
Abstract
The aim of this study was to assess whether pregnant mice represent a useful model to study the reproductive pathology of Campylobacter jejuni IA3902 using the end point of positive microbial culture of the organism from the fetoplacental unit. Pregnant BALB/c and CD-1 mice (14 days’ gestation) were inoculated orally and intraperitoneally (IP) with 1 × 109 colony-forming units/ml of C. jejuni IA3902. The organism was recovered by microbial culture from the fetoplacental unit in 10 of 10 BALB/c and 10 of 10 CD-1 IP-inoculated pregnant mice and in 29% (2/7) BALB/c and 38% (3/8) CD-1 orally inoculated pregnant mice. Gross reproductive lesions included necrosuppurative placentitis, fetal resorption, intrauterine fetal death, stillborn pups (dead neonates), and multifocal hepatitis. Histological changes consisted of locally extensive neutrophilic and necrotizing placentitis with intralesional bacterial colonies of C. jejuni, ulcerative endometritis, random multifocal hepatitis, and rare cholecystitis. Immunohistochemistry for the major outer membrane protein of C. jejuni revealed moderate to large numbers of the organism at the periphery of the placental discs, within trophoblasts and extracellularly, with invasion into the placental disc largely via the vascular network. The organism is trophic for neutral mucin, iron, and L-fucose within the murine placenta. C. jejuni IA3902 has affinity for the murine reproductive tract, specifically the fetoplacental unit, where it results in a necrotizing placentitis with positive microbial recovery after both IP and oral challenge in BALB/c and CD-1 pregnant mice.
Keywords
Campylobacter jejuni is a small (0.2–0.9 μm wide and 0.2–5.0 μm long), spiral, Gram-negative bacterium, principally known as a zoonotic pathogen. 12 The organism is nonspore forming but vigorously motile by means of a single polar flagellum at one or both ends. 21 Campylobacter is a leading cause of foodborne bacterial disease in humans and a leading cause of reproductive loss in sheep, with abortion rates that range from 5% to 50% in infected flocks. 17 C. jejuni colonizes the gastrointestinal tract of a wide range of animals that act as reservoirs of infection by fecal shedding. Insights into the molecular mechanisms of disease and immunopathogenesis have been severely limited due to the absence of convenient murine infection models. Historically, murine models of C. jejuni infection have been limited by sporadic colonization and/or absence of clinical disease. 4 The reasons for C. jejuni resistance in mice are likely influenced by continuous active competition from resident intestinal microbiota that outcompete C. jejuni for survival and tolerance of the murine immune system to C. jejuni. 19 A murine model for the study of C. jejuni abortion is needed to reduce research costs compared to the commonly used guinea pig model. There would also be an increased likelihood of reproducibility of experimental results compared to other animal models, including the piglet, weanling ferret, gnotobiotic canine pup, and primate models. 4 In addition, the reproductive system of the laboratory mouse has been extensively studied. Mice have a hemochorial placenta similar to humans, a short gestation period, and defined genetics, and their immune system has been well characterized and is similar to that of humans. 10 The mouse model has proven useful for the study of the pathogenesis of reproductive disease caused by Chlamydophila abortus and in several vaccine-related investigations. 9 A pregnant mouse model has been characterized for the study of bovine Tritrichomonas foetus infection, 1 and murine models have been established for human reproductive research on preeclampsia. 2
Several studies have demonstrated that abortion can be reliably produced in guinea pigs following either oral or intraperitoneal inoculation of C. jejuni. 7 Despite routine recovery of C. jejuni from naturally aborted ovine fetuses, there has been limited success in reproducing abortion in experimentally infected pregnant sheep, the species in which reproductive loss is of greatest concern. There have been no published reports of the reproductive pathology of C. jejuni in mice, which, because of their size and ease of handling, are the most commonly used laboratory research animal model. The mouse model has been used by some researchers to study other aspects of campylobacteriosis, and it has been demonstrated that bacteremia is a very common and consistent finding, which is an essential step in the pathogenesis of abortion. 22 Mice have also been used in several Campylobacter vaccination studies. The purpose of this study was to investigate whether mice represent a useful model to assess the reproductive pathogenicity of C. jejuni.
Materials and Methods
Experimental Design
The reproductive pathology of C. jejuni IA3902, a clinical isolate of clone SA, was assessed after either oral or intraperitoneal inoculation in 2 strains of mice. Sixty 7- to 8-week-old mice, including 30 outbred CD-1 (24 female and 6 male) and 30 inbred BALB/c mice (24 female and 6 male), were purchased from Charles River Laboratories. The BALB/c mouse strain was selected as it is frequently used in immunological research and reproductive studies due to ease of breeding. The CD-1 stock was selected to compare with an inbred strain (BALB/c) while being the outbred stock of mice most commonly used in research. All mice were acclimated for 5 days before use. Four female mice were group housed with a male of the same species for 14 days for breeding. On day 15, all study animals were bled and inoculated with abortifacient C. jejuni IA3902. Then, 0.1 to 0.2 ml of blood was collected from the lateral saphenous vein using a 25-gauge (G) needle.
Ten female mice of each strain were inoculated orally after 12 hours of fasting. Mice were lightly anesthetized via gaseous anesthesia in an anesthetic chamber with 2% isoflurane and then quickly removed and inoculated orally with 0.5 ml C. jejuni IA3902 at 109 colony-forming units (CFU)/ml, via gastric lavage using a stainless steel, ball-tipped 24-G 1.0-inch animal feeding tube. Ten female mice of each strain were inoculated intraperitoneally (IP). Each mouse was briefly placed on its back with its head tilted slightly downward and 0.1 ml C. jejuni IA3902 at 109 CFU/ml injected IP using a 25-G needle. After inoculation, female mice were immediately housed in individually identifiable cages for observation twice daily for 1 week. For each group, there were 4 control (noninoculated) female mice per strain also housed individually. Over the subsequent week, all mice were monitored twice daily for clinical signs of inactivity, unresponsiveness, ruffled hair coat, diarrhea, vaginal discharge, and abortion (fetal loss). All mice were humanely euthanized 1 week postinoculation using ketamine/xylazine followed by exsanguination. At the time of necropsy, the following samples were collected: heart blood for serum collection and bacterial culture, cecal swab, uterus and uterine contents (placenta and fetus) for bacteriology and maternal liver with gallbladder, uterus, and uterine contents for histopathology. The study was approved by the Iowa State University Institutional Animal Care and Use Committee (IACUC), protocol 5-15-8018-M, and fully compliant with the regulations of the Laboratory Animal Resources (LAR) group at Iowa State University. Appropriate personal protective equipment was worn when mice were inoculated, samples collected, and necropsy performed both in the animal treatment and in the necropsy rooms.
Necropsy, Microbial Culture, and Histopathology
All mice were subjected to a complete necropsy. At necropsy, all gross lesions were noted and samples collected for microbial culture and histological examination. Cecal swabs were obtained using sterile minitip cotton swabs with transport media immediately after euthanasia. Uterus, placenta, and fetal tissue were harvested in sterile petri dishes for Campylobacter culture. All samples for microbial culture were refrigerated immediately upon collection and plated on the day of collection. Cecal swabs and blood were directly streaked onto culture media. Uterus and fetoplacental tissue were minced, swabbed, and streaked onto MH agar containing a Campylobacter-selective supplement (polymyxin B, rifampicin, trimethoprim, and cycloheximide) and a Campylobacter growth supplement (sodium pyruvate, sodium metabisulfite, and ferrous sulfate). Plates were incubated for 48 hours in anaerobic jars under microaerobic conditions at 42°C. Campylobacter colonies were counted on each plate to determine the number of CFU in each sample.
Uterus, fetoplacental unit, yolk sac, liver, and gallbladder were examined histologically. Tissues were fixed in 10% neutral buffered formalin for 24 hours. Preserved tissues were paraffin embedded and 5- to 7-micron diameter sections were mounted on slides and routinely stained with hematoxylin and eosin.
Neutrophil Scoring of Reproductive Lesions
The mean number of neutrophils present in 5 fields (40× magnification) was assessed for each of 4 locations in the reproductive tissue, including the periphery and parenchyma of the placental disc, the uterine wall, and uterine lumen. Neutrophil counts were scored for each location using the following criteria (1 = <10 neutrophils/high-power field [hpf], 2 = 11–25 neutrophils/hpf, 3 = 26–50 neutrophils/hpf, and 4 = >51 neutrophils/hpf).
Lectin Histochemistry
Three-micron-thick serial sections were set on aminoalkylsilane-coated glass slides, baked at 56°C for 2 hours, and routinely deparaffinized in xylene and rehydrated in graded alcohol solutions and water baths. Endogenous peroxidase inhibition was achieved by immersion (2 immersions; 10 minutes/immersion) in baths of 3% hydrogen peroxide (H2O2) in water. Antigen was unmasked by treating sections with Tris-EDTA (pH 9.0) in a stream bath for 20 minutes, cooled to room temperature, and rinsed 3 times in phosphate-buffered saline (PBS) prior to placement in an automated cell staining system (BioGenex). The lectin used was the commercially available biotinylated Lotus tetragonolobus lectin agglutinin I (LTA; Vector) applied to sections at 20 mg/ml and incubated at 22°C for 30 minutes, followed by rinsing in a bath of PBS solution for 5 minutes. Lectin binding was visualized using a commercial kit (Vectastain Elite ABC; Vector) and chromogen (NovaRED; Vector) as per the manufacturer’s instructions; the sections were then counterstained with hematoxylin and mounted routinely. Specificity of lectin binding was confirmed by preabsorbing lectins in 250 mM L-fucose prior to application to serial sections. Negative controls were prepared from serial sections with the lectin omitted and replaced with dilution buffer. Lectin binding was subjectively quantified in the fetoplacental unit, uterus, and yolk sac (visceral and parietal layers) as low/absent, moderate, or abundant in random 40× magnification fields and classified as to whether staining was cytoplasmic or membranous.
Special Stains
Perls’s iron stain, Alcian blue pH 2.5, and the periodic acid–Schiff (PAS) reaction without diastase pretreatment were performed on 12 sections randomly and equally drawn from each of the groups along with 1 control mouse slide. Serial sections were cut to 5 μm, stained, and assessed to determine the presence or absence of material with characteristics consistent with iron, acid mucin, and neutral mucin within the fetoplacental unit, yolk sac (visceral and parietal layers), and uterine wall. Staining was subjectively classified as low/absent, moderate or abundant, and intracellular, extracellular, multifocal, or diffuse.
Immunohistochemistry
Placental discs collected at necropsy were processed, embedded, and sectioned at 3 microns; mounted on aminoalkylsilane-coated glass slides; and baked for 2 hours at 56°C. Slides were deparaffinized routinely in xylene and rehydrated in graded alcohol solutions and water. Inhibition of endogenous peroxidase was via 2 immersions for 10 minutes each in 3% H2O2 in water. Slides were incubated for 15 minutes at 37°C in 0.1% protease in Tris buffer of pH 7.6, then rinsed 3 times in PBS solution and stained automatically. Nonspecific binding was inhibited by incubation with 10% neutral goat serum at 22°C for 20 minutes. The primary antibody was directed against the major outer membrane protein of C. jejuni, as described by Zhang et al, 23 at a dilution of 1:300 and incubated at 22°C for 60 minutes and then rinsed in PBS for 5 minutes. A commercially available biotinylated secondary antibody was used at a dilution of 1:80 and incubated at 22°C for 15 minutes and then rinsed in PBS for 5 minutes. Slides were then incubated with horseradish peroxidase–streptavidin conjugate at 22°C for 15 minutes and rinsed in PBS for 5 minutes. The final reaction used a commercial chromogen. Slides were rinsed and routinely counterstained with Shandon Harris hematoxylin and Scott’s tap water. Slides were dehydrated through graded alcohol and xylene solutions prior to mounting. Positive controls were sourced from paraffin blocks of ovine placenta that was positive for C. jejuni IA3902 by microbial culture. Negative controls were from uninoculated mice in the study.
Statistical Analysis
A commercial statistical software package, SAS 9.4 version (SAS Institute, Cary, NC), was used to perform a nonparametric Kruskal-Wallis test to detect differences in mean neutrophil scores among inoculation routes (BALB/c oral, BALB/c IP, CD-1 oral, CD-1 IP, and control) and for different locations (placenta periphery and parenchyma, uterine lumen and wall) in mice with C. jejuni isolated from placenta and uterus by microbial culture. Second, the same nonparametric Kruskal-Wallis test was performed to test if different placental culture levels affected neutrophil scores at the placental periphery and in the placental parenchyma, as well as if different uterine culture levels affected neutrophil scores for the uterine lumen and uterine wall. Results were considered significant at values of P ≤ .05.
Results
Gross Pathology
C. jejuni IA3902 was recovered in high numbers (TNTC) from 14 mice. For mice with infected fetoplacental discs, there did not appear to be any obvious effect on litter size or crown-rump length compared to control mice. However, there was a significant decrease in the birth of live young: 93% of mice with infected fetoplacental discs failed to produce live young (Table 1).
Outcomes Following Infection of Pregnant BALB/c and CD-1 Mice With Campylobacter jejuni IA3902.
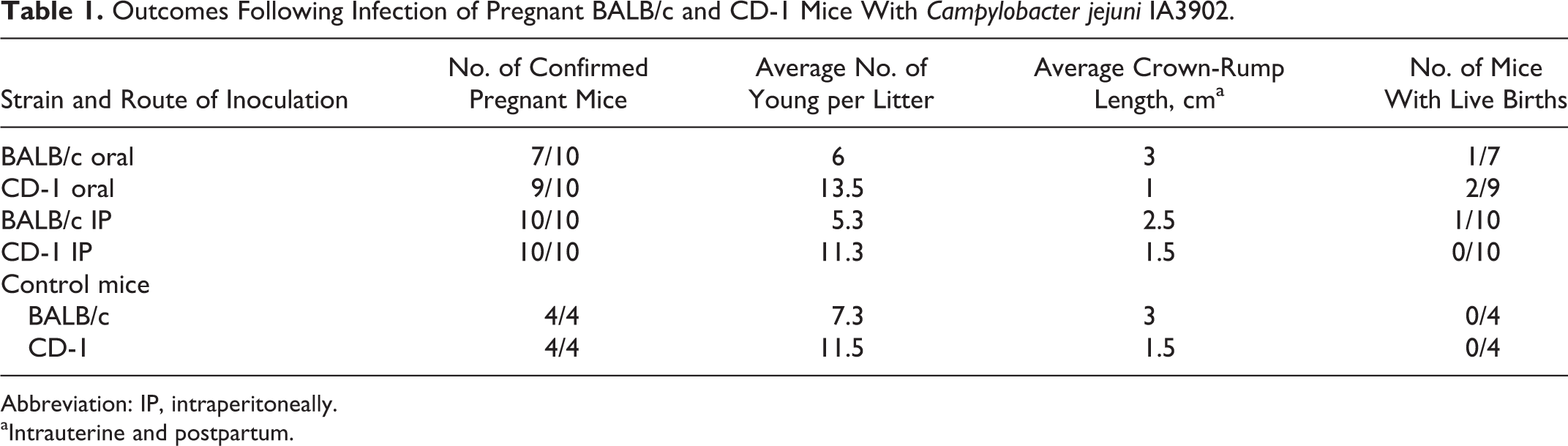
Abbreviation: IP, intraperitoneally.
aIntrauterine and postpartum.
Gross reproductive lesions (Table 2) ranged from the birth of dead pups/neonates (dead newborn mice found in cage), intrauterine fetal death (tan/white midsized conceptus), placentitis (edematous and hemorrhagic, yellow or green placental discs) (Fig. 1), and fetal resorption (placental disc without macroscopic evidence of a subjacent fetus) (Fig. 2).
Gross Reproductive Lesions Following Infection With Campylobacter jejuni IA3902 in Pregnant BALB/c and CD-1 Mice.a
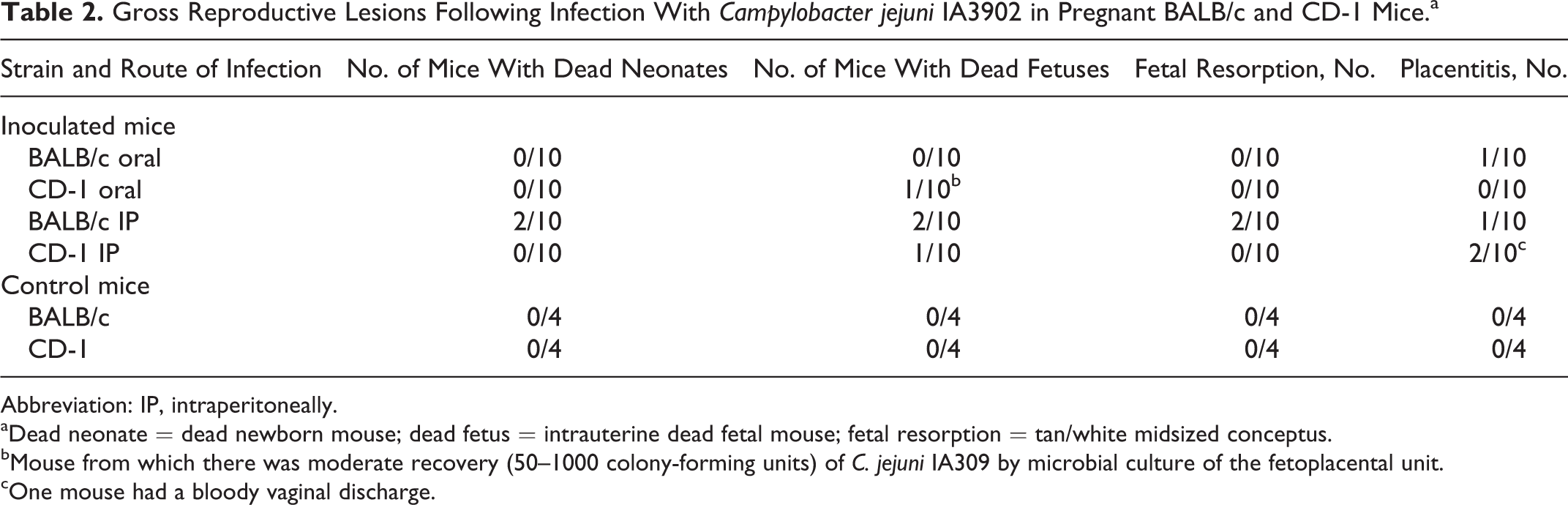
Abbreviation: IP, intraperitoneally.
aDead neonate = dead newborn mouse; dead fetus = intrauterine dead fetal mouse; fetal resorption = tan/white midsized conceptus.
bMouse from which there was moderate recovery (50–1000 colony-forming units) of C. jejuni IA309 by microbial culture of the fetoplacental unit.
cOne mouse had a bloody vaginal discharge.
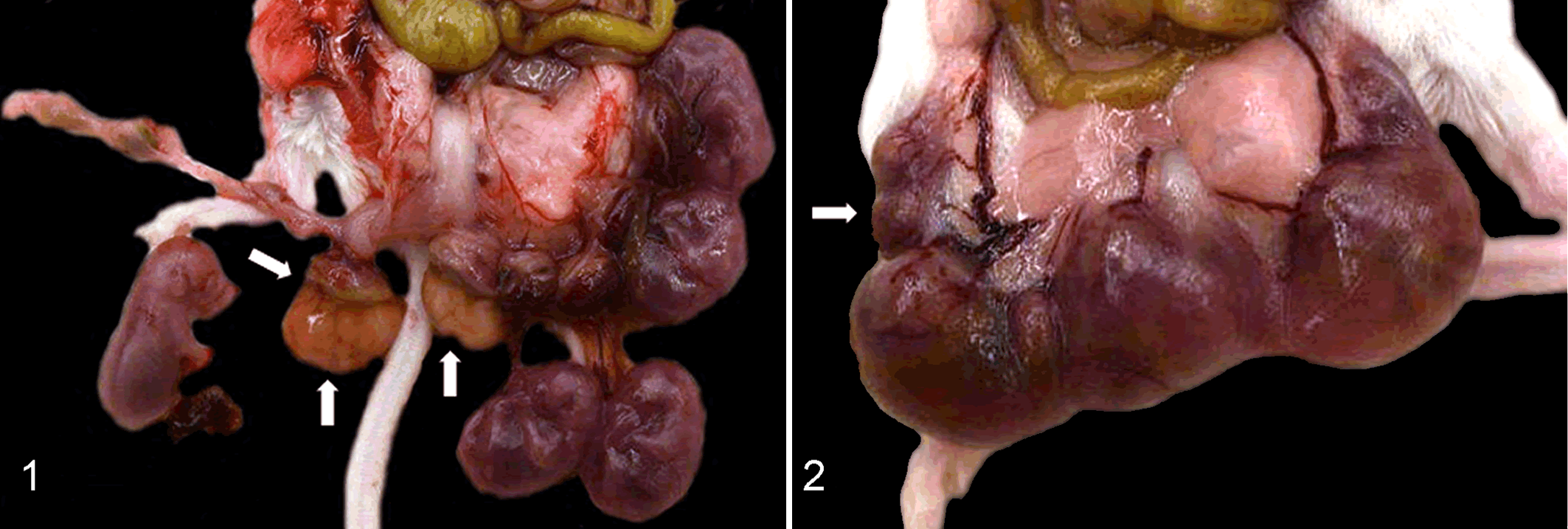
Uterus, mice infected with Campylobacter jejuni IA3902.
At necropsy, gross evidence of infection was found only in animals from which C. jejuni IA3902 was isolated in moderate and high counts, with 100% correlation between the presence of gross reproductive lesions at necropsy and the microbial recovery of moderate to high counts of C. jejuni IA3902. When there was high recovery (TNTC) of C. jejuni from the fetoplacental discs in BALB/c mice inoculated IP, there was a spectrum of lesions, including dead neonates and fetuses, fetal resorption, and placentitis. Both BALB/c and CD-1 mice had placentitis after intraperitoneal inoculation, and the BALB/c strain had placentitis after oral inoculation, with high recovery of the organism from infected fetoplacental tissue. There was intrauterine fetal death in 1 CD-1 mouse with moderate recovery (50–1000 CFU) of C. jejuni from the fetoplacental disc. There were no dead neonates, dead fetuses, or gross or histological evidence of fetal resorption or placentitis in any control mice.
There were rare, 1-mm-wide white foci on the liver of 2 BALB/c mice, 1 of which was inoculated orally and the other IP. C. jejuni was positively identified by culture in very high numbers (>1000 CFU) in both cases.
Microbial Culture
C. jejuni IA3902 was recovered from the cecum from all mice except 2 CD-1 mice (1 was orally inoculated and the other was inoculated IP). The organism was cultured more often and in much higher numbers from the placenta than from the free uterine wall, irrespective of strain of mouse or route of administration. C. jejuni IA3902 had an affinity for the murine fetoplacental unit as it could be cultured from the fetoplacental unit of all of the BALB/c and CD-1 mice that had been inoculated intraperitoneally. Heavy growth (>1000 CFU or TNTC) was detected in CD-1 mice twice as often as in BALB/c mice. After oral inoculation in both mouse strains, C jejuni IA3902 was recovered from the fetoplacental unit of 2 of 7 BALB/c mice (1 at <50 CFU and 1 at >1000 CFU) and 3 of 8 CD-1 mice (1 at 50–1000 CFU at 2 at >1000 CFU) (Table 3).
Microbial Recovery by Culture Following Infection With Campylobacter jejuni IA3902 in Pregnant BALB/c and CD-1 Mice.
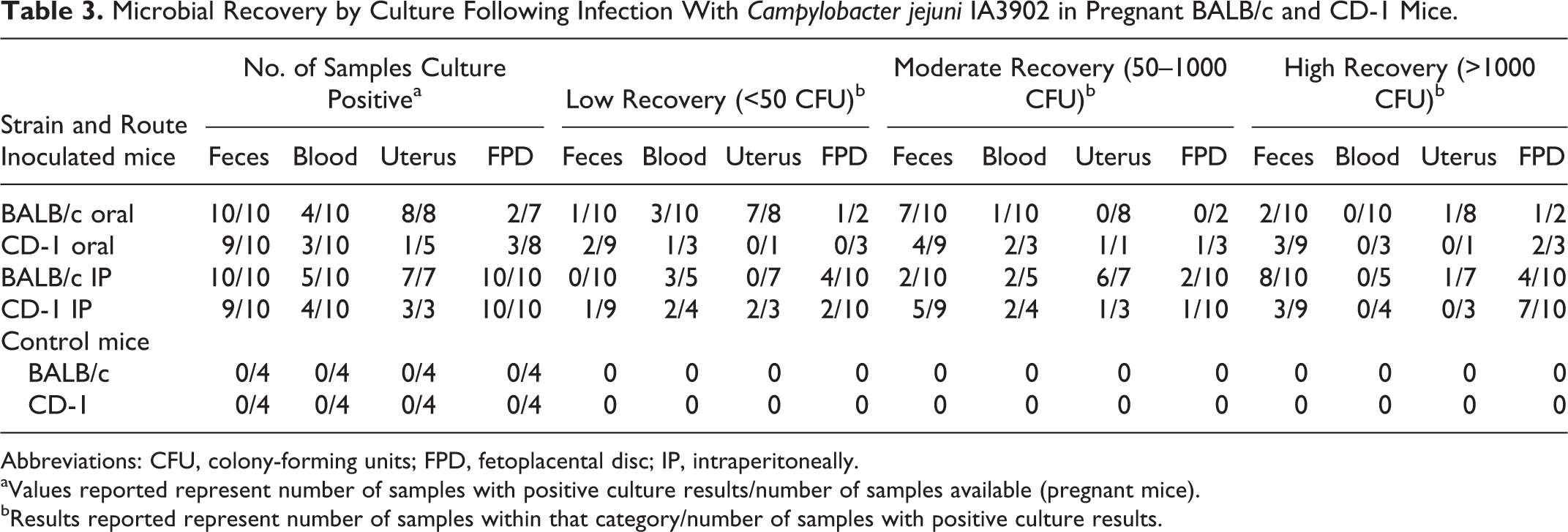
Abbreviations: CFU, colony-forming units; FPD, fetoplacental disc; IP, intraperitoneally.
aValues reported represent number of samples with positive culture results/number of samples available (pregnant mice).
bResults reported represent number of samples within that category/number of samples with positive culture results.
Histopathology
Histologically, the uterine lesions of C. jejuni IA3902 inoculated pregnant mice consisted of multifocal areas of neutrophilic infiltration, endometritis, endometrial ulceration, necrosis, and occasional hemorrhage. There was a neutrophilic placentitis with moderate to large numbers of neutrophils disrupting cellular integrity primarily along the periphery of the placental disc. Within these foci at the periphery of the infected placental discs, there were variable amounts of necrotic cellular debris and hemorrhage with occasional bacterial colonies 150 to 250 microns in diameter. C. jejuni IA3902 was isolated from the fetoplacental unit of all 20 pregnant intraperitoneally inoculated mice (10 BALB/c and 10 CD-1) and from 2 of 7 BALB/c and 3 of 8 CD-1 mice orally inoculated with the organism. Of 14 mice that had high recovery (TNTC) of C. jejuni IA3902 by microbial culture from the fetoplacental discs, 13 also had increased numbers of neutrophils within the periphery of the fetoplacental disc (neutrophilic placentitis) (Table 4). One mouse from the BALB/c IP group did not have a fetoplacental disc available for histological assessment but had high recovery (TNTC) of C. jejuni from the uterus. There were variable numbers of neutrophils within the uterine lumen and endometrium of nonpregnant mice with rare ulcerative endometritis.
Neutrophil Scores in Fetoplacental Discs With High Recovery (TNTC) of Campylobacter jejuni IA3902 Following Oral and Intraperitoneal Inoculation in Pregnant BALB/c and CD-1 Mice.
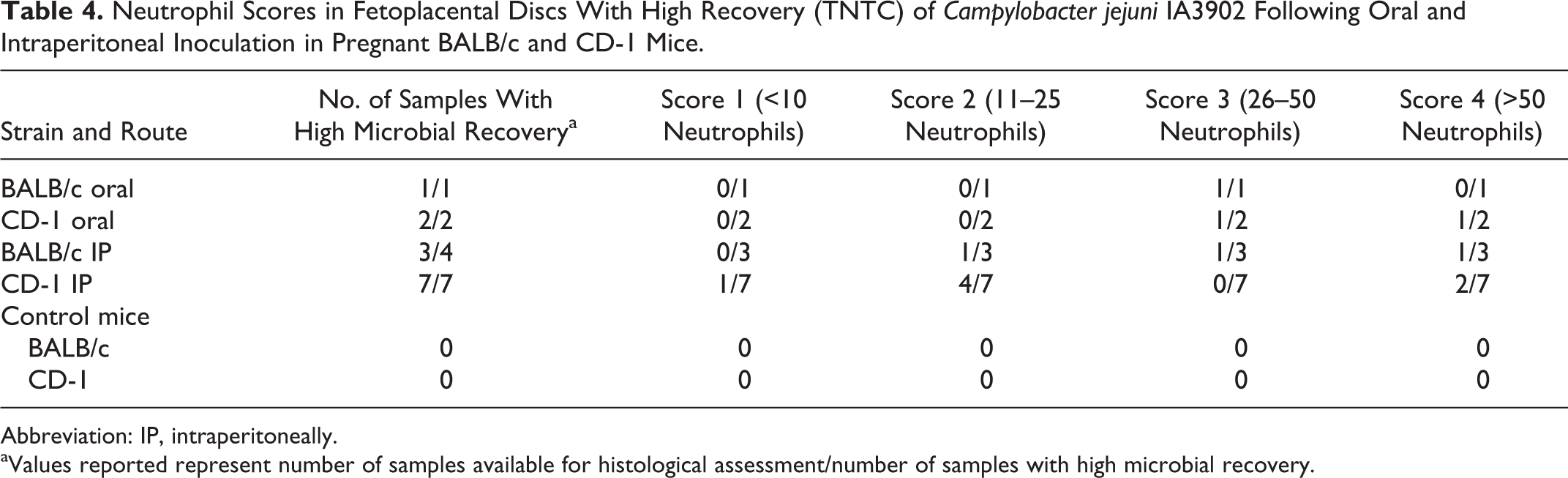
Abbreviation: IP, intraperitoneally.
aValues reported represent number of samples available for histological assessment/number of samples with high microbial recovery.
Inoculated mice that gave birth prior to the study termination had moderate to large numbers of neutrophils present within the submucosa and endometrial glands and had erythrocytes within the uterine lumen. Postpartum control mice had low numbers of neutrophils and erythrocytes within the uterine lumen and submucosa.
There was a significant difference in neutrophil scores among inoculated routes for lumen (P = .0159) and wall (P = .0057), and there was a significant difference among placental microbial culture levels for neutrophil scores for the placental periphery (P = .0049), uterine lumen (P = .0028), and uterine wall (P = .0089).
The spatial distribution of mucin, iron, and L-fucose within the murine placental unit and adjacent tissue was assessed in control and infected mice (Table 5). In sections stained with PAS, which stains neutral mucins, there was moderate to abundant globular staining in the placental disc and yolk sac within the cytoplasm of trophoblasts and yolk sac cells (intracellularly) and extracellularly (Fig. 3). Alcian blue staining, which stains acid mucins, was absent to low. Iron was present multifocally within the parenchyma of the placental disc, mainly within vascular sinusoids (Fig. 4). Lectin histochemistry for LTA demonstrated moderate to abundant, multifocal, cytoplasmic, and membrane staining of cells of the yolk sac and uterine mucosa and extracellular staining at the periphery of the placental disc (Fig. 5).
Histochemistry Findings from Murine Reproductive Tissue Following Infection With Campylobacter jejuni IA3902.a

Abbreviation: LTA, Lotus tetragonolobus lectin agglutinin.
aValues reported represent the average of all evaluated placental material. Scores represent the average of all evaluated tissue from 12 mice: –, low/absent; +, moderate; ++, abundant.
bMultifocal staining.
cMembrane staining.
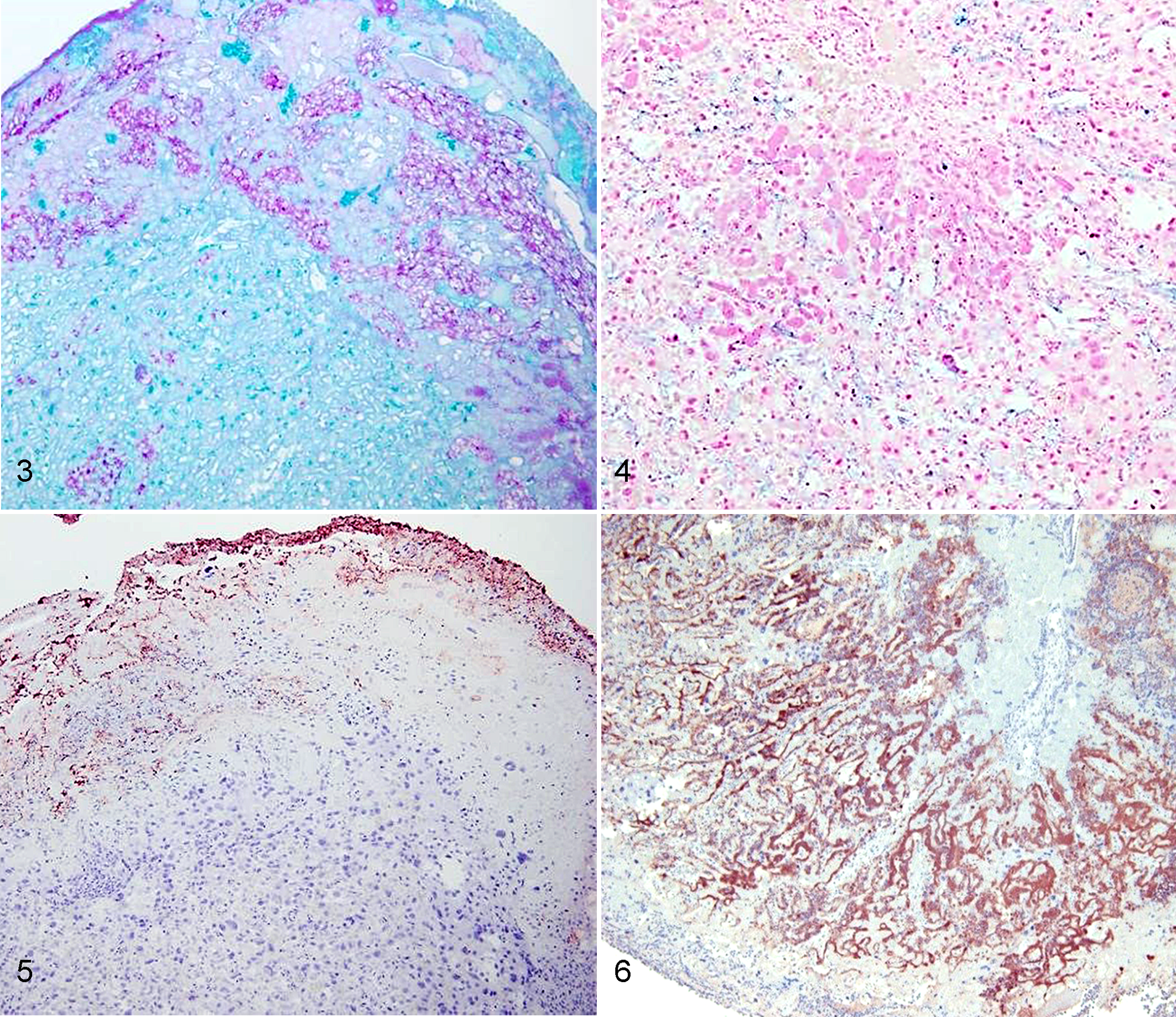
Placental discs, Mus musculus.
Histologically, 3 murine livers had random multifocal necrotizing hepatitis and mild lymphoplasmacytic cholangitis. Rarely, there were increased numbers of mixed inflammatory cells within sinusoids and in the submucosa and the mucosa of the gallbladder (cholecystitis).
Immunohistochemistry
Immunohistochemistry for major outer membrane protein of C. jejuni was performed on 1 placental sample randomly drawn from each of the 4 groups and 1 control. Staining for C. jejuni was completely absent in the control slide but was positively identified within each of the 5 murine fetoplacental discs from which high numbers of C. jejuni were isolated. There was positive staining within the cytoplasm of trophoblasts mainly at the periphery of the placental disc, within neutrophils, and extracellularly in vascular sinusoids of severely affected placental discs (Fig. 6).
Discussion
Several animal models for intestinal campylobacteriosis have been investigated, including an oral ferret, piglet, rabbit, chick/avian, and SCID mouse model, 14 while the pregnant guinea pig has been the primary model for reproductive studies. Despite over 30 years of research, there is no consensus on a widely acceptable animal model for human intestinal and reproductive campylobacteriosis research. Mice are the most commonly used animal to study genital chlamydial infections with 2 well-established murine models in which the course of disease and outcome can be varied depending on the mouse strain, inoculating dose, age of the mice, and hormone levels present. 15
There are several reasons why establishing a murine model for reproductive studies with C. jejuni IA3902 would be useful. The murine and human placenta are both discoid and hemochorial, which supports a similar pathogenesis in reproductive disease studies. Murine models have already been defined for a number of infectious agents and vaccines, and bacteremia has already been demonstrated to be a common and consistent finding with campylobacteriosis. 22 Murine studies would also be ideal for elucidating the genetic factors involved in host defense against C. jejuni infection as there are murine models with defined deletions for components of innate and adaptive immunity. 13 In addition, other experiments in pregnant mice have demonstrated that Campylobacter rectus, an exclusively oral bacterium, can induce adverse effects on the fetus and placenta. 3
In this study, there was a significant decrease in the birth of live young from mice with C. jejuni–infected placentas as 93% of mice with heavily infected fetoplacental discs failed to produce live young. There was also a high correlation between the presence of gross reproductive lesions at necropsy and microbial recovery of moderate to high counts of C. jejuni IA3902. This is similar to what has been demonstrated in guinea pigs. Abortions, while not observed in mice, occurred in 6 of 12 pregnant guinea pigs within 4 and 10 days after oral inoculation with C. jejuni IA39027 and in 1 of 2 pregnant sheep 15 days after oral inoculation with the same agent. 17 The short gestation in mice (18–22 days, compared with 63–70 days in guinea pigs and 147–152 in sheep), dose and time of inoculation, and the relatively short interval between challenge and necropsy may have precluded abortions from being observed in this study. Stillbirths have been reported as a result of C. jejuni IA3902 infection in sheep. 17 Stillbirth occurred in infected mice in this study but has not been reported in experimentally infected guinea pigs. Fetal resorption has been reported in pregnant mice after administration of lipopolysaccharide (LPS) 16,18 and was seen in this study. In 2 BALB/c mice, there was random multifocal hepatitis grossly and histologically with microbial recovery of the organism from hepatic tissue, in addition to cholecystitis. Multifocal hepatitis has also been reported with pregnant guinea pigs inoculated with C. jejuni IA39027 and in both species is likely associated with the bacteremia that precedes fetoplacental infection.
After inoculation with C. jejuni IA3902, there was histological evidence of neutrophilic inflammation within the uterine wall in both strains of mouse, irrespective of pregnancy status or route of inoculation. Endometrial lesions consisted of multifocal areas neutrophilic infiltration, endometrial ulceration, necrosis, and occasional hemorrhage. Histologically, inflammation of the placenta and endometrium also occurs in sheep 11 and guinea pigs. 7 There were variable numbers of neutrophils (scores between 2 and 4) within the endometrium of nonpregnant mice with rare ulcerative endometritis, although the organism could not be recovered by culture from this location in these animals 1 week postinoculation, at necropsy. In comparison, in nonpregnant guinea pigs, the organism was cultured at low levels (<50 CFU) from the uterus in 2 of 11 (18%) animals 2 days postinoculation. 6 It is notable that in the guinea pig study, all animals were challenged IP, the study was terminated at a much shorter interval of 48 hours postchallenge, and low bacterial counts were detected in 2 guinea pigs. Based on histologic changes, it appears that in mice, as in guinea pigs, C. jejuni is trophic for the uterus even in nonpregnant animals. There were no nonpregnant control mice at the end of this study.
Within the fetoplacental unit, there was a neutrophilic placentitis with moderate to large numbers of neutrophils disrupting cellular integrity primarily along the periphery of the placental disc, with necrotic cellular debris, hemorrhage, and bacterial colonies 150 to 250 microns in diameter. Neutrophilic placentitis, a feature of the reproductive tract infection in pregnant guinea pigs, 7 was also a consistent feature in infected fetoplacental tissue irrespective of mouse strain or route of infection. High neutrophil scores correlated with microbial recovery from heavily infected fetoplacental discs, as 12 of 13 mice (92%) from which C. jejuni IA3902 was recovered in high counts (TNTC) had neutrophils scores 2 or higher.
The proposed pathogenesis for Campylobacter abortion is oral ingestion followed by intestinal colonization and translocation, bacteremia, and infection of the fetoplacental unit. In both strains of mice, C. jejuni IA3902 could be isolated from the blood, uterus, and fetoplacental unit after oral inoculation, which supports the suitability of the model as it demonstrates replication of the natural disease with bacteremia resulting from intestinal invasion with subsequent infection of the fetoplacental tissue. However, in both strains of mice evaluated, the percentage of animals with demonstrable fetoplacental infection (29% for BALB/c and 38% for CD-1) is considerably lower than is typically observed in guinea pigs following oral inoculation (70%). 7 Although less desirable for the study of the overall pathogenesis of Campylobacter-induced reproductive disease, IP inoculation resulted in more consistent fetoplacental infection with accompanying gross and histologic lesions. Similar to findings in guinea pigs after IP inoculation, there was high correlation with fetoplacental infection and gross and histological lesions in C. jejuni IA3902–infected murine reproductive tissue.
The organism was positively identified via immunohistochemistry for the major outer membrane protein within the cytoplasm of trophoblasts and infected neutrophils located in necrotic foci at the periphery of the fetoplacental unit, in the uterine lumen and the endometrial submucosal tissue including the vasculature, and within giant trophoblasts at the periphery of infected fetoplacental units. Extracellularly, C. jejuni was located in and around necrotic foci at the periphery of fetoplacental units and within the vasculature of the most severely affected fetoplacental tissue/placental disc and uterus. This is partly in agreement with cellular investigations of C. jejuni, which supports a transcellular translocation. 5 Similarly, there is agreement with findings from pregnant guinea pig studies, where C. jejuni IA3902 was identified within the cytoplasm of trophoblasts, within phagocytes, in extracellular spaces surrounding trophoblasts in areas of placentitis and necrosis, and occasionally surrounding uteroplacental arteries. 7 The presence of C. jejuni within murine trophoblasts and vasculature of the fetoplacental unit is also consistent with the presence of the organism within trophoblasts and blood vessels of infected placenta in sheep. 11 In that same work, Hedstrom et al 11 proposed trophoblast necrosis with release of the organism to the chorionic villi and vasculature leading to subsequent hematogenous dissemination to the fetus.
There was strong positive staining for neutral mucin (PAS) in locations of the fetoplacental unit where C. jejuni was located but no staining with Alcian blue (acid mucin) in these same tissues. Similarly, there was strong positive staining for L-fucose (lectin) and iron (Perls’s iron) in foci of the fetoplacental unit that were heavily colonized with C. jejuni IA3902. This suggests that the organism may have affinity for foci in infected placental discs where there is abundant neutral mucin (at the periphery of the placental discs both within the cytoplasm of trophoblasts and extracellularly), iron (within vascular sinusoids of placental discs), and L-fucose (cytoplasmic and membrane staining of cells and extracellularly at the periphery of placental discs). Previous work has found that mucin, iron, and L-fucose are trophic factors for C. jejuni IA3902, and the findings in this murine study are consistent with data from guinea pig studies. 8
Exudation of leukocytes is also a feature of the reproductive tract infection with Campylobacter. 11 In this study, there was a statistically significant neutrophilic infiltrate within the endometrial wall (endometritis). This is in agreement with sheep studies using this pathogen, where histological lesions included suppurative necrotizing placentitis and/or endometritis, 17 and also in agreement with guinea pig studies where uterine lesions, although variable, consisted of suppurative endometritis, metritis, and hemorrhage. 7
Currently, the most commonly used model to assess the reproductive consequences of C. jejuni infection is the guinea pig model, where IP inoculation of IA3902 resulted in fetoplacental infection, abortion, and demonstrable metritis and placentitis in 100% of challenged animals. After oral challenge in pregnant guinea pigs, 7 of 10 placental samples had high numbers of C. jejuni, 7 with each animal from which C. jejuni was isolated consistently demonstrating metritis and placentitis. In this murine study, C. jejuni was recovered from the fetoplacental unit following oral inoculation in 2 of 7 BALB/c and 3 of 8 CD-1 mice, which was relatively low, yet there was 100% recovery of the organism from the fetoplacental disc of IP-inoculated mice from both the BALB/c and CD-1 groups. Since the outcomes following IP inoculation were similar, the disparity in results between guinea pigs and mice following oral inoculation is likely associated with differences in the ability of C. jejuni to colonize or translocate the intestine in these species. There was 95% isolation of C. jejuni from the feces in orally challenged mice and 95% isolation from IP-challenged mice at necropsy, which suggests similar widespread intestinal colonization in both these strains of mice. Most mice had moderate bacterial counts (50–1000 CFU) in the feces 1 week after inoculation, although BALB/c inoculated IP had high counts (>1000 CFU). So, although bacteremia in mice following oral inoculation has been demonstrated, 23 the main problem with the murine model using oral challenge may lie with establishing sufficient colonization in the gut to facilitate adequate translocation to the bloodstream. Oral inoculation would better replicate natural disease. To this end, the mouse model could be further developed, as murine intestinal colonization by Campylobacter sp. has been demonstrated to be improved by using germ-free mice, mice with humanized microbiota, or vancomycin pretreated mice. 20 Future studies could also investigate the reproductive pathology of C. jejuni IA3902 in Sigirr-deficient (–/–) mice. These mice are deficient in Single IgG IL1 Related Receptor (SIGIRR) and have been proposed as a model for C. jejuni pathogenesis as they develop significant intestinal inflammation in response to colonization by C. jejuni. 19
The 2 main limitations of this study were the relatively small number of pregnant mice of each of the 2 strains used and the slightly variable gestation lengths. From the results of this study, it appears that an appropriate murine model for studying the reproductive tract pathology of C. jejuni IA3902 would be the use of 14-day pregnant BALB/c mice, using a higher inoculum (eg, 1 ml of C. jejuni IA3902 at 109 CFU/ml) injected IP around day 14 or 15 of gestation with termination of the study within 3 to 4 days (day 18 of gestation). While 14-day pregnant CD-1 mice may also be a viable model, CD-1 mice in this study had larger litter sizes and did not have dead pups or evidence of fetal resorption; therefore, there was a larger range of lesions in BALB/c mice. It would be useful to conduct further investigations using larger sample sizes and larger inoculums via the oral route and with specifically delineated gestation lengths (eg, gestation at 10, 12, 14, and 16 days). Termination at 18 days’ gestation is suggested before the onset of live births. Two BALB/c mice had gross and histological evidence of necrotizing hepatitis, which was supported by microbial culture of the liver in both cases. In addition, there was histological evidence of cholecystitis. Microbial culture of the murine liver and bile should therefore be included in future studies. Overall, this study demonstrated that there is sufficient evidence to advance the murine pregnant mouse model for investigations into the reproductive pathology of C. jejuni IA3902.
Conclusion
C. jejuni IA3902 is the major zoonotic abortifacient pathogen in the sheep industry in the United States. The lack of an effective chemotherapeutic agent or effective vaccine against the organism underscores the value of establishing appropriate animal models like the murine model to facilitate much-needed research. The results from this study indicate that C. jejuni IA3902, when inoculated orally into BALB/c and CD-1 mice, has affinity for the murine reproductive tract, specifically the fetoplacental unit. The gross reproductive lesions include endometritis, placentitis, fetal resorption, fetal death, and birth of dead pups. There is correlation with the presence of gross lesions and high microbial counts. The murine fetoplacental unit contains trophic growth factors for C. jejuni IA3902, including L-fucose, mucin, and iron in locations where the organism was confirmed by immunohistochemistry for major outer membrane protein of C. jejuni, specifically at the periphery of the fetoplacental disc. In this location, the organism causes focal to locally extensive neutrophilic placentitis with intralesional bacterial colonies, as identified by histopathology and supported by high recovery (>1000 CFU) of the organism by microbial culture, conclusively meeting the end-point criteria in this study.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.