
Abstract
Chlamydiae have a worldwide distribution causing a wide range of diseases in human hosts, livestock, and companion animals as well as in wildlife and exotic species. Moreover, they can persist in their hosts as asymptomatic infections for extended periods of time. The introduction of molecular techniques has revolutionized the Chlamydia field by expanding the host range of known chlamydial species but also by discovering new species and even new families of bacteria in the broader order Chlamydiales. The wide range of hosts, diseases, and tissues affected by chlamydiae complicate the diagnosis such that standard diagnostic approaches for these bacteria are rare. Bacteria of the Chlamydiales order are small and their inclusions are difficult to detect by standard microscopy. With the exception of avian and ovine chlamydiosis, macroscopic and/or histologic changes might not be pathognomic or indicative for a chlamydial infection or even not present at all. Moreover, detection of chlamydial DNA in specimens in the absence of other methods or related pathological lesions questions the significance of such findings. The pathogenic potential of the majority of recently identified Chlamydia-related bacteria remains largely unknown and awaits investigation through experimental or natural infection models including histomorphological characterization of associated lesions. This review aims to summarize the historical background and the most important developments in the field of animal chlamydial research in the past 5 years with a special focus on pathology. It will summarize the current nomenclature, present critical thoughts about diagnostics, and give an update on chlamydial infections in domesticated animals such as livestock, companion animals and birds, as well as free-ranging and captive wild animals such as reptiles, fish, and marsupials.
Highlights
Chlamydiosis in animals can range from asymptomatic infection to severe diseases with life-threatening illnesses Zoonotic infections due to Chlamydia abortus and Chlamydia psittaci are well-known Chlamydia-related bacteria might be pathogenic to animals and pose zoonotic risks Chlamydia spp. are either endemic or epidemic in different hosts Studies of new hosts have uncovered new chlamydial pathogens or expanded host ranges for existing species Virtually all chlamydial species might easily and frequently cross host barriers Characteristics of Chlamydia-induced inflammatory changes depend on the chlamydial species, the infected host and the tissue affected
The Chlamydiales—An Introduction
The members of the Chlamydiales order are fascinating Gram-negative, obligate intracellular bacteria sharing a unique biphasic developmental cycle while multiplying in eukaryotic hosts. The order is composed, to date, of 9 families of which members infect humans, wild and domesticated mammals, reptiles, amphibians, and fish. The biphasic developmental cycle (Fig. 1) begins with the elementary bodies (EBs), the extracellular and infectious form of the bacteria (size of 0.2 µm), infecting host mucosal epithelial cells. After attachment to and entry into the host cell, EBs develop within a membrane-bound endocytic vacuole called an inclusion and inside, differentiate into larger (0.8 µm), metabolically active but noninfectious reticulate bodies (RBs). After multiple rounds of division, RBs redifferentiate into infectious EBs, and mature EBs complete the developmental cycle, exiting the host cell via lysis or extrusion of the inclusion. 2 Under adverse environmental conditions, developing chlamydiae may enter a state referred to as persistence, more recently renamed the chlamydial stress response or the aberrant RB phenotype. 12 Stressed chlamydiae (termed aberrant bodies [ABs]) remain viable, but do not develop into EBs and are noninfectious. In vivo, the AB phenotype may contribute to prolonged, chronic inflammation, fibrosis, and scarring through continuing stimulation of the host immune system, although this concept is still highly controversial. 87 Due to their intracellular lifestyle, chlamydiae are dependent on host metabolites. This has considerable implications for diagnostics as these bacteria cannot simply grow on agar plates, requiring cell culture for their isolation. Isolation procedures require fresh tissue or swab samples immediately transferred into special transport or preservation media and proper storage at –80°C, conditions not routinely used or available in pathology laboratories. Formalin fixation and paraffin embedding (FFPE) of tissues or autolytic changes of organs impede any further isolation of chlamydial organisms.
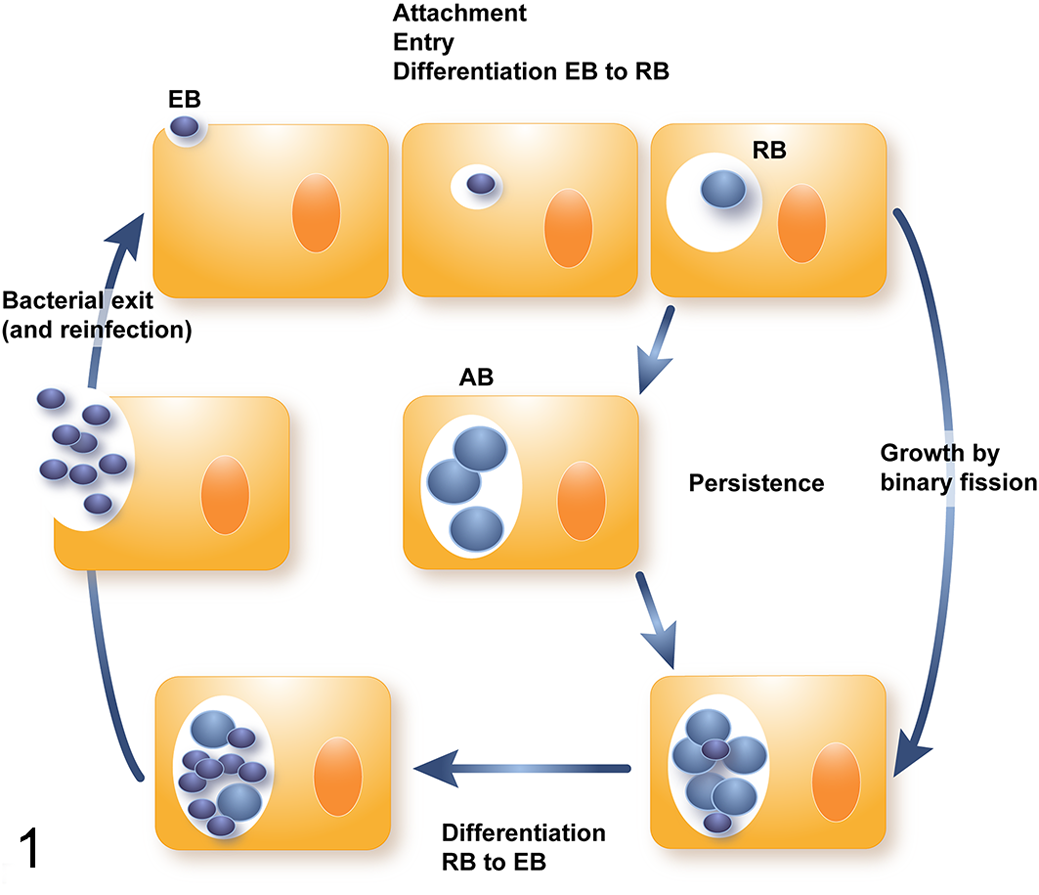
Chlamydia developmental cycle. The elementary body (EB) attaches to a host cell and differentiates into a reticulate body (RB) after entry, enclosed in a membrane-bound vacuole. The RB grows by binary fission and later differentiates into EBs, which are released and infect a new host cell. The persistence state is characterized by the formation of the aberrant body (AB) induced by stressful conditions.
Until recently, members of the Chlamydiaceae family were considered to be important pathogens of humans and animals with a well-defined taxonomic diversity and host range. This paradigm has been challenged with the help of molecular techniques revealing (a) a significantly expanded taxonomic diversity encompassing at least 8 other families of bacteria, colloquially referred to as Chlamydia-related bacteria (CRB) or Chlamydia-like organisms (CLO), within the order Chlamydiales; and (b) an expanded host range for certain chlamydial species, for example, C. psittaci and C. pneumoniae, that has posed new questions about the role of cross-host transmission in the epidemiology of these pathogens. Furthermore, these studies of new hosts have not only uncovered expanded host ranges for existing species but also identified new chlamydial species, for example, in reptilian and avian hosts. 170 Together, the animal host range of bacteria in the Chlamydiales order is significantly broader than thought before with over 400 host species documented globally, the majority of these being wild animals. 27 Infections of the latter including birds, reptiles, amphibians, fish and eutherians focusing on wild ungulates and marsupials have been recently reviewed 27 and are only discussed in this review in the view of related histomorphological findings.
Chlamydiosis in animals can range from asymptomatic infection to severe diseases with life-threatening illnesses depending on the host species affected and chlamydial species involved. Disease manifestations in chlamydial infections include conjunctivitis ranging from mild signs to corneal opacity, rhinitis, pneumonia, mastitis, arthritis/polyarthritis, pericarditis, polyserositis, encephalomyelitis, placentitis leading to abortion, stillbirth or weak neonates, endometritis/metritis, orchitis/epididymitis/urethritis, infertility, enteritis, and more. Still they might not be encountered by the pathologist because the lesions are too mild to cause death of the animal or chlamydiae are not considered as differential diagnosis due to the presence of other more common and well-known pathogens. Therefore, chlamydial infections, with the exception of chlamydial abortion in ruminants and avian chlamydiosis (also known as psittacosis/ornithosis) may present a diagnostic challenge for pathologists. Given the diversity of potential host species, chlamydial species and range of clinical signs, this review will try to summarize the most important aspects in the Chlamydia field tailored to the veterinary pathologist but also to any other interested reader. Human chlamydial infections due to C. trachomatis and C. pneumoniae as well as C. muridarum, the chlamydial species isolated from laboratory mice and hamsters and primarily used as infection inoculum in mouse studies as models for female genital infections, will be excluded from this review. Recent investigations in Peromyscus spp rodents from sylvatic and laboratory sources indicated that chlamydial infection is enzootic in this mouse species and that C. muridarum, or a closely related species or strain, is likely the agent in the tested rodent species. 126 Atypical Chlamydiaceae closely related to C. muridarum have been also detected by PCR in roe deer (Capreolus capreolus) in south-western France. 1 Moreover, C. muridarum was identified in oral and cloacal swabs of chickens, ducks and geese in China. 49
The CBR or CLO share phenotypic and genetic similarities with the Chlamydiaceae but are phylogenetically separated. Recently, 20 years of research on the CLO, since the detection of its first member Simkania, has been reviewed in detail. 173 Therefore, we will only elucidate the most important pathomorphological findings related to CLO-infections in the present review.
It is far beyond the scope of this review to cover all aspects and developments of the recent years. Therefore, we have limited our review to the past 5 years summarizing the most important developments in the field of animal chlamydial research.
Chlamydial Nomenclature
The chlamydial nomenclature has been under constant change in recent years. Besides the addition of new families, it was also proposed to split the genus Chlamydia into 2 genera, Chlamydia and Chlamydophila, based on analysis of the 16 S and 23 S rRNA genes. 45 This change was controversial. After significant debate within the chlamydial research community, a reversion to a single genus, Chlamydia, was proposed and is now accepted. 13,141 To date, the order Chlamydiales consists of 9 families: Chlamydiaceae, Clavichlamydiaceae, Cribchlamydiaceae, Parachlamydiaceae, Parilichlamydiaceae, Piscichlamydiaceae, Rhabdochlamydiaceae, Simkaniaceae, and Waddliaceae. 173 The family Chlamydiaceae contains a single genus, Chlamydia, including 11 species: 141 C. abortus, C. avium, C. caviae, C. felis, C. gallinacea, C. muridarum, C. pecorum, C. pneumoniae, C. psittaci, C. suis, and C. trachomatis as well as well as the 3 Candidatus (Ca.) species: Ca. C. ibidis, 176 Ca. C. sanzinia, 168 and Ca. C. corallus. 172 The Chlamydiaceae cause several diseases in humans (Table 1) and animals (Table 2), some of which are zoonotic, and this also refers to the CRB (Table 3). In humans, different serovars of C. trachomatis cause eye and urogenital infections responsible as the leading cause of infectious blindness worldwide (blinding trachoma) 166,167 and the most common bacterial sexually transmitted infection (STI). 111,113,157 Respiratory infections in humans are caused by C. pneumoniae, a chlamydial species also infecting horses, frog, reptiles and marsupials. 137 Zoonotic infections due to C. psittaci, a common pathogen of birds, have the best known zoonotic potential of the human pathogenic chlamydiae and cause relatively rare respiratory infections in birds but are associated with severe clinical manifestations in humans. 53,81,165 Infection with C. abortus in pregnant women after contact with aborting/lambing sheep and goats may also lead to abortion and, if untreated, to life-threatening illnesses. 44 Several other animal pathogenic chlamydial species, including C. felis, C. caviae, and C. suis, are known, or suspected, to cause infrequent human infections with various clinical presentations.
Disease in Humans Caused by Chlamydiae.

Diseases in Animals Caused by Bacteria Classified and Proposed to be in the Genus Chlamydia.

Diseases Caused by Chlamydia-Related Bacteria (CRB).

Historical Background
Trachoma, one of the diseases with a chlamydial etiology causing blindness in humans and one of the earliest known ocular diseases, has been identified in Egypt as early as 15
Remarkably, in 1909 an Austrian veterinarian reported smears from cows with transmissible vaginitis and healthy ones from several farms containing comparable forms to the so called “trachoma-bodies” and named them “Chlamydozoa” (Fig. 3). 16 Although these findings were well documented by a conclusive figure, the report remained unnoticed.
Psittacosis most probably originated in South America where psittacine birds were kept as pets, although alternative hypotheses have been proposed that Australian native parrots may have been the original source. 24 Fra Bartolomeo in 1615 described diseases in human individuals having had contact with psittacine birds. 122 In the second half of the 19th century, Jacob Ritter, a Swiss physician described an epidemic of psittacosis in his household causing 3 fatalities from 7 affected patients. 131 In his report, Ritter suspected that pet parrots and finches recently introduced into the house were the source of the infection and, accordingly, determined the incubation period while reporting the nontransmissibility of the disease from human to human. 122 The main clinical finding Ritter reported was an atypical pneumonia, called pneumotyphus at that time. In 1895, the term psittacosis derived from the Greek for parrot was first applied for this disease. 108 Later in 1893, Nocard had isolated a gram-negative bacterium from parrots dying of psittacosis (Bacillus psittacosis). 85 This particular organism subsequently was not regularly isolated from human or avian patients and later turned out to belong to the genus, Salmonella. 5,117 The inconsistent bacteriological isolations prompted the suspicion of a viral etiology of psittacosis especially during the pandemic of 1929–1930. 122 In 1930, independently but almost simultaneously, Levinthal, 89 Coles, 33 and Lillie 92 reported small, filterable bodies in infectious material and named them “Levinthal-Coles-Lillie (L.C.L.) bodies.” 102 Samuel P. Bedson first reported the biphasic developmental cycle of the psittacosis agent in tissues from experimentally inoculated mice. 14 In 1935, Burnet and Rountree 28 propagated the infectious bodies successfully on the chorioallantoic membrane of embryonated chicken eggs.
Until Pagès proposal of a taxonomy for Chlamydiaceae in 1966, 114 there were at least 7 attempts to define and name what we now know as chlamydiae. In 1958, the International Code of Nomenclature of Bacteria and Viruses was adopted and the genus Chlamydia replaced Miyagawanella, proposed by Brumpt in 1938. 25 The genus name “Chlamydia”, proposed by Jones and colleagues 77 had time precedence over Bedsonia 101 and Rakeia. 88 In 1971, Storz and Page proposed the order Chlamydiales, which contained only one genus, Chlamydia. In that genus, Chlamydia trachomatis and Chlamydia psittaci were distinguished as separate species 115 on the basis of sulfonamide susceptibility and the demonstration of glycogen accumulation within the inclusion by iodine staining.
Diagnosis of Chlamydial Infections—A Pathologist’s Point of View
The very first descriptions of human and animal diseases involving chlamydiae were based on anamnesis, clinical symptoms, epidemiology and presenting pathology. 16,131 Chlamydial infections can have an acute or chronic and even subclinical course of disease depending on the host species infected and the chlamydial species involved. The wide range of diseases and hosts makes a diagnosis difficult and diagnostic approaches cannot be standardized. Other than ovine and avian chlamydiosis, chlamydiae are often considered pathogens of little significance and, accordingly, detailed diagnostic investigations are not performed. Moreover, macroscopic and/or histologic changes might not be pathognomonic or indicative for a chlamydial infection or they are even not present at all. Chronic chlamydial infections in animals, particularly ruminants and pigs, are subclinical and ubiquitous in nature 130 and do often not induce any typical morphological lesions. Due to the variety of clinical pictures and animal hosts, and since Chlamydiaceae are often diagnosed in combination with other infectious agents, a definitive diagnosis typically requires laboratory investigations in most cases.
To detect chlamydiae in smears, several staining methods can be used, for example, modified Machiavello, modified Gimenez, Giemsa, Castaneda, or modified Ziehl-Neelsen (MZN) stain (reviewed in Sachse et al). 146 Intracellular chlamydial inclusions can be visualized by Giemsa staining on tissue sections, but methylene blue staining was demonstrated to be more reliable compared to Giemsa when detecting C. abortus in fetal membranes. 36 However, these techniques are nonspecific and can cross-react with other bacterial species; care, therefore, must be taken with interpretation of results. During the 1960s, ultrastructural investigations confirmed the bacterial nature of chlamydiae by transmission electron microscopy (TEM) but, today, is primarily used for research purposes. 123
Nowadays, antigen detection in pathology laboratories is generally performed on formalin-fixed and paraffin-embedded (FFPE) tissue samples using different histochemical and immunohistochemical techniques. 146 Immunohistochemical methods using monoclonal antibodies directed against chlamydial surface antigens, such as LPS or MOMP, are more sensitive compared to histochemical staining 146 and routinely used for diagnostic purposes. 21 Various other approaches have been developed such as enzyme-linked immunosorbent assays (ELISAs) and fluorescent antibody tests (FATs). Although these methods successfully demonstrate the presence of chlamydial organisms in the diagnostic specimen, they generally do not allow the identification of the involved chlamydial species and/or serotype/genotype because they use either nonspecific cytochemical stains or monoclonal antibodies based on Chlamydiaceae-specific antigens, such as the chlamydial LPS. 146
The role of histopathology during the diagnostic process should not be underestimated, as it is important to ensure that the infectious agent identified is related to the lesion present and that it is not a mere contaminant, possibly originating from the environment. 146 This is especially important since nucleic acid amplification tests are considered the gold standard to diagnose animal and human chlamydial infections and novel culture-independent metagenomic approaches enable identification of novel chlamydial species, at least at the Candidatus level. However, the significance of detecting chlamydial DNA in specimens is questionable as a sole diagnostic approach, in the absence of other methods or without related pathological findings. Demonstration of (1) the presence of viable, replicating bacteria and (2) etiological importance are mandatory to draw any conclusion on whether chlamydiae are involved in the disease process or not.
Avian Chlamydiosis (Chlamydia psittaci, Chlamydia avium, Chlamydia gallinacea)
Traditionally, avian chlamydiosis, also named psittacosis or ornithosis, was due to infection with C. psittaci (reviewed in Knittler and Sachse, 81 Sachse et al, 145 and Szymańska-Czerwińska and Niemczuk 165 ). The latest findings related to C. psittaci have been extensively reviewed 81,125,145 and therefore are discussed only briefly.
Chlamydia psittaci is known to occur in more than 465 avian species, including domestic, companion and wild birds. The highest percentage of infection is found in pigeons and psittacine birds. The infection can lead to systemic and occasionally fatal disease in birds, but especially older psittacine birds and poultry may not show clinical signs but shed the organisms for extended periods of time. Necropsy of affected birds often reveals multifocal hepatic necrosis, splenomegaly and fibrinous airsacculitis, pericarditis, and peritonitis. In a recent 5-year retrospective study of avian diseases at the University of Georgia including 153 captive bird species, bacterial infection was the most frequently diagnosed cause of death with gram-negative bacteria and C. psittaci most commonly involved. 110 The highest percentage of bacterial infections was reported in Psittaciformes, followed by Passeriformes, Galliformes, Columbiformes, and Anseriformes.
Zoonotic infections due to C. psittaci vary from inapparent to severe systemic disease with pneumonia, myocarditis and encephalitis (reviewed in Radomski et al 125 and Sachse et al 145 ). Human psittacosis cases are most common in bird keepers, poultry workers, veterinarians, and health care workers. Human-to-human transmission can occur but is exceptionally rare apart from a recent reported outbreak in Sweden. 179
Atypical Chlamydiaceae species in European and Asian chicken flocks 49,191 and urban pigeons 142 were described in recent years and resulted in the proposal of 2 new avian chlamydial species and one Candidatus species in the genus Chlamydia: Chlamydia gallinacea and Chlamydia avium 143,144 and Candidatus C. ibidis. 176 C. gallinacea and C. avium were detected alongside C. psittaci in the same flock and even in the same bird.
C. gallinacea has been detected in domestic poultry, guinea fowl, turkeys, and ducks, which did not shown signs of disease. It is thought that C. gallinacea is an endemic organism in chickens with the ability to persist over time. 49 Experimentally infected chickens remained without overt clinical disease but showed significant body weight reduction. 49 The type strain of C. gallinacea has been recently sequenced and harbors a plasmid, a suspected virulence factor in some but not all chlamydial species. 59 C. gallinacea has been reported in chickens in different European countries, Argentina and China. 47,49,64 Recently, C. gallinacea was detected in wild birds (woodcocks, Scolopax rusticola) from South Korea, but based on phylogenetic analyses of polymorphic ompA sequences, the Korean C. gallinacea was a genetic variant different from European and Chinese isolates. 74 The zoonotic potential of C. gallinacea has been suggested after an outbreak of atypical pneumonia in French poultry slaughterhouse workers. 84 Interestingly, a recent study in China 90 detected DNA of C. psittaci and C. gallinacea from whole-blood samples, milk, feces and vaginal swabs of clinically healthy dairy and beef cattle. The serovars found were identical with those of poultry in the same region suggesting bird to ruminant transmission. As it stands, C. gallinacea should be considered an avian pathogen with the potential to infect humans and other animals. The potential macroscopic or histomorphological lesions remain to be elucidated.
C. avium has been found in pigeons and psittacines in European countries with limited data linking it to respiratory disease in parrots and pigeons. 143 The zoonotic potential of C. avium is unknown. Initial isolates of C. avium were obtained from cloacal swabs, feces or internal organs from psittacine birds and pigeons which were asymptomatic or had respiratory signs and/or diarrhea. 144
Candidatus Chlamydia ibidis was isolated from cloacal swabs of a feral African sacred ibis (Threskiornis aethiopicus). 176 The isolate was obtained from a healthy ibis shot during an official culling operation carried out in Western France in 2009 and 2010. To date, the virulence characteristics of Ca. C. ibidis, its geographical origin, its infectivity and pathogenicity for poultry and bird wildlife or other animals and its zoonotic potential is completely unknown.
The complexity of chlamydial infections in birds is further manifested by a recent report of polymerase chain reaction (PCR)–based avian C. abortus strain detection in Polish wildfowl. 164 To date, the pathogenicity of these new avian C. abortus strains remains unknown and needs further investigation. Moreover, a new species, intermediary between C. psittaci and C. abortus, from a red-tailed hawk was retrieved by whole-genome sequencing. 78 It has been hypothesized that birds-of-prey or other scavengers and predators might acquire multiple chlamydial species in their intestinal tracts resulting in emergence of novel species by recombination events.
Chlamydia psittaci in Horses
Few reports of chlamydial infections in horses are available in the literature associated with equine pneumonia or abortion after infection with C. abortus or C. pneumoniae. 100,175 Chlamydial abortion in mares due to C. psittaci is a known cause of equine abortion 19,55,163 but has recently gained new attention when an avian-like C. psittaci strain was found in equine placentitis cases associated with subsequent zoonotic human psittacosis in New South Wales, Australia. 31,71,120 In 2014, this outbreak caused 5 cases of human psittacosis in veterinary students and staff exposed to equine fetal membranes. A mild, diffuse, interstitial placentitis was present in a mare who delivered a live but ill foal that died suddenly one week later. Infection with C. psittaci was diagnosed by PCR and multilocus sequence typing showed assignment to the highly virulent C. psittaci 6BC lineage, identical to sequences recently described in Australian parrots and humans. 24 The authors 71 postulated transmission via indirect contact, presumably via fecal environmental contamination from C. psittaci-infected parrots as the infection source. These findings highlight the risk of zoonotic transmission originating from C. psittaci-infected equine placental membranes and depending on the geographical localization, indicate the need to consider C. psittaci as differential diagnosis in equine abortion.
Chlamydia abortus in Ruminants and Other Hosts
Small ruminants can be infected with C. abortus, C. pecorum, C. psittaci, and more rarely C. suis. C. abortus causes late-term abortions, stillbirths and birth of weak neonates which fail to survive beyond 48 hours after birth in sheep and goats and to a lesser extent in cattle, pigs, horses and wild ruminants (reviewed in Longbottom and Coulter 95 ). Moreover, C. abortus was also suggested to play a role in yak abortion in China after its isolation from 9 aborted fetuses. 91 Ovine chlamydiosis due to C. abortus, also known as enzootic abortion of ewes or ovine enzootic abortion (OEA), is the most common infectious abortigenic agent worldwide, causing significant economic losses. 96 Chlamydiosis of small ruminants due to C. abortus is a zoonosis with pregnant women being particularly at risk (reviewed in Essig and Longbottom 44 ). Recently, a case of an atypical pneumonia due to C. abortus in a laboratory worker involved in intranasal experimental C. abortus infections in sheep was reported. 112
C. abortus strains within Europe, in contrast to C. suis strains, have a limited diversity but strong geographical signatures. They do not harbor a plasmid and recombination was not identified in a recent study. 151
The clinical history of abortion in sheep and goats, often in late pregnancy (ie, the last 2–3 weeks), precedes the investigation of a macroscopically changed placenta (if available) with edema, hemorrhage, necrosis, and purulent exudate (Fig. 4). The histopathology shows a purulent to necrotizing placentitis with vasculitis and, if well preserved, intratrophoblastic basophilic chlamydial inclusions might be observed (Fig. 5) (reviewed in Borel et al 21 and Sachse et al 146 ). If a large number of organisms are present in infected placental specimens, the disease can be diagnosed by stained smears, antigen tests, immunofluorescence, or immunohistochemistry (IHC) labeling or by molecular methods such as PCR. In the majority of abortion cases (71.6%) where OEA was present in a flock, other abortigenic agents such as Toxoplasma, Campylobacter, Salmonella and Listeria spp. were also diagnosed, 96 highlighting the importance of considering coinfections and mixed infections during abortion investigations. Interestingly, while a live temperature-sensitive vaccine strain 1B of C. abortus has been available to control infections, it was reported that this vaccine strain can lead to abortions as shown in Europe 184 and recently in New Zealand, 148 causing lesions indistinguishable from naturally induced C. abortus cases.
In sheep, chlamydial placentitis develops late in gestation and from then on gradually progresses to a diffuse inflammatory response, with thrombotic vasculitis and tissue necrosis (reviewed in Sammin et al 147 ). The pathogenesis of abortion might be the result of a combination of impairment of materno-fetal nutrient and gaseous exchange, disruption of hormonal regulation of pregnancy, and induced cytokine aggression. 44 Fetal pathology can occur after thrombo-embolic infection of the fetus and results in focal necrosis in liver, lung, spleen, and less frequently in lymph nodes and brain. 29 During experimental infection, ewes were susceptible to infection from early gestation onward and infections of the placenta and fetuses occurred from about 60 days of gestation, but pathological changes were not detected until after 90 days of gestation. 29 This included rapid replication of C. abortus in the trophoblasts at the limbus of the placentomal hilus and continuous spread of infection to the intercotyledonary and cotyledonary placenta.
In nonpregnant ewes, relatively low doses of infectious organisms (EBs) can induce chlamydial latency as shown in intranasal infection studies. 97 During latency, the infection remains inapparent in the nonpregnant animals and only becomes evident during a subsequent pregnancy. When the animals become pregnant, the infection proliferates, and initiates placental infection and abortion. 44 The route of primary infection is presumed to be oronasal following ingestion of organisms and establishing an initial infection in the tonsil from where it may disseminate by blood or lymph to other organs (reviewed in Essig and Longbottom 44 ). Chlamydial latency was recently investigated experimentally in sheep using 3 different infection doses of C. abortus. 97 The high dose stimulated a protective immunity resulting in a much lower abortion rate whereas the medium and low doses resulted in a latent infection with infection of the placenta and abortion in the subsequent pregnancy, the key feature of latency. 97 In this study, the most consistent histological lesions were found in the brains of aborted lambs, lambs born dead and those that died soon after birth, manifesting as focal leucoencephalomalacia mostly located in the cerebral white matter and being indicative of anoxic damage. Histological lesions in the fetal liver were composed of suppurative periportal hepatitis and focal necrosis. Lesions in the fetal lung consisted of neutrophil infiltration in alveoli, suppurative interstitial inflammation and interlobular edema and hemorrhage. The latter experimental study by Longbottom et al 97 was performed using the C. abortus strain S26/3, isolated from the placenta of a sheep that aborted due to OEA in Scotland, while the same authors 93 used strains LLG and POS, which have been isolated in Greece from aborted goat and sheep fetuses and which are genetically different to S26/3, in a recent comparative study. In doing so, they showed that the placental pathology, infectious load and serological response was clearly less pronounced for the strains LLG and POS than for the C. abortus strain S26/3.
C. abortus has also been associated with epididymitis, pneumonia, arthritis and conjunctivitis in ruminants and has been isolated from feces of healthy sheep and goats (reviewed in Rodolakis and Laroucau 133 ). In a recent study, C. abortus was detected in the oviduct of 25% of cows by PCR but a firm link with pathology could not be established as a higher percentage of PCR-positive oviducts were collected from macroscopically normal reproductive tracts (37.5%), than from those with lesions (18.1%). 4 Histological lesions in cows with PCR-positive oviducts comprised ovarian cysts, mucometra, ovarian adhesions, fibroproliferative lesions of the oviducts, and uterine abscess.
C. abortus has been recently detected by PCR and sequencing in cases of ovine, caprine and bovine infectious keratoconjunctivitis in India including mixed infections with C. psittaci and Mycoplasma spp. 50
Chlamydia pecorum in Ruminants
Chlamydia pecorum can infect a wide host range that includes production animals such as sheep, goats, pigs and cattle, but also wild ruminants and wild boars, as well as the koala. The infection is thought to be endemic in cattle and sheep and the majority of infections are reported as clinically inapparent, mostly localized in the gastrointestinal tract. However, the significance and economic impact of these subclinical infections in livestock remains uncertain. 90,130 More recently, chlamydial infections due to C. pecorum in sheep and cattle have been associated with important clinical manifestations such as infectious arthritis, conjunctivitis, encephalomyelitis, infertility, enteritis, reduced growth rates, mastitis, and pneumonia as reviewed in detail. 177 Recent and new knowledge has been mostly accumulated from studies in sheep and cattle from Australia and New Zealand. Initial reports date back to 1940 and 1960 and described encephalomyelitis cases in cattle and polyarthritis in feeder lambs in the USA. 177 Polyarthritis and keratoconjunctivitis due to C. pecorum infections are commonly seen in young lambs and calves. Arthritis is characterized by an inflammatory and proliferative response in synovial membranes possibly progressing to chronic changes with articular erosions and fibrotic thickening. These changes can result in weight loss of the animal, animal welfare issues and economic losses for the farmer. Ovine arthritis cases in Australian weaned lambs showing joint stiffness and depression were described recently. 178 The majority of animals also suffered from concurrent conjunctivitis. Histopathology in 2 lambs revealed chronic, multifocal lymphoplasmacytic synovitis and mild, multifocal hyperplasia of synoviocytes.
C. pecorum-induced keratoconjunctivitis in ruminants can lead to prominent conjunctival follicular formation and corneal neovascularization. 177 However, other studies 119 could not find an association between the presence of chlamydial DNA in sheep conjunctival samples and the onset of clinical ocular disease. These authors 119 also detected C. abortus, C. suis, and CLO in conjunctival swab samples by PCR and sequencing.
Infection with C. pecorum in young cattle can cause fatal sporadic encephalomyelitis with encephalomyelitis, systemic infection and fibrinous polyserositis including pericarditis, pleuritis, peritonitis and arthritis. Brain lesions consist of a severe meningoencephalitis with vasculitis, thrombosis and malacia most severe in the brainstem and cerebellum. 72 Histological brain lesions might overlap with those of Histophilus somni. 65 By IHC, chlamydial antigen can be demonstrated in the cytoplasm of the endothelium and macrophages of brain, spleen and lung and in the mesothelium. 72 Typing of C. pecorum strains revealed differences between strains detected in brain, heart, lung, liver and the intestine, suggesting that different strains may have different pathogenic potential, an observation that was previously also suggested from studies of sheep. 73
Associations between C. pecorum infection and enteritis or reproductive diseases such as placentitis and abortion, vaginitis, endometritis, mastitis, and orchitis have been reported with shedding of the organism through the fecal or vaginal route. C. pecorum has been also implicated in cases of pneumonia in sheep and cattle, inducing a fibrinous and broncho-interstitial pneumonia in older reports (reviewed in Walker et al 177 ). A recent report identified C. pecorum in 8 out of 98 cases of bovine pneumonia in Scotland, while other common respiratory pathogens such as BHV-1, BRSV, and PI-3 were excluded. 189 However, the pathologic findings in the lungs were not further specified in this case report. Experimental chronic but subclinical C. pecorum infections in calves led to follicular bronchiolitis with marked BALT hyperplasia in the lungs (reviewed in Reinhold et al 130 ).
C. pecorum is abortigenic in small ruminants although to a much lesser extent than C. abortus. Histological lesions in goat abortions consisted of a suppurative and necrotizing placentitis with vasculitis and thrombosis, lymphohistiocytic and neutrophilic hepatitis and fibrinosuppurative enteritis in fetal liver and intestine. 48 Lesions in the placenta and fetal liver were similar to C. abortus-induced abortion whereas the intestinal lesions have not been described before and might represent a unique pathological feature for infection with C. pecorum.
Moreover, subclinical C. pecorum infections might have a significant impact on herd performance leading to reduced growth rates in calves. 124 Asymptomatic endemic C. pecorum infections reduced growth rates in calves by up to 48%, indicating a potential for C. pecorum-induced growth depression due to chronic low-level systemic inflammation. This study 124 also confirmed that infection takes places early in life and repeatedly by multiple strains circulating ubiquitously in herds with mixed infections or simultaneous infection with 2 or more strains at different mucosal sites.
In summary, clinical disease due to C. pecorum infection might be triggered by other factors such as stress and coinfections from the host side and on the pathogen side, depending on the virulence of the strain resulting in a multifactorial pathogenesis.
Chlamydia pecorum in Koalas
Chlamydial infections in the koala (Phascolarctos cinereus), the iconic Australian marsupial, have been associated with keratoconjunctivitis (Fig. 6), rhinitis/pneumonia, and urogenital tract disease with C. pecorum and C. pneumoniae as involved chlamydial species (reviewed by Polkinghorne et al). 121 Chlamydiosis in the koala has a prevalence of up to 100% in some populations in the eastern states of Australia. 121 C. pecorum, initially named C. psittaci, is more virulent in the koala than C. pneumoniae and infection can lead to serious complications such as blindness after ocular infection and infertility after genital infection. 66 The urogenital infection is sexually transmitted but also mothers are thought to infect their joeys through close contact. The inflammation of the urinary tract is called “wet bottom” or “dirty tail” due to brown urine staining and wetness of the fur in this region caused by cystitis with subsequent incontinence and urine drippling. Chlamydial infection in the koala can be considered in 3 categories: subclinical infection (no signs of disease), overt disease (presence of obvious external clinical signs such as conjunctivitis and “wet tail”) or inapparent overt disease (lesions are not clinically obvious but can be only detected using ultrasound, postmortem examination or histopathology). 180 Animals with active clinical disease shed the highest level of C. pecorum organisms/DNA and might therefore be the most important source of transmission. 121 For many years, anecdotal evidence suggested that chlamydial disease was primarily limited to koalas in northern populations. 121 A more recent study has revealed that chlamydiosis is indeed evident in South Australian koala populations, although at a much lower level than in their northern counterparts. 156
Histologically, inflammatory infiltrates consist of lymphocytes and plasma cells including follicle formation and to a lesser extent, macrophages and neutrophils and accompanying fibrosis. Such inflammatory processes mainly involve the eye (conjunctivitis with conjunctival hyperplasia and fibrosis, corneal edema and opacity due to ulceration, vascularization and pigmentation) and the female urogenital tract (uterus, uterine tubes, cervix, ovarian bursa, vagina, urinary bladder) resulting in endometritis, metritis, salpingitis, vaginitis and cystitis. 54 Cystic lesions of the ovarian bursa are considered a sequela of previous inflammation and fibrosis with concomitant hydrosalpinx. However, other organs (kidney, prostate, cloaca, lung) can be also affected resulting in chronic interstitial, pyogranulomatous nephritis or pyelonephritis, prostatitis, proctitis, and interstitial pneumonia. 57 A case of pyogranulomatous bronchopneumonia, proliferation of the bronchiolar and alveolar epithelium and interstitial fibrosis with chlamydial inclusions in bronchiolar epithelium was recently reported in a juvenile male koala that died after an episode of respiratory disease. 99 Chlamydial organisms were demonstrated by IHC and TEM and C. pecorum DNA was detected by PCR.
In general, chlamydial antigen can otherwise be typically detected by IHC in epithelial cells and macrophages including alveolar, hepatic and splenic macrophages of affected organs. 26,54,57
While C. pecorum-induced urethritis and prostatitis in the male koala is well known, chronic-active granulomatous orchitis and epididymitis with interstitial fibrosis and resulting aspermia has been described recently in koalas with history of cystitis. 75 Chlamydial inclusions were demonstrated in Sertoli cells of the testis by IHC and TEM and chlamydial DNA was detected by PCR.
Chlamydia suis
Chlamydial species known to infect pigs are C. suis, C. abortus, C. pecorum, and C. psittaci. Such infections have been associated with a wider range of lesions such as conjunctivitis, pneumonia, pericarditis, polyarthritis, polyserositis, pseudo-membranous or necrotizing enteritis, periparturient dysgalactiae syndrome, vaginal discharge, return to estrus, abortion, delivery of weak piglets, increased perinatal and neonatal mortality and inferior semen quality, orchitis, epididymitis, and urethritis in boars (reviewed in Schautteet and Vanrompay 150 ). Before 1999, C. suis strains were referred to as C. trachomatis, and the type strain of C. suis, S45, was isolated in Europe in the late 1960s from feces of an asymptomatic pig in Austria by Kölbl. 150 Infection with C. suis is mostly associated with conjunctivitis, respiratory infections, reproductive disorders and enteritis. C. suis caused enteritis when inoculated in gnotobiotic piglets 51,134 or induced histological intestinal lesions in asymptomatic young weanling pigs. 135 However, under field conditions, chlamydiae are commonly found in the intestine of pigs and are mostly associated with endemic subclinical infections. 58 Recent studies mostly focused on the tetracycline-resistant C. suis strains which have been initially detected in the USA 86 and later in Italy, Switzerland, Belgium, Israel and Cyprus (reviewed in Borel et al 22 ). C. suis is the only chlamydial species known to have naturally acquired genes encoding for tetracycline resistance and it has been shown recently, that selective pressure appears to further promote resistance in fattening pig herds. 181 By PCR and microarray, C. suis DNA has been detected in conjunctival swabs of Nepalese trachoma patients 37 and Belgian slaughterhouse workers. 39,40 Recently, isolation of C. suis from nasal, pharyngeal and stool samples taken from Belgian farmers was reported, demonstrating the presence of live organisms. 38 These findings indicate the potential zoonotic transmission of C. suis from pigs to humans. Taken together, infection with C. suis might be opportunistic in nature and ocular, respiratory and intestinal virulence variation of C. suis infection might be influenced by the age and immune status of the host, possible coinfections and genetic variations of C. suis strains including high recombination rates. 154
Chlamydia felis and Chlamydia caviae
C. felis and C. caviae are agents of infectious conjunctivitis in cats and guinea pigs, respectively, but their DNA has also been detected in swab samples of dogs (C. felis, C. caviae), horses (C. caviae) and rabbits (C. caviae). 116
C. felis was initially named Bedsonia sp. (psittacosis/lymphogranuloma-venereum trachoma group), the agent of feline pneumonitis causing conjunctivitis, rhinitis and pneumonitis in cats. 6,7 Later, it was grouped into C. psittaci and finally named C. felis. 141 Although the original isolate was derived from the lung of a cat with pneumonia, 6 conjunctivitis is now known to be the most common clinical sign of infection and the term “feline pneumonitis agent” seems misleading. 11 C. felis is endemic among pet cats worldwide with reports of zoonotic infection. 149,161 The first report of a zoonotic infection presented as an acute follicular keratoconjunctivitis in a cat owner, whose cat had a recent history of rhinitis and conjunctivitis. 149 In cats, C. felis primarily targets the conjunctival epithelium and causes acute to chronic or recurrent conjunctivitis, particularly in young cats (2 to 12 months of age). Infected cats have mucoid/mucopurulent to follicular conjunctivitis and rhinitis (reviewed in Sykes 162 ). Chlamydial inclusions can be demonstrated in Giemsa-stained smears prepared from conjunctival scrapings but PCR-based methods are more sensitive and specific. During the acute neutrophilic conjunctivitis phase, high levels of C. felis are excreted, peaking approximately 1–2 weeks postinfection. Thereafter, in the chronic conjunctivitis phase, lower levels of C. felis are excreted persisting up to 2 months postinfection prior to resolution of clinical signs and cessation of chlamydial shedding. Cats can also remain persistently infected for longer time periods and may maintain an asymptomatic carrier status. C. felis infections have been also linked to reproductive tract disease and lameness but etiological evidence remains weak. 16
During experimental infection, signs of conjunctivitis appeared 5–10 days post infection and lasted for 22–45 days. In some cats, mild rhinitis but no clinical signs of lower respiratory tract disease were present. By histology, small foci of broncho-interstitial pneumonia were detected in 3 out of 6 cats. Chlamydial inclusions were present from day 7 to day 14 in the cytoplasm of conjunctival smears. 61 Experimental infection of kittens not only induced conjunctivitis but also resulted in fever, lethargy, lameness and reduction of weight gain. 174 In addition, vaginal excretion of C. felis was observed in cats infected at an age of 4–6 months. Prolonged vaginal and rectal shedding in cats with conjunctivitis has been reported indicating that the genital and intestinal tracts might be a reservoir for persistent infection. 162 C. felis has also been found in the lung, spleen, liver, kidney, and peritoneum of cats, however, the significance of these findings has to be elucidated. 162
The species, C. caviae, was taxonomically separated from C. psittaci in 1999 along with several other species formerly considered to be biovars of the latter species. 45 C. caviae is a chlamydial pathogen that has been exclusively isolated from guinea pigs namely from the conjunctiva (GPIC, guinea pig inclusion conjunctivitis) 109 but is extensively used as an animal model of human C. trachomatis ocular 128 and genital infections. 127 C. caviae is one of the most common infectious causes of conjunctivitis in guinea pigs, but can also induce rhinitis, lower respiratory tract disease and abortion. Guinea pigs with conjunctivitis can be evaluated by preparing cytological smears from conjunctival swabs stained with Giemsa. 160 Intracytoplasmic chlamydial inclusion bodies can be observed in conjunctival epithelial cells, enabling the initial diagnosis of GPIC which can be confirmed by PCR demonstrating C. caviae as the involved chlamydial species. In a larger case study, 98 59 out of 123 guinea pigs were positive for C. caviae, 48 out of 59 presented ocular signs (ocular discharge, conjunctivitis, chemosis, keratitis), nasal discharge and other respiratory signs and vaginal discharge whereas 11 PCR-positive guinea pigs remained asymptomatic. PCR-positive symptomatic guinea pigs suffered most often from a neutrophilic conjunctivitis with visible chlamydial inclusion bodies in cytological smears. DNA of C. caviae was also detected in a rabbit (mild seromucous discharge and conjunctivitis), a cat (no clinical signs) and the owner (mild serous ocular discharge) of one guinea pig collection suggesting interspecies transmission and zoonotic potential.
During experimental ocular infection studies, conjunctival inflammation in infected guinea pigs is seen for about 30 to 40 days, chlamydial inclusions and PMNs are present for up to 21 days and mononuclear cells (lymphocytes, fewer plasma cells, and macrophages) are found on days 7 to 25 in conjunctival scrapings. 106 Sporadically, a mild, diffuse epithelial and subepithelial keratitis of the upper half of the cornea occurred. Repeated ocular infection with C. caviae resulted in severe conjunctival chronic inflammation lasting for months followed by pannus and follicle formation in the palpebral conjunctivae, scarring of the lower conjunctivae and deformities of the lower lid. The inflammation was dominated by mononuclear cells and rarely by small numbers of PMNs or chlamydial inclusions. 107 The pathological picture during repeated ocular C. caviae infection closely resembled trachoma disease in human patients, mediated most likely through a delayed-type hypersensitivity.
After vaginal experimental infection, C. caviae ascends to the upper genital tract leading to inflammation and fibrosis of the oviducts. This ascension results in tubal obstruction, hydrosalpinx and infertility as observed in C. trachomatis-infected women, 127 making this animal an extensively used model for studying the pathogenesis of female infertility.
Chlamydia pneumoniae
In humans, C. pneumoniae, an agent of respiratory infection, is nearly ubiquitous in humans, with seropositivity rates of 70–80% in older populations, suggesting most people experience infection during their lifetime. 32 In animals, infection with C. pneumoniae has been reported in horses, koalas and other Australian marsupials, frogs and reptiles. 136,137 Moreover, DNA of C. pneumoniae was recently detected in whole-blood samples and vaginal swabs of cows from China. 90 The horse strain, N16, was isolated from the respiratory tract of a horse and contains a plasmid, 158 however, proof of associated pathology is weak. Previous case studies linked the presence of C. pneumoniae with disease in free-ranging or captive frog species 15,17,129 but recent data are missing.
The initial description of C. pneumoniae infection in koalas was in the late 1990s by Wardrop. 182 Clinical signs reported in koalas included respiratory distress, sneezing, coughing, chest congestion, difficulty in breathing, rhinitis and nasal discharge. 182 However, C. pneumoniae infections in koalas are usually low grade compared to C. pecorum infections in this host. Unfortunately, detailed information on pathologic changes in the upper and lower respiratory tract due to C. pneumoniae is missing as recent studies mostly focused on genomic and in vitro phenotypic comparisons between the C. pneumoniae koala type strain LPCoLN and human strains of C. pneumoniae. 103 –105 On the other hand, comparisons of strain-specific differences between C. pneumoniae isolates from both human and animal origin might help to understand the zoonotic potential and evolutionary history of this pathogen. Such data showed that animal isolates were more diverse than the isolates of human origin with a number of genes in animal isolates observed to be intact while their human homologues were decayed. 103,104 This observation has led to the suggestion that animal strains are ancestral to human strains and that cross-host transmission to humans from animals may have occurred several times in the evolutionary history of this pathogen. 103,104,137
C. pneumoniae and unidentified Chlamydiales were detected by PCR in western barred bandicoots (Perameles bougainville) with ocular signs such as corneal opacity, conjunctivitis, ocular discharge, and blepharitis. 183 Further studies confirmed C. pneumoniae as cause of conjunctivitis, cataract and corneal scarring, as well as possible pneumonia in bandicoots. 83 The C. pneumoniae strain B21 isolated from a western barred bandicoot also harbors a plasmid like the koala LPCoLN strain and the horse N16 strain and is, as expected, most closely related to the koala strain. 136
Chlamydial Infections in Reptiles
Chlamydial infections have been reported in both free-ranging and captive reptilian hosts including puff adders, boas, chameleons, crocodiles, turtles and tortoises. Initial reports described C. psittaci as the causative agent of chlamydiosis in reptiles 62,132 but nowadays, C. pneumoniae is considered the most widespread. The infection manifests as granulomatous inflammation in inner organs such as heart, liver, spleen and lung. In a retrospective study of captive (private, zoo) reptiles (n = 90), 155 9 snakes and chelonians were positive for C. pneumoniae in granulomas of the lung, pericardium, heart, liver and kidney. Of these, 3 cases had mixed infections with mycobacteria. Other previous reports describe cases of histiocytic granulomas (puff adders), 68 granulomatous inflammation (flap-necked chameleon), 70 necrotizing enteritis (green iguanas), 20 necrotizing myocarditis (green turtles), 60 proliferative pneumonia (Burmese python and other snakes), 20,46 multiple granulomas and lympho-plasmacytic gastroenteritis in emerald tree boas, 67,69 and acute hepatitis and conjunctivitis in crocodiles. 62,63 An unusual case of C. pneumoniae infection was recently described in a royal python showing neurological signs. 34 Histology revealed lymphocytic enteritis with histiocytic granulomas, granulomatous splenitis and meningitis. Chlamydial IHC was positive in splenic granulomas and PCR identified C. pneumoniae in pooled visceral tissues.
More recently, it has become clear that chlamydiae might more often be present as opportunistic pathogens in reptiles. By PCR, C. pneumoniae was commonly detected in routinely sampled reptiles from Japanese zoos including snakes, turtles, lizards, and chameleons 79 and in cloacal swab samples of snakes from an Argentinian recreational area. 46 During a longitudinal case study in a collection of 58 snakes, 138 C. pneumoniae infections were not necessarily lethal for snakes but were common in clinically inconspicuous carriers.
A follow-up study 171 investigated and characterized C. pneumoniae and potential other novel chlamydial infections in the choana and cloaca samples of 137 clinically healthy captive snakes from 6 private collections resulting in the description of novel chlamydial genotypes. In summary, unique 16S genotypes were designated from 21 choanal or cloacal swab samples, with closest sequence identity to uncultured Chlamydia sequences, C. pneumoniae strains or a chlamydial sequence isolated from a tortoise, indicating the presence of potentially novel Chlamydia species. Such novel chlamydial species might be either present as asymptomatic choanal/cloacal infections or may be able to cause disease in coinfection with other pathogens (Fig. 7). Culture-independent full genome sequencing of genomic DNA obtained from the choana of a captive, clinically inapparent Madagascar tree boa (Sanzinia madagascariensis volontany) revealed the novel Candidatus species Chlamydia sanzinia, 168 which contains a plasmid and falls in a phylogenetic clade with C. pneumoniae and C. pecorum. As a follow-up to this investigation, the same methods were applied to a larger selection of chlamydial PCR-positive specimens from the collection previously screened by Taylor-Brown et al. 171 While this method was less successful than the initial study, the genome of another novel chlamydial species, Candidatus Chlamydia corallus, was retrieved from the choana of an asymptomatic captive Amazon Basin emerald tree boa (Corallus batesii). 172 Notably, while chlamydial chromosomal sequences could not be resolved for the remaining samples, plasmid sequences could be, suggesting additional and untapped Chlamydia genus diversity in the remainder of the samples. In summary, all these recent findings indicate that snakes may still harbor a significant and largely uncharacterized level of diverse and novel chlamydial species requiring further investigation.
CRB, CLO with Proven Pathogenic Potential
At least 13 new Candidatus species of CRB have been reported in reptilian, amphibian, avian, piscine and protozoan hosts in the last 4 years. 170 Of these, CRB seem to play a pathogenic role in ruminant abortion and human miscarriage (family Waddliaceae and Parachlamydiaceae) as well as in epitheliocystis in fish, the common gill disease (families Simkaniaceae, Ca. Parilichlamydiaceae, Ca. Piscichlamydiaceae, and Ca. Clavochlamydiaceae).
CRB-related diseases in ruminants have been recently reviewed in detail. 187 Waddlia chondrophila and Parachlamydia acanthamoebae have been demonstrated in placentas of women with adverse pregnancy outcomes such as miscarriage (reviewed in Ammerdorffer et al). 3
Waddlia chondrophila was initially isolated from a case of bovine abortion in the United States in 1986 and initially classified as Rickettsia. 42 However, pathological observations associated with this finding were not reported. A subsequent study from Germany detected W. chondrophila in a septic stillborn calf in a coinfection with Neospora caninum. 56 Pathological findings included lymphohistocytic meningoencephalitis with fibrinous exudate and a focal leukoencephalomalacia in the fetal brain, acute suppurative pneumonia and bronchiolitis and a multifocal suppurative necrotizing placentitis with vasculitis but it remained unclear if these lesions were caused by W. chondrophila, N. caninum, or both. In a Swiss study investigating 343 placental samples from late-term bovine abortion cases, 3 were positive by Waddlia-specific real-time PCR, and of these, 2 were confirmed by IHC. 18 All 3 cases showed a purulent to necrotizing placentitis and additional vasculitis as well as the presence of Parachlamydia in 2 cases. Again, it remained unclear if both or only one agent caused bovine abortion. Most recently, W. chondrophila DNA has been detected in vaginal swabs and placental tissues taken from Tunisian cows that had aborted and were also coinfected with Listeria monocytogenes. 8 –10 However, a recent experimental infection study failed to demonstrate a clear link between W. chondrophila infection and abortion in cattle. 185 Only 1 out of 9 heifers intravenously infected with W. chondrophila on day 105 to day 110 of pregnancy had inflammatory lesions of the chorioallantois but this did not result in abortion. Although this study 185 could confirm the third and fourth of Koch’s postulates, it was concluded that W. chondrophila might represent an opportunistic pathogen and that its role in abortion remains to be confirmed. 185
Parachlamydia has been related to bovine abortion cases and to a lesser extent caprine and ovine abortion in Switzerland, England, Scotland, Ireland, Hungary, and Tunisia. 9,18,23,41,82,139,140,186,188,190 Parachlamydial cases revealed necrotizing or purulent to necrotizing placentitis sometimes with vasculitis in common. The presence of a nonsuppurative interstitial pneumonia in fetal lungs was also reported in relation to parachlamydial infection but IHC remained negative. 186 Parachlamydia was demonstrated in placental lesions by PCR, IHC, or TEM. Positive immunolabeling for Parachlamydia in the placenta was present within the cytoplasm of trophoblasts primarily associated with areas of intense infiltration of polymorphonuclear neutrophils. 186 Mixed infections with other abortigenic agents such as Neospora caninum, Coxiella burnetii, C. abortus, Brucella abortus, Campylobacter, bovine herpesvirus type 1 and with mycotic placentitis are reported but interaction with Parachlamydia remains unclear. 9,18,140,186 To confirm an etiological role for Parachlamydia in bovine abortion, its isolation from abortion material and subsequent fulfilment of Koch’s postulates awaits confirmation.
CLO were associated with cattle pneumonia in Scotland, namely P. acanthamoebae, uncultured Chlamydiales and Rhabdochlamydia, 189 and Parachlamydia caused mild respiratory signs in calves when experimentally infected by direct inoculation into the lungs. 94 These signs were dose-dependent and accompanied by macroscopic foci of pneumonic lung lesions corresponding to intrabronchial inoculation sites. Histologically, purulent to fibrinous bronchopneumonia was present at 2 dpi followed by subacute bronchopneumonia at 4 dpi and bronchiolitis obliterans and perivascular cuffing between 10 and 14 dpi. 94 During an intranasal and intratracheal experimental infection mouse model, P. acanthamoebae induced severe interstitial pneumonia which was purulent in the early phase (2–4 days after infection, dpi) and mononuclear at 7–21 dpi. 30,118
Transmission modes of CRB are unknown but aquatic sources might play a role (reviewed in Wheelhouse and Longbottom 187 ). A zoonotic potential has to be considered as CRB have been indirectly linked to recurrent pregnancy failure and miscarriage in women as well as pneumonia and bronchiolitis in children and patients with community-acquired pneumonia. 187
The common abortion diagnostics do not include testing for CRB. Correlation of histological changes such as purulent to necrotizing or necrotizing placentitis with or without vasculitis, detection of the antigen by IHC within lesions and its DNA by PCR is recommended to unambiguously establish a diagnosis of CRB-related abortion. In well-preserved placental specimens, intracellular bacteria and inclusions might be already observed in H&E-stained sections. 21,187 However, more data are needed to learn more about the pathogenicity, associated lesions, significance and transmission routes of CRB-related abortions in ruminants.
Chlamydial Agents of Epitheliocystis in Fish
Epitheliocystis is a disease primarily of the gills of marine and freshwater fish species and is so named due to the presence of cyst-like inclusions within the epithelial cells of the fish gills (reviewed in Stride et al 159 ). More commonly observed in juvenile fish than in adults, clinical signs are sometimes observed in fish with epitheliocystis and may include respiratory distress and lethargy, among other signs. Mortalities are thought to be uncommon but have been reported at rates of up to 100% in juvenile aquaculture production systems for various species. 159
Histology remains the diagnostic method of choice for detecting epitheliocystis. Histopathologically, epitheliocystis infections are often benign without any proliferative host response observable. Where host responses are observed these can appear as local to multifocal epithelial hyperplasia with interlamellar filling that can lead to fusion of the lamellae and cellular hypertrophy. The most severe morphological changes are associated with mortalities in juvenile fish. 159
Prior to the introduction of molecular methods, a chlamydial etiology was long suspected due to the visualization of chlamydial-like cell forms in epitheliocysts by TEM. 35 PCR-based methods utilizing Chlamydiales broad-range PCR primers were subsequently able to confirm a chlamydial origin for these inclusions. 43 Since this confirmation, an increasingly diverse range of novel CRB in the phylum Chlamydiae have been described in >20 fish species, including uncultured agents in a previously described chlamydial family, Simkaniaceae, and 3 entirely newly described families composed exclusively of epitheliocystis agents, Ca. Parilichlamydiaceae, Ca. Piscichlamydiaceae, and Ca. Clavochlamydiaceae. 159 In the absence of a culture system for these pathogens, little is still known about the biology of these novel CRB, although the continued identification of new chlamydial epitheliocystis agents in each fish species examined has led to the suggestion that (a) a significant unrecognized diversity of CRB may be present in fish and (b) each CRB may have coevolved with their respective host. 159 A recent culture-independent genome study of the epitheliocystis agent of orange-spotted grouper (Epinephelus coioides), Candidatus Similichlamydia epinephilii, revealed that while phylogenetically distant from the family Chlamydiacaceae, the genomes of these bacteria are quite similar in size and virulence gene content. 169
Complicating the differential diagnosis of epitheliocystis for pathologists is the recent realization that other bacteria from completely different bacterial phyla including beta- and gamma-proteobacteria 80,152 may cause similar diseases as CRB. Many of these alternative agents may have subtle differences in their morphology compared to chlamydial agents so microscopic differentiation may be possible, 153 however, this will need to be supported by PCR and in situ hybridization for confirmatory diagnosis at this stage.
Conclusions
Zoonotic diseases like chlamydial abortion in ruminants due to C. abortus and psittacosis/ornithosis in birds due to C. psittaci, both presenting with characteristic pathomorphological lesions, have been well known for decades. With the advent of molecular techniques, emerging novel chlamydial species have been detected in a variety of host species and host barriers for known chlamydial species may be less stringent than previously thought. At present, nucleic acid amplification tests are considered the state-of-the-art methods to diagnose chlamydial infections in animals and humans due to their sensitivity and specificity. This might implicate that the pathologist is of minor importance and his or her role might be underestimated in chlamydial diagnostics. However, the behavior of well-characterized chlamydial veterinary pathogens in new hosts is understudied and reports mostly rely on molecular findings only. Moreover, many of the emerging CRB are thought to have the potential to cause disease in their respective hosts, but pathogenesis and associated pathological changes are awaiting proof of this. In the future of chlamydial research, as such, the pathologist is an important player when investigating associated pathology due to natural infection with novel chlamydial species or known chlamydial species in novel hosts or when evaluating the pathogenesis of novel chlamydial species in experimental animal models.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.