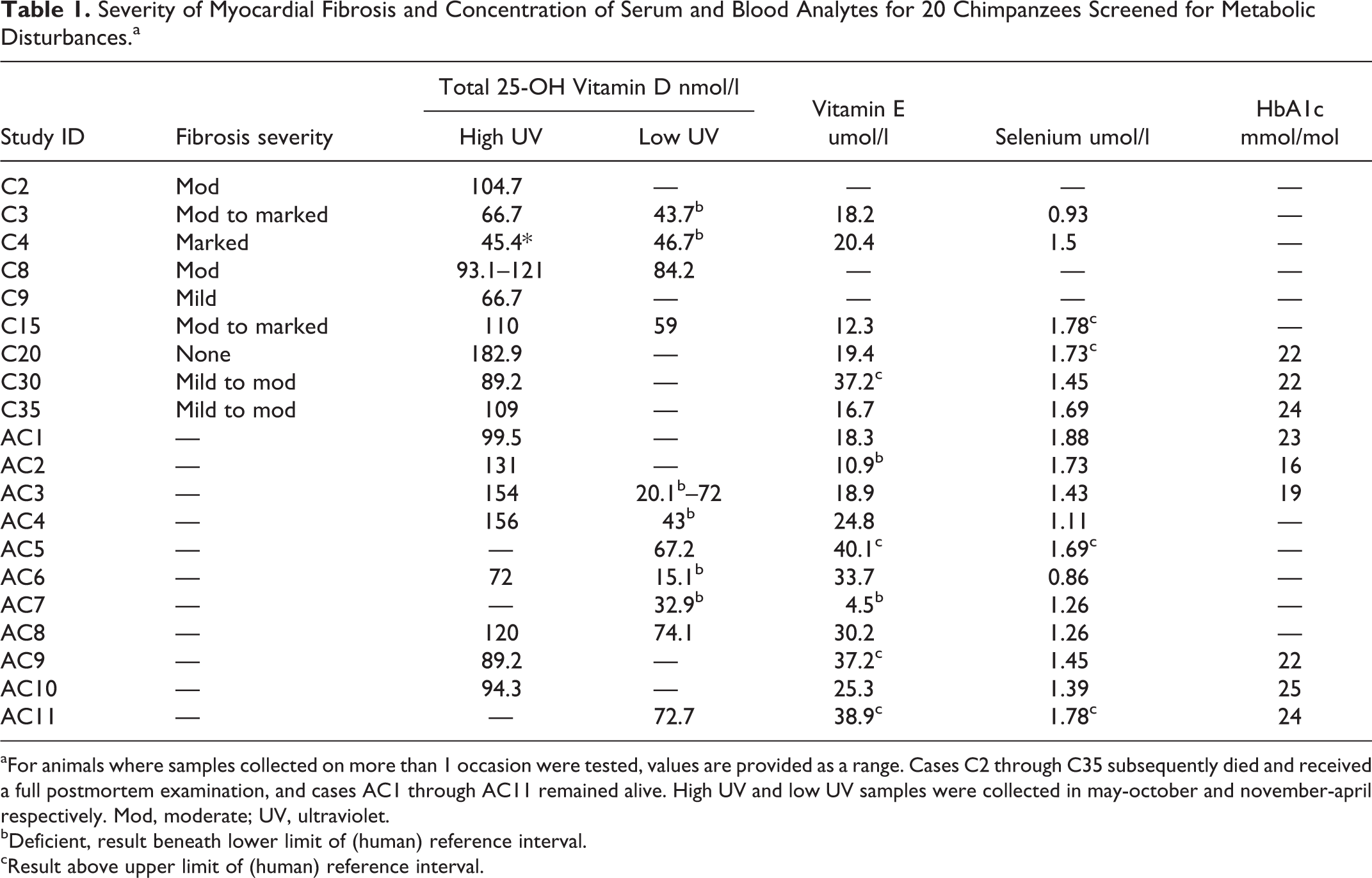
aFor animals where samples collected on more than 1 occasion were tested, values are provided as a range. Cases C2 through C35 subsequently died and received a full postmortem examination, and cases AC1 through AC11 remained alive. High UV and low UV samples were collected in may-october and november-april respectively. Mod, moderate; UV, ultraviolet.
bDeficient, result beneath lower limit of (human) reference interval.
cResult above upper limit of (human) reference interval.
Age at Death
The age at death of the European zoo-housed chimpanzees (n = 33; median = 32 years, [95% CI, 28.02, 37.98 years]) was significantly higher (P < .001) than that of the African Sanctuary (n = 25; median = 12 years, [95% CI, 11.28–14.64 years]) housed chimpanzees
Discussion
Our findings demonstrate that European zoo-housed chimpanzees exhibit a similar spectrum of cardiovascular lesions as those described in humans and other species, including primary cardiomyopathies such as HCM and secondary ventricular hypertrophy. Given that many, if not all, of these conditions are associated with pathological remodeling caused by the excessive deposition of ECM proteins, 52 this suggests a great need for care when interpreting of the presence of MF in the chimpanzee. It also demonstrates the need for a comprehensive cardiac postmortem examination and the histologic evaluation of consistent samples taken from multiple areas of the heart to ensure adequate diagnoses can be made.
A considerable proportion (n = 22/35; 63%) of chimpanzees in this study did, however, die due to or with cardiac fibrosis, which was not accompanied by macroscopic changes and for which the cause, despite extensive whole-heart examination, remains unclear. It has long been known that this pathological entity, referred to here as idiopathic myocardial fibrosis, is a frequent postmortem finding among captive chimpanzees and other great apes. 16,45,46 Inconsistencies in postmortem examination and sampling between zoological collections has, however, previously resulted in a lack of good-quality comparable data available for research on this topic. 16,45,46
This study therefore also aimed to expand on current descriptions of chimpanzee IMF and investigate potential causes. It utilized a consistent, detailed, and published methodology to examine and subsequently characterize the macroscopic, histopathologic, and immunohistochemical features of chimpanzee IMF. The changes associated with IMF in chimpanzees closely resembled a condition of the same name that is associated with sudden cardiac death in humans. 19 In the chimpanzees examined as part of this study, however, the degree of inflammation was mildly more prominent than would typically be observed in humans with the same condition (M. Sheppard, personal communication). This led to speculation that chimpanzee IMF may in fact be the sequela of an acquired, inflammatory, and perhaps infectious condition.
This possibility was investigated by IHC and more detailed characterization of the inflammatory response. The findings, however, were instead suggestive that the observed scar-related, minimal to mild lymphocyte and macrophage population was in fact related to a mild, chronic reaction and did not appear to be the inciting cause of myocardial damage. This is further supported by our inability to demonstrate the involvement of viral infection in IMF in this study. Since chimpanzee blood samples are commonly pan-herpesvirus-positive on PCR, the 3 positive PCR results for pan-herpesvirus in cardiac tissue were interpreted as likely being the result of blood contamination. The possibility that other, less known viruses might be involved in the pathogenesis of IMF, however, cannot be ruled out and warrants further investigation.
Interestingly, granulocytes were often present within the lesions in those chimpanzees that suffered acute, sudden cardiac deaths. These lesions also often had acute cardiomyofiber lysis and/or contraction bands that were not seen in chimpanzees that had been euthanized. Given that such myofiber lysis can occur in the event of terminal dysrhythmic events, these findings therefore support the previously held notion that death in affected chimpanzees might be caused by the pro-arrhythmogenic effect of MF. 11,20,24
Myocardial fibrosis is a nonspecific response to cardiac insult or injury and one of the most common histologic features of the diseased heart. The fact that MF is a feature in primary and secondary cardiomyopathies as well as many other cardiac and noncardiac diseases has previously made definitive interpretation of its presence in the chimpanzee heart difficult. Given that the pathophysiology of the 2 types of fibrosis (replacement and interstitial) differ, 28,39 determining which is predominant in chimpanzee IMF could provide powerful insight into its pathogenesis.
This study demonstrated that both IF and RF are histopathological features in chimpanzee MF and may be part of the same disease process, with RF developing following chronic, progressive, and/or severe IF. Given that IF can be halted or even reversed, this finding has implications for the potential treatment of affected individuals and therefore reduction of mortality.
In humans, one of the most common causes of nonischemic IF is systemic hypertension. 18 Left ventricular hypertrophy was indeed diagnosed in 5 adult chimpanzees as part of this study and can result from systemic hypertension. Review of the clinical history supplied for these animals revealed that these animals had a history of chronic renal disease, suggesting that systemic hypertension might have been the cause of their left ventricular hypertrophy. To which degree these chronic secondary changes caused or contributed to the MF present in these animals is unclear. The chimpanzee diagnosed with DCM (C29) also had histologic lesions of chronic, marked renal disease. This animal’s DCM phenotype was therefore interpreted as being the result of terminal cardiac decompensation following a pressure overload and prior undiagnosed left ventricular hypertrophy. These findings therefore highlight the need for extensive postmortem examination of not just the heart but the whole body to be carried out to rule out all possible primary and secondary causes of MF before denoting it as idiopathic. It also suggests a need for further research into the potential overlap between renal disease and the condition reported as IMF and (as has recently been described by Chilton and colleagues 9 ) cardiorenal syndrome in chimpanzees.
Dysfunctional myocardial remodeling, interstitial fibrosis, and perivascular fibrosis are also features of diabetic cardiomyopathy. 1,40 While our study failed to demonstrate any evidence of inadequate glycemic control in the animals that were tested, it is notable that metabolic disease, diabetes mellitus, and its major risk factor obesity are common in captive chimpanzees. 36,53 Indeed, 1 IMF-affected chimpanzee (C17) that underwent postmortem examination only was declared by the submitting zoo to have a clinical history of diabetes mellitus. Screening for diabetes (resting blood glucose, fructosamine, HbA1c) should still therefore form part of routine health assessment, especially for geriatric animals.
Despite previous reports that cardiovascular disease is more common among male than female chimpanzees, 24,47 our article showed cardiac disease and specifically IMF to affect males and females with similar frequency (n = 13/21 [62%] males vs 9/14 [64%] females). However, moderate-marked IMF was present in 7 of 13 (54%) males but only 2 of 9 (22%) females, and the mean ages differed between the sexes (male = 27 years vs. female = 39 years). Although the number of animals limits this comparison, the results suggest that males might be affected more severely and/or earlier in life.
The progressive accumulation of collagen in the heart is a recognized feature of cardiac aging in humans even in the absence of other risk factors such as hypertension, smoking, and high cholesterol. 8,33 The age difference between the European zoo (median age = 32 years) versus African sanctuary (median age = 12 years) chimpanzees in this study might therefore explain the observed difference in IMF frequency between them. However, IMF was observed in European zoo-housed chimpanzees as young as 10 years of age (C1) and in 2 subadults (C18, C32; 19 years), suggesting that age alone does not explain the apparent discrepancies in disease prevalence between the 2 study populations.
Another key difference between them was the samples examined. All European samples were subject to the comprehensive macroscopic and histopathologic examination protocol, whereas only small, single-site cardiac sections were available for histopathologic evaluation for all but 2 of the African sanctuary chimpanzees, and so early degenerative changes might have been missed.
If the observed difference in the incidence of IMF between the sanctuary- versus zoo-housed chimpanzees is valid, then differences in their living conditions and management could be considered as possible cardiac disease risk factors. The 2 populations differ in their geographical location, housing type (forest vs. indoor enclosures), climatic conditions (including UV light exposure), diet, rearing history, and genetic lineage. Among other things, these factors are likely to influence serum concentrations of vitamin D.
While there is no published research documenting serum vitamin D concentrations for chimpanzees living in their natural environment, it has been shown that captive chimpanzees without regular access to unfiltered sunlight do indeed experience clinically significant deficiencies. 10,54 This finding was also substantiated by the results of our study, with clear seasonal differences and low levels of serum vitamin D during the low UV season of November to April. Clinical investigations in humans suggest an association between vitamin D deficiency and cardiovascular disease risk. 55
Prior antemortem vitamin D data were available for 9 of 35 (26%) chimpanzees undergoing postmortem cardiac examination, and of these, 3 cases appeared to suggest a possible link between vitamin D deficiency and death due to IMF: Case C4 had inadequate vitamin D levels in the winter 2 years before death as well as during his last summer, while case C3 had inadequate levels at the time of death in winter but sufficient levels the previous summer. Case C20 was one of the only adult zoo chimpanzees examined that was not affected by cardiac fibrosis and also had the highest summer levels of vitamin D (182.9 nmol/L) of all chimpanzees examined. No vitamin D levels were available for this animal during the low UV season, so it is not possible to conclude whether a seasonal deficiency existed. While there are a number of possible explanations for this animal’s comparatively higher vitamin D levels, it is interesting that this chimpanzee was also affected with complete alopecia and paler skin pigmentation, and it could be hypothesized, therefore, that this may have facilitated better cutaneous vitamin D synthesis. 25
The limitations in this preliminary data set, not least the variation in time between collection and death, means that these observations are only circumstantial. The findings do, however, highlight this potential association as an important area for further investigation.
This study showed, for the first time, an association between the presence of myofibroblasts and the presence and severity of chimpanzee MF. Upon injury, TGFβ1 stimulates the transformation of human cardiac fibroblasts into profibrogenic myofibroblasts, a process that, in vitro, is inhibited by vitamin D. 37 The increase in myofibroblast numbers producing excessive collagen seen in diseased versus healthy myocardial tissues as demonstrated by IHC in this study suggests that a similar process might take place in the chimpanzee heart. This in turn suggests that the low serum vitamin D concentrations reported might promote the progressive accumulation of collagen, which is seen in chimpanzee IMF, and therefore be associated with an increased risk of disease or accelerated aging in these animals. Additional work would be needed to confirm this hypothesis, but if confirmed, it might suggest that correcting vitamin D deficiency could reduce disease prevalence and progression. Given these findings and the fact that vitamin D is known to play an immunoregulatory role in various diseases, its potential role in cardiac and overall health of captive chimpanzees warrants further investigation.
Conclusion
In conclusion, our study shows that, as in laboratory and American zoo-housed populations, a significant proportion of European zoo-housed chimpanzees also exhibit myocardial fibrosis. In most instances, this fibrosis is not accompanied by any macroscopic lesions. Despite extensive postmortem examination, no obvious cause could be identified, and the pathogenesis is not yet understood. The results of this study identify some important avenues for further exploration of the etiopathogenesis of chimpanzee myocardial fibrosis.
Supplemental Material
Supplemental Material, Combined_supplemental_materials-Strong_et_al_rev - Idiopathic Myocardial Fibrosis in Captive Chimpanzees (Pan troglodytes)
Supplemental Material, Combined_supplemental_materials-Strong_et_al_rev for Idiopathic Myocardial Fibrosis in Captive Chimpanzees (Pan troglodytes) by Victoria Strong, Sophie Moittié, Mary N. Sheppard, Matyas Liptovszky, Kate White, Sharon Redrobe, Malcolm Cobb and Kerstin Baiker in Veterinary Pathology
Footnotes
Acknowledgements
We thank the Pathology team at the School of Veterinary Medicine Science at The University of Nottingham and specifically Alan Lasslett and Mel Hagarty for their excellent technical assistance. Thanks also goes to Lucy Rudd for her hard work and The EAZA Great Ape TAG for their ongoing support. Finally, we are extremely grateful to Tacugama Chimpanzee Sanctuary, Jenny Jaffe, Dr Gerry M. Dorrestein, and all zoological collections, pathologists, and veterinary surgeons who donated samples, data, or their time and expertise for this project.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.