
Abstract
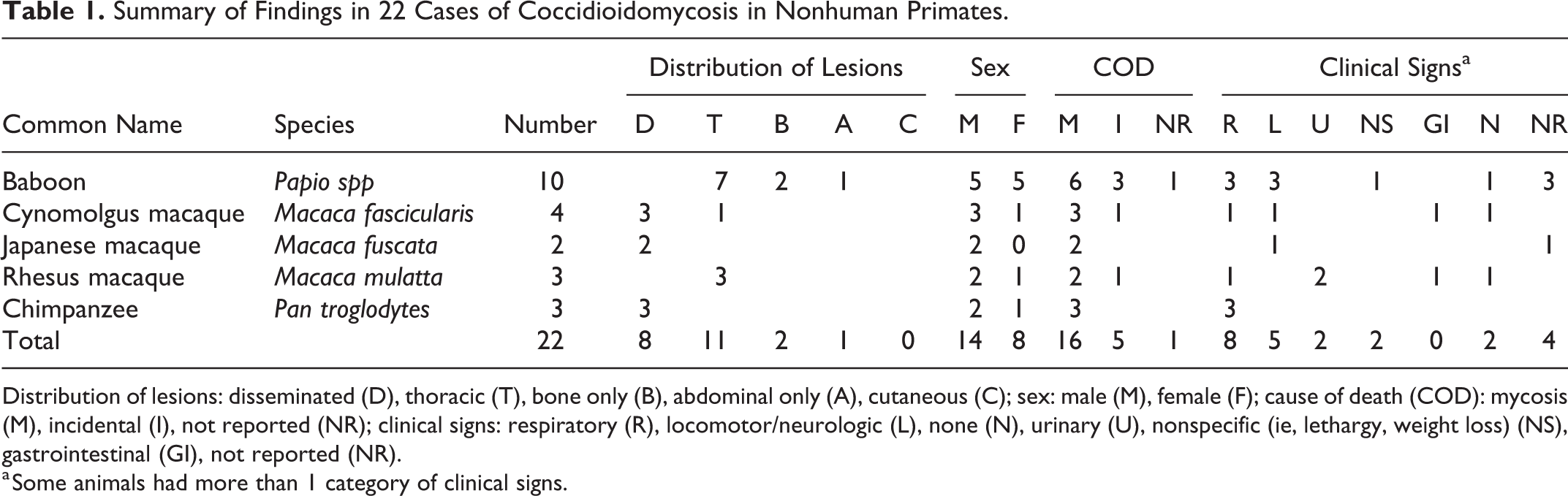
Coccidioidomycosis in nonhuman primates has been sporadically reported in the literature. This study describes 22 cases of coccidioidomycosis in nonhuman primates within an endemic region, and 79 cases of coccidioidomycosis from the veterinary literature are also reviewed. The 22 cases included baboons (n = 10), macaques (n = 9), and chimpanzees (n = 3). The majority died or were euthanized following episodes of dyspnea, lethargy, or neurologic and locomotion abnormalities. The lungs were most frequently involved followed by the vertebral column and abdominal organs. Microscopic examination revealed granulomatous inflammation accompanied by fungal spherules variably undergoing endosporulation. Baboons represented a large number of cases presented here and had a unique presentation with lesions in bone or thoracic organs, but none had both intrathoracic and extrathoracic lesions. Although noted in 3 cases in the literature, cutaneous infections were not observed among the 22 contemporaneous cases. Similarly, subclinical infections were only rarely observed (2 cases). This case series and review of the literature illustrates that coccidioidomycosis in nonhuman primates reflects human disease with a varied spectrum of presentations from localized lesions to disseminated disease.
Coccidioidomycosis, also known as valley fever, California fever, or San Joaquin Valley fever, is a fungal disease that is endemic within the Western Hemisphere in semiarid regions known as the Lower Sonoran Life Zone. This zone within the United States encompasses the southern parts of Texas, Arizona, New Mexico, and much of central and southern California. 22,28 Endemic regions outside of the United States include semiarid regions of Mexico, especially northern Mexico, as well as smaller endemic foci within Central and South America. 28 It is caused by a geophilic dimorphic fungus of which 2 nearly identical species are recognized, Coccidioides immitis and Coccidioides posadasii. C. posadasii was recently proposed as a new species based on genetic and phenotypic analysis 22 and is generically known as the non-California species. C. immitis is almost exclusively found in the endemic areas of California while C. posadasii is found throughout the Lower Sonoran Life Zone; this geographic distribution can often help identify the etiologic agent. 28
As dimorphic fungi, these organisms change between a parasitic (spherule) and a saprophytic hyphal (mold/mycelial) form depending upon their environment. They exist in a saprophytic mycelial form when in the soil, producing arthrospores (arthroconidia) that are aerosolized when the soil is disturbed by human activities or natural forces. These arthrospores cause infection in humans and other mammals by transforming into pathogenic spherules upon inhalation. 37 After reproducing by endosporulation, the spherules rupture, releasing hundreds of endospores into the surrounding tissue. These released endospores also mature into new spherules, and the cycle continues until host control is achieved. 25 These spherules can often be seen on histologic examination of infected tissues.
Cases of coccidioidomycosis in humans are primarily characterized by an acute self-limiting respiratory disease that is generally asymptomatic or subclinical. 51 Clinical disease classically presents as respiratory disease characterized by dyspnea, coughing, and respiratory distress. Typical gross pathological changes include the presence of pulmonary nodules and cavities throughout the lung tissue, often accompanied by thoracic lymph node enlargement. 55 In less than 2% of cases, hematogenous or lymphatic dissemination leads to chronic, progressive disease 18 involving the viscera, skin, soft tissue, bone, and the central nervous system. 1 Risk factors for dissemination include immunosuppression secondary to human immunodeficiency virus (HIV) or organ transplantation, ethnic and sex predispositions, and occupation. 1 The incidence of coccidioidomycosis reported in humans has increased substantially between the years of 1997 and 2011. 14
Focal, regional, and systemic cases have been reported in many species of nonhuman primates (NHPs), exhibiting a course of disease similar to that seen in humans. These species include geladas, 46 baboons, 3,27,48,56 mandrills, 33,47 woolly monkeys, 46,47 chimpanzees, 6,29,30,32,37,39,47,53 macaques, 2,8,10,26,41,47 mangabeys, 44 guenons, 20,47 spider monkeys, 20,47 gorillas, 4,31,40,47 capuchins, 40 squirrel monkeys, 47 and lemurs. 9 Nonhuman primates appear to be particularly vulnerable to arthrospore exposure in known endemic regions. 5,15,18
An overview of the pathology associated with Coccidioides spp infection in NHPs has not been previously reported. Here we provide a review of the cases previously described,* and describe a retrospective series of 22 naturally acquired cases that were diagnosed at multiple pathology services, including the United States Army Medical Research Institute of Infectious Disease (USAMRIID), Southwest National Primate Research Center at the Texas Biomedical Research Institute (SNPRC), and Texas Veterinary Pathology Associates (TVP). SNPRC and TVP are located in San Antonio, Texas, which is at the very eastern boundary of the geographical endemic distribution of C. immitis/C. posadasii. 12
Materials and Methods
A search for histologically confirmed cases of coccidioidomycosis within the pathology records at the authors’ respective pathology services was performed. Search terms included coccidioidomycosis and Coccidioides. The cases identified from those search results are presented.
All of the cases in this series were housed for at least a short time in indoor/outdoor group housed facilities at primate research centers in San Antonio and southeast Texas. In addition, all cases were in the endemic area when diagnosed with coccidioidomycosis with the exception of case 15 that was temporarily housed in this endemic region for approximately 14 weeks.
The cases originating from SNPRC, San Antonio, Texas, represent a population of more than 2000 NHPs, comprising approximately 1500 baboons, 500 macaques, and 150 chimpanzees. The TVP cases originated from multiple primate facilities; TVP is a diagnostic pathology company that provides pathology services to multiple primate facilities in San Antonio and southeast Texas. The USAMRIID case was a primate received from one of the normal vendors used for the institute. Outdoor access when provided to animals in this case series consisted of enclosures with indoor and outdoor areas or large multiacre outdoor corrals. All animal care and research were conducted under an Institutional Animal Care and Use Committee–approved protocol in compliance with the Animal Welfare Act, Public Health Service (PHS) policy, and other federal statutes and regulations relating to animals and experiments involving animals. The facilities where this research was conducted are accredited by the Association for Assessment and Accreditation of Laboratory Animal Care, International and adhere to principles stated in the Guide for the Care and Use of Laboratory Animals, National Research Council, 2011.
Tissues were collected for histologic evaluation as required for diagnosis and fixed in 10% neutral buffered formalin, processed conventionally, embedded in paraffin, cut at 5 μM and stained with hematoxylin and eosin or other stains, including periodic acid–Schiff reaction (PAS) and Gomori’s methenamine silver stain (GMS) using standard techniques as needed for diagnosis. When indicated, individual tissues were frozen in liquid nitrogen and stored at –80°C, fixed in 2% glutaraldehyde for electron microscopy, placed in normal saline or transport medium for cytogenetic evaluation, cultured for bacteria and viruses, or frozen in Optimal Cutting Temperature Compound (Tissue-Tek) compound for frozen sectioning. Further evaluation using immunohistochemistry was performed as required. Necropsies were performed by a board-certified pathologist or trained laboratory animal veterinarian. Histologic evaluation of tissues was performed by board-certified veterinary pathologists.
Electron Microscopy
Samples of formalin-fixed tissue from case 15 (Suppl. Table S1) were retrieved and prepared for electron microscopy. Briefly, tissues were postfixed in osmium tetroxide, dehydrated in ethanol, infiltrated in a 1:1 solution of propylene oxide/Epon resin, and embedded in Epon resin. Embedded tissues were thin sectioned (∼70-nm sections), and sections were mounted on copper mesh or slot grids with a plastic support film. Sections were contrasted with uranyl and lead salts prior to observation. Sections were observed on a JEOL 1011 (Peabody, MA) transmission electron microscope at 80 kV and digital images were acquired using a Orca HR digital camera (Hamamatsu Corporation, Bridgewater, NJ) controlled by AMT image acquisition software.
Immunohistochemistry
For immunohistochemical identification of Coccidioides sp, an immunoalkaline phosphatase technique using a rabbit polyclonal antibody raised against C. immitis was used. This polyclonal antibody is reactive against Coccidioides sp and, based on analysis of tissues with known infections, does not cross-react with Blastomyces, Cryptococcus, Histoplasma, Rhizomucor, or Candida spp.
In addition, a polyfungal analysis was performed using a polyfungal goat serum that was raised against a shared epitope of Aspergillus fumigatus, Aspergillus niger, and Aspergillus flavus. Based on analysis of tissues with known infections, this goat serum is known to react with Blastomyces, Coccidioides, Histoplasma, Candida, Fusarium, Apophysomyces, Basidiobolus, Exserohilum, and Microsphaeropsis with variable reactivity to Zygomycetes. It is not known to cross-react with Cryptococcus.
Polymerase Chain Reaction
As previously described, 43 DNA was purified from paraffin-embedded tissue, and polymerase chain reaction (PCR) amplification was performed using a panfungal primer set, ITS3/ITS4, which amplifies the ITS2 region of the ribosomal DNA, to identify Coccidioides sp genomic material.
Literature Review
A search of the PubMed and PrimateLit databases was conducted using the following key words: primate, coccidioidomycosis, and Coccidioides. In addition, references from the articles obtained in the initial search were examined to find other reports not included in the initial database query.
Results
The pathology database search revealed 22 cases: 16 from SNPRC, 5 from TVP, and 1 from USAMRIID. A summary of the cases by species is listed in Table 1; Supplemental Table S1 contains a complete list of the clinical and pathology findings in each case.
Summary of Findings in 22 Cases of Coccidioidomycosis in Nonhuman Primates.
Distribution of lesions: disseminated (D), thoracic (T), bone only (B), abdominal only (A), cutaneous (C); sex: male (M), female (F); cause of death (COD): mycosis (M), incidental (I), not reported (NR); clinical signs: respiratory (R), locomotor/neurologic (L), none (N), urinary (U), nonspecific (ie, lethargy, weight loss) (NS), gastrointestinal (GI), not reported (NR).
a Some animals had more than 1 category of clinical signs.
The 16 cases found within SNPRC’s records were diagnosed sporadically over a period of 30 years and consisted of 10 baboons, 3 chimpanzees, and 3 macaques; males and females were equally represented. All cases were born at the facility or originated within endemic regions of the United States. The 5 cases within TVP’s records were diagnosed in a period of 10 years and consisted of 5 macaques, 3 rhesus macaques, and 2 cynomolgus macaques. The single case found in USAMRIID’s records (records that covered a period of 40 years) was a cynomolgus macaque.
Clinical Findings and Gross Lesions
Baboon
Ten cases were discovered in baboons, including cases 7 and 8 that were described in previously published case reports. 3,48 The clinical history was not available for 3 of the 10 baboon cases (cases 2, 8, and 9). Among the 7 cases in which clinical history was available, 3 developed respiratory signs (cases 1, 3, and 10), 3 had locomotion deficiencies (cases 4, 6, and 7), and 1 had no observed clinical findings (case 5). The respiratory signs consisted of coughing (n = 2) or labored breathing (n = 1). Motor deficiencies included hindlimb paresis to paralysis (n = 2) and weakness and reluctance to move (n = 1). The animal with no observed clinical findings was euthanized due to reasons unrelated to fungal infection, and the lesions attributable to fungal infection in this case consisted of a mass attached to the wall of the right main bronchus, which extended to the subjacent lymph nodes and lung. These gross findings were considered incidental and unrelated to the reasons for euthanasia.
Lesions were most commonly noted in thoracic organs and less often the vertebral column and abdominal cavity. Lesions in the thoracic organs most commonly included the lungs (n = 7), regional lymph nodes (n = 3), heart (n = 2), trachea (n = 1), and esophagus (n = 1). Gross lesions within the thoracic organs included adhesions, granulomas, abscesses, ulcers, or masses. In some cases, the lungs had dark brown to black discoloration with black to brown fluid that extended throughout all lung lobes on cut section. In other cases, the lungs were firm, mottled gray-red-pink, with suppurative exudate expressed from the airways on cut section. Thoracic lesions were absent in 3 baboons (cases 4, 6, and 7). Two of these presented with hindlimb paresis or paralysis, and all 3 had locally extensive areas of granulomatous inflammation and necrosis that affected the vertebrae, adjacent soft tissue, and spinal cord. Case 4 had an irregularly thickened thoracic vertebrae. Case 6 lacked pulmonary lesions and was the only baboon with lesions involving abdominal viscera, with granulomas in the spleen and associated lymph nodes.
Chimpanzee
The 3 chimpanzees diagnosed with coccidioidomycosis all presented with respiratory signs, including tachypnea. In addition, case 11 also presented with ascites, and this case was clinically diagnosed as C. immitis/C. posadasii by fungal culture of a thymic cyst biopsy. Despite itraconazole treatment, this chimpanzee’s condition continued to decline and it was euthanized.
The infection was disseminated to multiple organs in all 3 chimpanzees, including the lungs (n = 3), pleura (n = 1), spleen (n = 3), lymph nodes (n = 3), liver (n = 3), thoracic lymph nodes (n = 1), diaphragm (n = 1), peritoneum (n = 1), and kidneys (n = 1). The gross lesions in these organs were as described above and appeared as granulomas and abscesses. In addition, excessive fluid was observed in the abdomen (n = 2) and pericardium (n = 1), and was characterized as clear, seropurulent with fibrin tags, or suppurative.
Macaque
Nine macaques were diagnosed with coccidioidomycosis: 3 rhesus macaques, 4 cynomolgus macaques, and 2 Japanese macaques. Clinical signs varied and body systems affected included respiratory (n = 2), urinary (n = 2), locomotor/nervous (n = 2), and gastrointestinal (n = 2). The clinical signs in individual macaques were described as coughing and gagging, urinary incontinence and rear-limb paresis, open-mouth breathing and collapse, ruptured bladder, bloody diarrhea with bloodwork consistent with renal disease, diarrhea and dehydration, subcutaneous edema, hindlimb paresis, or incontinence. One macaque (case 15) had no clinical signs attributable to fungal infection. One macaque (case 16) was also seropositive for Trypanosoma cruzi.
The distribution of lesions was limited to thoracic organs in 4 macaques (cases 15, 20, 21, and 22) and was disseminated (intrathoracic and extrathoracic) in 5 macaques (cases 14, 18, 19, 16, and 17). The specific lesion location was not available for all animals but was reported in 6 of 9 macaques as affecting the lungs (n = 5), spleen (n = 2), liver (n = 2), kidneys (n = 3), or muscle (n = 1). The lung lesions in macaques were described as having yellow purulent foci (abscesses), nodules (Fig. 1 and 2), miliary foci, or areas of dark discoloration (infarction). Subcutaneous and scrotal edema, hydropericardium, and ascites were additional lesions.
Coccidioides spp infection, thorax and lungs, rhesus macaque.
Case 15 was administered 1608 plaque-forming units (PFUs) of monkeypox virus by aerosol and was euthanized at the end of the study on day 29 postinfection. Clinical signs attributable to monkeypox were evident by day 4 postinfection, including pyrexia, lymphadenopathy, and mild cutaneous pox lesions. Clinical respiratory signs were not observed before or after monkeypox challenge. Gross examination revealed few faintly visible resolving pox lesions that consisted of 1- to 2-mm whitish/brown discolored spots on the skin of the axillary and inguinal areas. The left lung was diffusely firm, while the right lung contained few 5- to 10-mm pale firm foci affecting 10% of the apical and diaphragmatic lobes. All examined lymph nodes were mildly enlarged, and the spleen was markedly enlarged with rounded edges and prominent white pulp on cut surface. All noted clinical and gross necropsy abnormalities in this case, except for the lung lesions, were attributable to the monkeypox infection and were typical for aerosol exposure to this virus. 24
Histopathology
Histologic findings were similar in all organs for all species and were characterized by multifocal areas of granulomatous inflammation composed of numerous multinucleate giant cells admixed with lymphocytes, plasma cells, histiocytes, and viable and degenerate neutrophils and eosinophils as well as fungal spherules that were 25 to 30 microns in diameter with a 2- to 3-micron-thick wall. The spherules rarely underwent endosporulation; endospores filling the spherules were 2 to 5 microns in diameter (Fig. 3). The spherules were often contained within the cytoplasm of the multinucleate giant cells and were often degenerate. Within pulmonary lesions, the inflammatory foci obscured the pulmonary architecture, and the bronchioles and alveoli were often dilated and filled with inflammatory cells, cellular debris, edema, and moderate numbers of fungal spherules. In some cases, the alveoli were occasionally lined by plump cuboidal pneumocytes (type 2 pneumocyte hyperplasia). Within the bone and associated soft tissues, the foci of granulomatous inflammation were accompanied by necrosis and degeneration of the mature bone.
The distribution was classified as thoracic, disseminated, bone only, or abdominal based on location of histopathologic lesions. The definition of disseminated distribution was based on finding histologic lesions in pulmonary and nonpulmonary locations, including visceral organs and/or bone. Thoracic distribution was the most common and was found in 11 cases (7 baboons, 1 cynomolgus macaque, and 3 rhesus macaques). Disseminated distribution occurred in 8 cases (3 chimpanzees, 3 cynomolgus macaques, and 2 Japanese macaques). Bone only and abdominal were the least frequent distributions and occurred in 2 animals (cases 4 and 7) and 1 animal (case 6), respectively, and all 3 were baboons.
Concomitant findings in some animals included amyloidosis in 6 animals (2 baboons, 3 rhesus macaques, and 1 cynomolgus macaque) in varying locations, including kidney, lymphoid organs, and pancreas. Pyelonephritis and myocardial infarction were present in another case, and both were considered unrelated to mycosis. The gastrointestinal, endocrine, and reproductive systems did not reveal other lesions attributable to Coccidioides spp infection, except for those reported above.
Confirmatory Tests
Routine histopathologic examination supported by special stains, including PAS reaction (Fig. 4), provided a diagnosis in most cases. Other confirmatory tests used to characterize the cases in this series included tissue culture, polyfungal immunohistochemistry, agent-specific immunohistochemistry, PCR, and electron microscopy. Fungal culture was used to confirm the diagnosis of coccidioidomycosis in 2 chimpanzees from a biopsy (case 11) and from tissues collected after death (case 13).
Immunohistochemistry was performed on the 15 cases in which paraffin-embedded tissue was available, using antibodies specific for Coccidioides spp as well as polyfungal antibodies. Both methods identified Coccidioides spp in 14 of the 15 cases (cases 1–13, 15, 19, and 17). Fungal antigen was located in spherules located within multinucleate giant cells as well as extracellular free spherules.
PCR testing was conducted on paraffin-embedded tissue from these 15 cases using Coccidioides sp.–specific primers and was positive in 5 cases (cases 2, 3, 12, 13, and 19). The assay did not distinguish C. immitis and C. posadasii.
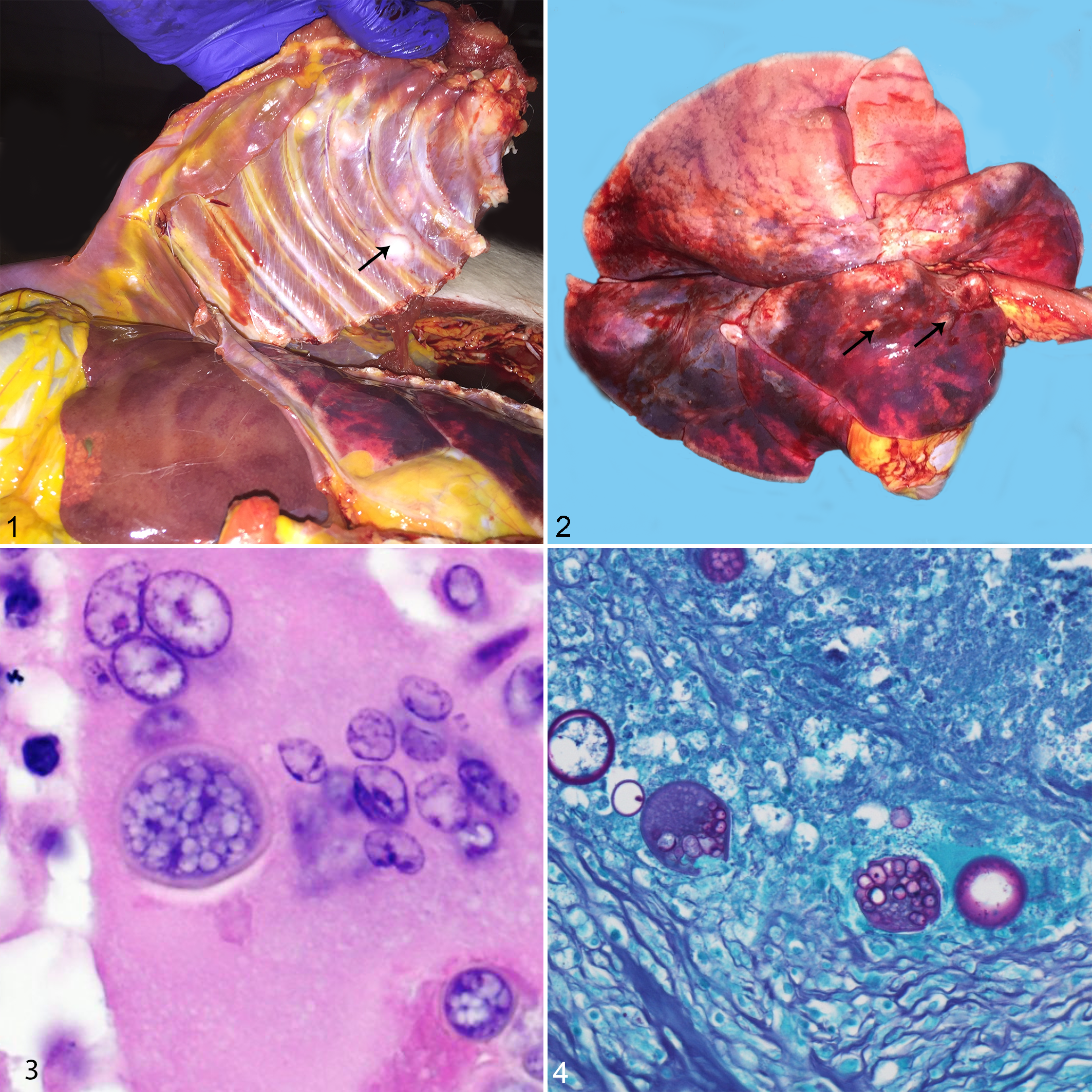
Electron microscopy on the lung tissue from case 15 identified mature and immature spherules that were approximately 15 to 20 microns in diameter and had a 1-micron-thick electron-dense wall (Fig. 5). Mature spherules contained endospores while immature spherules lacked endospores (Fig. 6).
Coccidioides spp infection, lung, cynomolgus macaque, case 15. Transmission electron microscopy.
Literature Review
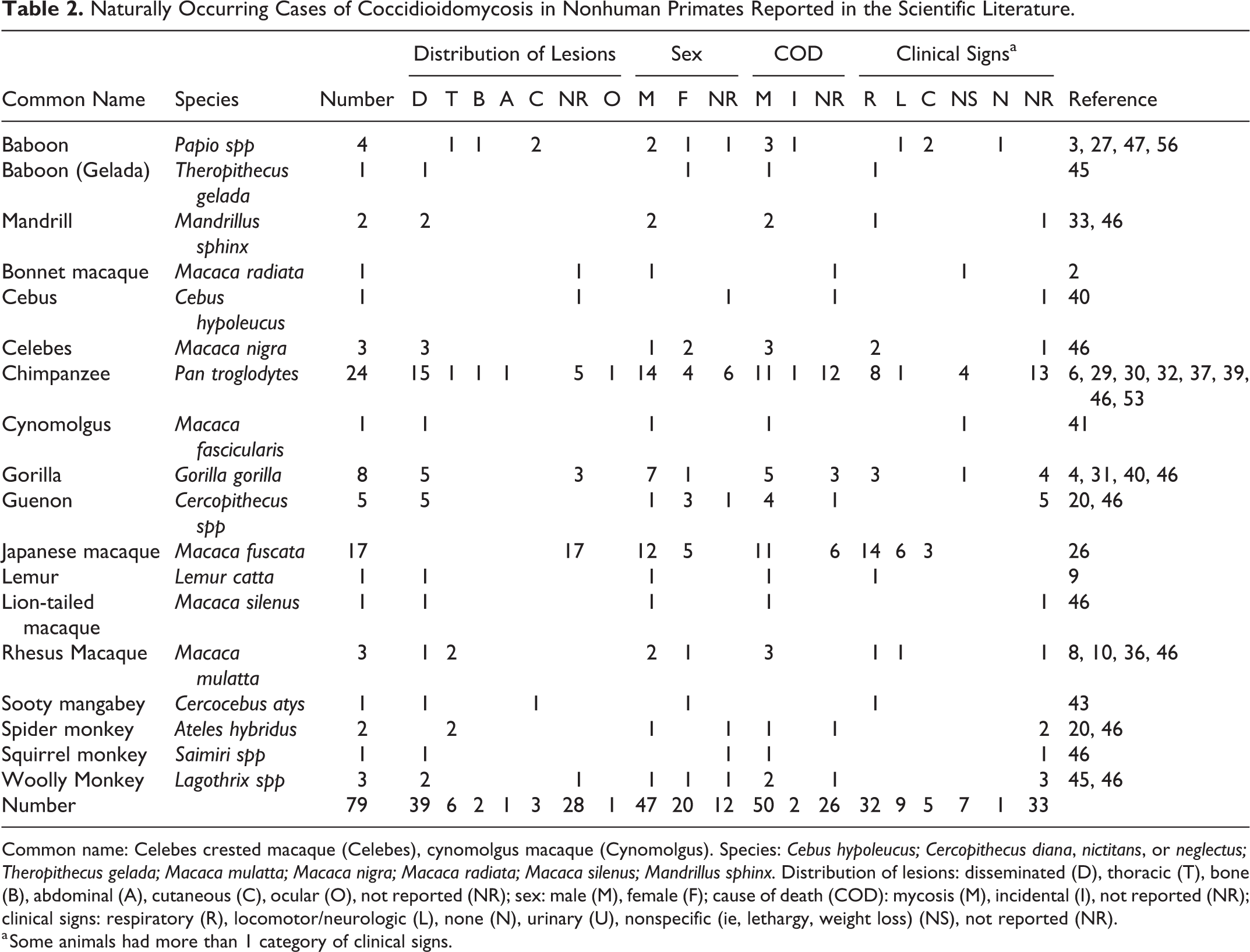
A search within the PubMed database revealed 79 naturally occurring cases of coccidioidomycosis in NHPs (48 male, 19 female, 12 not stated) (Table 2). Of these, 3 cases were also reported above. The most common species affected were macaques (n = 26), chimpanzees (n = 24), and gorillas (n = 8). The clinical signs were described in 46 cases and affected the respiratory system (n = 32), skin (n = 5), and vertebrae and/or neurological system (n = 9) or were nonspecific such as weight loss (n = 7) or abdominal distension (n = 3).
Naturally Occurring Cases of Coccidioidomycosis in Nonhuman Primates Reported in the Scientific Literature.
Common name: Celebes crested macaque (Celebes), cynomolgus macaque (Cynomolgus). Species: Cebus hypoleucus; Cercopithecus diana, nictitans, or neglectus; Theropithecus gelada; Macaca mulatta; Macaca nigra; Macaca radiata; Macaca silenus; Mandrillus sphinx. Distribution of lesions: disseminated (D), thoracic (T), bone (B), abdominal (A), cutaneous (C), ocular (O), not reported (NR); sex: male (M), female (F); cause of death (COD): mycosis (M), incidental (I), not reported (NR); clinical signs: respiratory (R), locomotor/neurologic (L), none (N), urinary (U), nonspecific (ie, lethargy, weight loss) (NS), not reported (NR).
a Some animals had more than 1 category of clinical signs.
Twenty-eight reports did not include any gross pathologic findings because the animal was alive at the time of study publication, a necropsy was not performed, or findings were not included in the description. Gross lesions were found in the lung (n = 21), spleen (n = 10), liver (n = 7), bone (n = 7), lymph nodes (n = 7), heart (n = 5), esophagus (n = 4), pleura (n = 4), trachea (n = 3), kidney (n = 3), testis (n = 2), peritoneum (n = 2), diaphragm (n = 2), pancreas (n = 1), colon (n = 1), retina (n = 1), brain (n = 1), and meninges (n = 1). The distribution of lesions was disseminated (n = 39), intrathoracic (n = 6), bone (n = 2), abdominal (n = 1), ocular (n = 1), or cutaneous (n = 3).
Of particular note, 3 chimpanzees lacked involvement of the respiratory system. These had a granulomatous abdominal mass encasing the pancreas and the abdominal aorta (case 28) and an osteolytic mass incorporating the 11th, 12th, and 13th thoracic vertebra accompanied by clinical neurologic signs (case 34), as well as bone lesions in the atlas, its corresponding occipital condyle, and a rib, with soft tissue involvement limited to a perforated esophagus (case 15).
Experimental infection of 93 rhesus macaques with Coccidioides was reported in 5 studies published between 1953 and 1966 (Table 3), using aerosolization, intratracheal injection, or natural exposure. 5,7,16 –18 The naturally exposed cases were infected by housing the animal in an area that was known to be infested. 18 When arthrospores were administered via intratracheal injection, all monkeys showed evidence of pulmonary infiltration. The pulmonary lesions varied from few focal lesions to extensive pulmonary destruction, and 1 study attributed the severity of lesions to the administered dose. 5 The disease progressed from minor inflammation of the lungs and enlarged lymph nodes to necrotic, granulomatous nodules and lymph node inflammation. Within 15 days of exposure, the animals developed nodules within the lungs, regional lymph nodes, liver, and spleen, as well as fibrinous adhesions between organs. In macaques infected with aerosolized arthrospores, the gross lesions were consistent with natural cases. 7
Reports of Experimental Infection of Rhesus Macaques With Coccidiodes sp.

Discussion
We report 22 natural cases of coccidioidomycosis in NHPs and review natural and experimental cases of coccidioidomycosis in the nonhuman primate literature. As a group, primates appear particularly susceptible. 50 The cases were distributed between males and females of all ages.
Pulmonary and disseminated histologic lesions consisted of pyogranulomas or granulomas. There are frequently aggregates of intermixed neutrophils resulting from the initial reaction to the endospores released from matures spherules. The lesion is thought to form a granuloma or pyogranuloma as it matures and is composed of the common components of a granulomatous reaction, including giant cells. 11
The clinical signs and pathology observed in the naturally occurring cases reported here are generally consistent with those cases that have been described previously in the literature. Among the cases reported in the literature, overall, the species with the most reported cases were macaques (n = 34), chimpanzees (n = 27), baboons (n = 12), and gorillas (n = 8). All other species had few reports, and almost all involved disseminated disease. The exception was the spider monkey, which had 2 reported cases, both with lesions confined to the thorax. Macaques and chimpanzees frequently had disseminated disease (21/34 and 18/22 with lesions reported, respectively). None of the 12 affected baboons had disseminated disease, and lesions were confined to the thorax (n = 7), vertebrae and adjacent soft tissue (n = 2), skin and subcutis (n = 2), or spleen and associated lymph node (n = 1). The authors suspect that 1 or both of the baboon cases with cutaneous involvement 27,56 were misdiagnosed as both were diagnosed in the United Kingdom, and neither had a reported history of travel to the Americas. The reason that baboons had a difference in distribution of lesions was not apparent, and the histologic nature of the lesions described in the reports was not different.
Despite the presence of gross and histologic lesions at postmortem examination, no clinical signs attributed to coccidioidomycosis were observed in 2 NHPs in our case series, a cynomolgus macaque (case 15) and a baboon (case 5). Similarly, serologic surveillance studies have shown that many infections in humans and animals are asymptomatic. In a survey of dogs in Arizona, 70% of seropositive dogs exhibited no sign of disease. 49 Similarly, 100% of a group of naturally exposed monkeys developed subclinical infections after being contained in outdoor housing in a river basin for a year in an endemic region (Tucson, Arizona). None of these monkeys exhibited clinical signs, but all were positive by a coccidioidin skin test and complement fixation test; 40% of the exposed monkeys had histologic lesions. 18 In NHPs housed outdoors at the California Primate Research Center in 1977 to 1978, 4 of 119 (3%) were seropositive; the seropositive cases were attributed to a dust storm that affected the area, as there were no seropositive animals when they were surveyed prior to the dust storm. 2 Approximately 60% of infected humans are asymptomatic; the remainder can develop manifestations that range from mild influenza-like illness to pneumonia. Overall, less than 5% of infected persons have progressive pulmonary infection or extrapulmonary dissemination of the disease. 35
In one of these subclinical cases, case 15, an initial presumptive diagnosis was based on the morphologic appearance of fungal spherules in the tissue section, but additional confirmatory testing was performed because this diagnosis was unexpected because of the geographic location (Maryland) and the absence of clinical signs related to fungal infection. The results all supported the diagnosis of coccidioidomycosis. In this case in addition to the lung lesions, other nonpulmonary lesions were noted, but all of these nonrespiratory clinical and gross necropsy abnormalities were attributable to experimental monkeypox exposure. 24 The source of infection of this case is speculated to be inhalation of dust-borne arthrospores while the animal was temporarily housed in Texas. C. posadasii is the most likely cause based on the geographical location history of this primate and the geographic distribution of this species. Infection of this monkey during this relatively brief period of exposure (14 weeks) is supported by the findings of Converse and Reed 18 that infection could be established after aerosol exposure of rhesus macaques to as few as 10 arthrospores. This pathogen is considered one of the most virulent and highly contagious fungal pathogens, and thus it should only be grown under biocontainment conditions, as infections have occurred following exposure to arthrospores in the laboratory. 54
Animal models of coccidioidomycosis include mice, 19 rats, 21 hamsters, 21 guinea pigs, 57 dogs, 18,23 cattle, 38 and primates. 5,7,16 –18,36 Since NHPs appear to be particularly susceptible to infection and the disease progression parallels that in humans, they appear to be a good model of the disease. Although the mouse is used most extensively as an animal model due to its rapid disease progression and the availability of genetically defined animals, it often requires euthanasia to collect sufficient tissue samples, and its small size makes it difficult to assess individual organ pathology. 15 There are also substantial pharmacokinetic discrepancies between mice and humans, complicating the study of vaccines and treatments. 52 Both the mouse and the hamster rapidly develop disseminated disease regardless of the method of inoculation, and death occurs within 4 weeks of inoculation in 83.3% of hamsters. 21
Infection of NHPs via natural as well as experimental infection closely resembles the course of disease in humans. The gross lesions in macaques experimentally infected with aerosolized arthrospores are also consistent with natural cases. 7 The predictable disease progression with both aerosol and intratracheal inoculation makes NHPs good candidates for modeling infection with C. immitis/C. posadasii. Additional advantages of using NHPs as a model of coccidioidomycosis include a slower disease progression and more variations in disease forms; this may allow the use of fewer animals and lead to more targeted treatment studies.
Dogs and cats are also affected by this mycotic infection, and comparisons to the disease in humans and NHPs can be made. Similar to humans and NHPs, infections in dogs are predominantly limited to the lungs and associated lymph nodes. Disseminated infection in dogs has been recognized in 20% of the cases, and locations affected include the bones, joints, and lymph nodes, with bone being the most common site. In dogs, in contrast to other species including mice, hamsters, humans, and NHPs, the appendicular skeleton is more often affected than the axial skeleton. 25,50 The brain lesions in dogs are typically granulomatous mass lesions while in humans, meningeal lesions typically occur. 50 Cats, like humans, develop cutaneous lesions more commonly than other domestic species. Dissemination occurs in 50% of feline infections, and the skin is the most common site of disseminated infection. 25
When comparing the disease in NHPs to humans, the disease in both includes lung cavitation and bone infections nearly exclusively within the axial skeleton; infection of the appendicular skeleton is rarely reported. However, NHPs do differ from humans in the usual lack of skin and subcutaneous lesions, which occur in more than 65% of cases of disseminated disease in humans. 19,26 The Japanese macaque is a notable exception, as 3 of 17 had a clinical history of cutaneous involvement that was identified by culture to be Coccidioides spp. 26 In humans, the skin lesions are often associated with erythema nodosum, 35 which has not been described in animals.
Morphology of the Coccidioides spherules in histological sections is unique, and a strongly presumptive diagnosis can be made with histopathology alone. The diagnosis of coccidioidomycosis is not always straightforward if there is an absence of histologically apparent arthrospores; available confirmatory tests include fungal culture, immunohistochemistry (IHC), PCR, and in situ hybridization (ISH). C. immitis/C. posadasii exists in 2 forms: a hyphal form and a spherule form. The hyphal forms are rare in tissue sections because they develop at room temperature (25°C). The hyphal form has been observed in tissues from humans, especially in cases with large cavitary lesions. 42 The spherules are observed in the tissue phase and only produced in culture under special growth conditions. The presence of misleading Coccidioides elements that may resemble other pathogens such as Blastomyces dermatitidis or hyphomycetes has been reported in tissues from humans. These misleading elements include budding cells; branched, septate hyphae; and moniliform hyphae consisting of chains of thick-walled arthroconidia. 34
Other fungal organisms with similar but distinguishable morphology include Emmonsia spp, Blastomyces dermatitidis, and Rhinosporidium seeberi. The major distinguishing factor of coccidioidal spherules is the formation of thick-walled endospores within the coccidioidal spherule as it matures. B. dermatitidis does not form endospores but forms broad-based buds and is smaller than mature coccidioidal spherules. R. seeberi forms endospores that have thin walls. In addition, rhinosporidial sporangia are much larger than coccidioidal spherules. Emmonsia spp have the closest morphology but do not form thick-walled endospores and instead form fruiting bodies that line the interior of the wall and result in a honeycomb pattern; these are much less distinct than those of Coccidioides spp. In addition, Emmonsia spp adiaspore walls are thicker than Coccidioides sp. 11 Fungal culture, IHC, PCR, and ISH are other methods to provide an unequivocal diagnosis. Fungal cultures must be treated with precautions in the laboratory because the infectious arthroconidia may develop after incubation at room temperature. 25
Attempts to retrieve genomic material from the paraffin-embedded tissues had poor sensitivity; only 5 of 15 were PCR positive for Coccidioides spp. The PCR assay used did not distinguish C. immitis from C. posadasii. The genomes of C. immitis and C. posadasii only differ by several DNA polymorphisms within 2 microsatellite loci, 22 and their differentiation requires fresh unfixed material.
Serology is the primary method for the antemortem diagnosis of coccidioidomycosis in animals, and it uses the same reagents and controls as for serodiagnosis of the disease in humans. The relationship of serology results to clinical disease is not known for NHPs. Seropositive dogs may have clinical or subclinical disease, but seropositivity of cats correlates well with the presence of clinical disease. 25
Coccidioides spp was found as a contaminant of a primary rhesus monkey kidney (RhMK) cell culture obtained from a commercial supplier; the source of this contamination was not identified. 45 It suggests that screening for this organism should be considered for cell lines derived from NHPs.
In the past decade, the number of reported cases in humans has increased, but the reason is uncertain. Contributing factors are thought to include an increased population in the endemic area, climatic change (drought and increasing temperatures in the southwestern United States), dust storms, soil disturbance caused by increased construction activity, growing numbers of humans who are immunocompromised or have other risk factors for severe disease, and immigration of previously unexposed persons from areas where coccidioidomycosis is not endemic. 13,14
In this report, we have characterized the distribution of coccidioidomycosis in terms of anatomic distribution, types of clinical signs, and species of NHPs affected by reviewing past cases in our records as well as those reported in the scientific literature. We found that as a group, NHPs are particularly susceptible to disease, and presentation of coccidioidomycosis parallels disease in humans with presentations from localized clinical infections to disseminated disease as well as subclinical infections. A wide variety of NHP species were represented. Baboons represented a large number of cases in the case series presented here and had a slightly unique presentation with absence of disseminated disease. The reason for the differing presentation in this species is unknown. Cutaneous infections in NHPs were also observed but had a very uncommon lesion distribution. Subclinical infection was also observed, albeit in only a few cases but this may simply reflect the lack of routine antemortem surveillance for this disease. The presence of subclinical infections has the potential to interfere with experimental studies performed in NHPs, and consideration for the presence of this disease for NHPs that are housed, even temporarily, in endemic areas may be warranted. This survey will provide information to help fill the knowledge gap for the wide variety and incidence of presentations of this disease in NHPs, especially if the disease prevalence in other species, including humans, continues to increase.
Supplemental Material
Supplemental Material, DS1_VET_10.1177_0300985818787306 - Coccidioidomycosis in Nonhuman Primates: Pathologic and Clinical Findings
Supplemental Material, DS1_VET_10.1177_0300985818787306 for Coccidioidomycosis in Nonhuman Primates: Pathologic and Clinical Findings by Keith Koistinen, Lisa Mullaney, Todd Bell, Sherif Zaki, Aysegul Nalca, Ondraya Frick, Virginia Livingston, Camenzind G. Robinson, J. Scot Estep, K. Lance Batey, Edward J. Dick, and Michael A. Owston in Veterinary Pathology
Supplemental Material
Supplemental Material, DS2_VET_10.1177_0300985818787306 - Coccidioidomycosis in Nonhuman Primates: Pathologic and Clinical Findings
Supplemental Material, DS2_VET_10.1177_0300985818787306 for Coccidioidomycosis in Nonhuman Primates: Pathologic and Clinical Findings by Keith Koistinen, Lisa Mullaney, Todd Bell, Sherif Zaki, Aysegul Nalca, Ondraya Frick, Virginia Livingston, Camenzind G. Robinson, J. Scot Estep, K. Lance Batey, Edward J. Dick, and Michael A. Owston in Veterinary Pathology
Footnotes
Acknowledgements
The authors gratefully acknowledge Jayson Felty and the TxBiomed library staff and the SNPRC veterinary pathology and clinical staff and numerous contributions of USAMRIID’s pathology personnel, including William Aguilar, Neil Davis, Phil Fogle, Angela Grove, Gale Krietz, and Christine Mech.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Disclaimers
Opinions, interpretations, conclusions, and recommendations are those of the author and are not necessarily endorsed by the US Army.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This investigation used resources that were supported by the Southwest National Primate Research Center (grant P51 RR013986) from the National Center for Research Resources, National Institutes of Health, which are currently supported by the Office of Research Infrastructure Programs through P51 OD011133 as well as the Office of Biodefense Research Affairs (OBRA)/National Institute of Allergy and Infectious Diseases (NIAID) with interagency agreement (A120-B.11) between USAMRIID and NIAID. This investigation was conducted in facilities constructed with support from the Office of Research Infrastructure Programs (ORIP) of the National Institutes of Health through grants C06 RR014578, C06 RR015456, and C06 RR016228.
Supplementary material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.