
Abstract
The authors describe a spontaneous case of amyloid A (AA) amyloidosis in an adult female Japanese quail (Coturnix japonica). The bird developed AA amyloidosis secondary to chronic peritonitis caused by a Gram-negative bacillus infection. Mild amyloid deposition was also identified in the intestinal tract of apparently healthy adult individuals, suggesting that quail may develop intestinal amyloidosis with age. Based on these observations, it was hypothesized that quail can develop AA amyloidosis following inflammatory stimulation with lipopolysaccharide (LPS). Therefore, adult quail were repeatedly injected with LPS and the development of AA amyloidosis was confirmed. The amyloid deposition in this model increased when quail amyloid was intravenously injected as an amyloid-enhancing factor. The experiments were repeated with young quail, but amyloid deposits were not observed following LPS injections. However, AA amyloidosis did develop when quail amyloid was injected in addition to LPS. These results indicated that adult quail develop AA amyloidosis after inflammatory stimulation with LPS. Furthermore, quail AA amyloidosis was shown to have transmissibility regardless of age. Interestingly, the authors found that administration of chicken amyloid fibrils also induced AA amyloidosis in young quail. This is the first report of cross-species transmission of avian AA amyloidosis.
Keywords
Amyloidosis is a group of disorders caused by deposition of amyloid fibrils that are fibrous insoluble proteins formed as a result of misfolding of several precursor proteins. The disease is divided into localized or systemic types, according to where the fibrils are deposited, and it is classified into 36 types according to the type of precursor protein or pathology. 30 Amyloid A (AA) amyloidosis is one of the systemic amyloidoses, caused by deposition of AA, which occurs in various species, including humans and birds. 2,7,14,15 The precursor protein of AA is serum AA (SAA), which is an acute phase protein mainly synthesized in the liver soon after the levels of interleukin-1 (IL-1), IL-6, and tumor necrosis factor-alpha (TNF-α) become elevated. 26 It is thought that during chronic inflammatory diseases or malignant tumors, the long and sustained supersaturated state of SAA causes it to misfold into AA, resulting in the development of AA amyloidosis.
AA amyloidosis can be experimentally induced in many mice strains by repeated injection of inflammatory stimulants such as silver nitrate or casein. 13,17,31 Furthermore, it can be rapidly induced by administration of allogeneic or xenogeneic amyloid fibrils, which act as an amyloid enhancing factor (AEF) in animals under inflammatory conditions. 3,8 Therefore, AA amyloidosis is thought to be a transmissible disorder similar to prion diseases. There have been occasional reports of outbreaks of AA amyloidosis in zoo and domestic animals, 18,22,32 indicating the horizontal transmissibility of AA amyloidosis under natural conditions, 19 but this route of transmission has not been investigated to date.
Avian AA amyloidosis occurs frequently in waterfowl and chickens secondary to chronic inflammation due to infections. 2,4,15,32 There are some reports of the experimental induction of AA amyloidosis by chronic inflammatory stimulation such as by repeated administration of casein or oil-emulsified bacterins in ducks and chickens. 5,16 Moreover, we have previously reported oral transmission of AA amyloidosis in vaccinated chickens, 20 suggesting that horizontal spread of avian AA amyloidosis within the flock could occur.
Although the experimental chicken model of AA amyloidosis is a valuable transmission model, the birds are hard to handle and require a breeding space appropriate to their large body size. In contrast, Japanese quail are less aggressive and are smaller than chickens, so they have high value as an experimental model that is easy to handle and requires only a small breeding space. 10 However, although there are many reports of AA amyloidosis in ducks and chickens, there is only 1 case report of quail AA amyloidosis that was associated with serositis. 21
In this study, we describe a spontaneous case of quail AA amyloidosis secondary to chronic inflammation due to infection with Gram-negative bacilli leading us to hypothesize that quail might develop AA amyloidosis following inflammatory stimulation with lipopolysaccharide (LPS) and that transmission of amyloid materials would enhance development of amyloidosis in the recipients. Therefore, we performed the following 3 experiments: (1) adult quail AA amyloidosis was induced by repeated administration of LPS, (2) adult quail AA amyloidosis was transmitted between birds of the same species by administration of allogeneic amyloid fibrils, and (3) AA amyloidosis was transmitted between young quail. Furthermore, cross-species transmission of AA amyloidosis by administration of chicken amyloid fibrils also occurred in young quail.
Materials and Methods
Pathology of Spontaneous Quail AA Amyloidosis
Case history
Japanese quail (Coturnix japonica) were reared for breeding purposes under the conventional environment at the Laboratory of Veterinary Physiology in Tokyo University of Agriculture and Technology (TUAT). All birds were kept in wire cages and received ad libitum access to tap water and mashed feed (Quail Cosmos, Aichi, Japan). The birds were recorded as a crossbreed of WE strain (derived from Nisseiken, Tokyo, Japan) and NIES-L strain (derived from National Institute of Environmental Studies, Tsukuba, Japan), although the crossing ratio and number of filial generations were not recorded.
Female quail were given to our laboratory for another experiment after the laying period, and 1 severely emaciated quail was found dead on the day after they were received. The carcass (group A, quail No. 1) was immediately submitted for macroscopic, histopathological, and immunohistochemical examination.
To examine the background lesions of adult Japanese quail, 33 apparently healthy hens (group A, quail Nos. 2–34) and 7 cocks (group A, quail Nos. 35–41) in the same flock were euthanized and autopsied. The research program was approved (27-99) by the Animal Care and Use Committee at TUAT, and the research was performed according to the guidelines for animal experiments at TUAT.
Pathological examinations
All 41 birds were subjected to macroscopic, histopathological, and immunohistochemical examinations. The birds were euthanized by exsanguination after deep anesthesia with 4% isoflurane (DS Pharma Animal Health Co, Ltd, Osaka, Japan). At necropsy, liver, spleen, kidney, heart, lung, proventriculus, duodenum, pancreas, cecum, and pectoral muscle were collected. A number of peritoneal masses were also collected from quail No. 1. Tissues were fixed in 10% neutral buffered formalin and embedded in paraffin. For histopathological examination, 2-μm tissue sections were prepared and stained with hematoxylin and eosin, Congo red (CR) with/without potassium permanganate oxidation, and Gram and Ziehl-Neelsen stains. Amyloid deposits were detected in CR-stained sections as an emerald-green birefringence identified under polarized light. The severity of amyloid deposition in each organ was graded visually in CR-stained sections as follows: 0 = no deposition, 1 = mild deposition only in vessel walls, 2 = mild deposition in vessel walls and interstitial tissues, 3 = moderate deposition in vessel walls and interstitial tissues, and 4 = severe deposition in vessel walls and interstitial tissues. Immunohistochemistry was performed with an anti-chicken SAA1 mouse antiserum (19), anti-human SAA mouse antibody (ab687, Abcam, Tokyo, Japan), or anti-Escherichia coli LPS antibody (bs-2351R, Bioss Inc, Woburn, MA, USA).
Electron microscopy
Formalin-fixed duodenum (group A, quail No. 1) that was histologically identified as AA amyloidosis was cut into smaller pieces, washed in 60-mM 4-(2-HydroxyEthyl)-1-piperazineethanesulfonic acid buffer, and fixed in an equal volume mixture of 2% OsO4 and 0.2 M cacodylate buffer for 1 hour. The tissue was dehydrated through graded alcohol and propylene oxide washes before being embedded in epoxy resin (Polybed812, Polysciences, Warrington, PA, USA). Ultrathin sections (70 nm) were cut using an ULTRACUT N microtome (Leica Microsystems, Wetzlar, Germany) and collected onto collodion/carbon-coated copper grids (Nisshin EM, Tokyo, Japan). Sections were counterstained with EM Stainer (Nisshin EM) and lead citrate and examined with a JEM1400 Plus transmission electron microscope (JEOL, Tokyo, Japan) at an acceleration voltage of 80 kV.
Mass spectrometry–based proteomic analysis
Amyloid fibrils were extracted from formalin-fixed, paraffin-embedded duodenum with severe amyloid deposition (group A, quail No. 1) and digested with trypsin according to the method of Kamiie et al. 9 The tryptic digests were subjected to nanoflow LC−mass spectrometry (MS)/MS system equipped with an Orbi-Trap mass spectrometer (Orbi-Trap XL, Thermo Scientific, Boston, MA, USA) and queried according to the method of Taoka et al. 33 The MS/MS data were converted to a Mascot generic format with the Proteome Discoverer software (Thermo Scientific, ver 1.1). The files were processed with the MASCOT algorithm (version 2.2.1, Matrix Science, London, UK) to assign peptides using the Swiss-Prot sequence database. The chicken Swiss-Prot database was used (release 2016) because there was insufficient information in the database for quail. All results of peptide searches were extracted from the Mascot DAT files using the STEM software, 28 expressed as tab delimited text files, and pooled in a Microsoft Excel spreadsheet.
Sequence alignment of SAA
Though the amino acid sequence of quail SAA was not determined, the predicted amino acid sequence was obtained from the NCBI Protein Database (XP_015719668.1), and the signal peptide of quail SAA was predicted by SignalP 4.1. 24 The amino acid sequence of chicken SAA and human SAA1 were obtained from UniProt Knowledgebase (chicken, D6P887; human, P0DJI8). Then, all sequences were aligned by ClustalW. 34
Experimental Induction of Quail AA Amyloidosis
Animals
Fourteen adult male and 41 adult female Japanese quail that were no longer required for breeding purpose were obtained from Laboratory of Veterinary Physiology. Fertilized eggs were also obtained from Laboratory of Veterinary Physiology to provide 13 male and 12 female 5-week-old Japanese quail. These individuals were recorded as a crossbreed of WE strain and NIES-L strain, although the crossing ratio and number of filial generations were not recorded. Between 1 and 6 quail were housed per cage and maintained under conventional conditions. Tap water and mashed feed (Toyohashi Feed Mills Co, Ltd, Aichi, Japan) were supplied without restriction. At necropsy, the birds were euthanized by exsanguination after deep anesthetization with 4% isoflurane (DS Pharma Animal Health Co, Ltd). The research program was approved (27-99) by the Animal Care and Use Committee at TUAT and the research was performed according to the guidelines for animal experiments at TUAT.
Experiment 1: Experimental induction of AA amyloidosis in adult quail
Twenty-seven adult female quail were used for the experiment. Quail in group B were given intraperitoneal (i.p.) injections of 0.5 ml phosphate buffered saline (PBS) twice a week. Quail in groups C through G were injected i.p. twice a week with 2, 4, 8, 16, or 32 mg/kg bodyweight (BW) of LPS (Sigma-Aldrich Japan, Tokyo, Japan) diluted in 0.5 ml PBS, respectively. Quail in groups B through G were euthanized and autopsied 5 weeks after the first injection. The doses of LPS were set up as nonlethal and pro-inflammatory doses, referring to the dose required to cause an acute phase response in quail, 1.0 to 22.5 mg/kg BW, 12 and the nonlethal dose in chickens, 517 mg/kg BW. 1 At necropsy of groups B through G, spleen samples were collected and frozen for use as AEF in the following experiments.
Experiment 2: Experimental transmission of AA amyloidosis in adult quail
Fourteen adult male and 14 adult female quail were used for the experiment. The frozen spleen samples from birds that developed splenic amyloid deposition in groups C through G was diluted 20 times in saline and used as quail AEF (qAEF). Quail in groups H through M were injected i.p. twice a week with 2 mg/kg BW of LPS solution diluted in 0.5 ml PBS. Quail in groups K through M were additionally injected intravenously with 0.2 ml qAEF at the time of first administration of LPS. All quail in groups H through M were euthanized and autopsied at 1 (groups H and K), 3 (groups I and L), or 5 (groups J and M) weeks after the first administration of LPS.
Experiment 3: Experimental induction and allogeneic or xenogeneic transmission of AA amyloidosis in young quail
Thirteen male and 12 female 5-week-old quail were used for the experiment. Quail in groups P through T were injected i.p. twice a week with 2 mg/kg BW of LPS solution diluted in 0.5 ml PBS. Quail in groups R and S were additionally injected intravenously with 0.2 ml qAEF at the time of first administration of LPS. Quail in group T were intravenously injected with 0.2 ml chicken AA fibrils that were extracted by Pras’s method 25 from livers of AA amyloidosis-affected chickens as chicken AEF (cAEF) at the time of first administration of LPS. Quail in groups P through T were euthanized and autopsied at 2 (groups P and R) or 5 (groups Q, S, and T) weeks after the first administration. To examine the background lesions in untreated controls of the same age, quail in groups N and O were euthanized and autopsied at 7 or 10 weeks old, respectively.
Pathological examination
Liver, spleen, kidney, heart, lung, proventriculus, duodenum, pancreas, cecum, and pectoral muscle samples were collected at necropsy. Methods for histopathology, histopathologic scoring, and immunohistochemistry were as described previously. The amyloid index (AI) scores of each individual were calculated by dividing the total deposition score of all organs by the number of organs examined. AI scores of each group were statistically analyzed to determine the promotion of amyloid deposition by LPS injection in experiment 1 (group B vs groups C–G, t-test), examine the enhancing effect of AEF injection on amyloid deposition in experiment 2 (groups H and J vs groups K–M, t-test), or compare the deposition levels by experimental periods (group R vs group S, t-test) and amyloidogenicity between qAEF and cAEF (group S vs group T, t-test) in experiment 3. In experiment 1, the dose-response relationship between LPS level and AI score was examined by Spearman’s rank correlation coefficient.
Results
Pathology of Spontaneous Quail AA Amyloidosis
Pathological examinations
The necropsy of quail No. 1 (group A) revealed abdominal distention and severe atrophy of the pectoral muscle. Pale yellow or white spherical masses of 2 to 20 mm diameter were observed on the peritoneum and surface of the liver and intestine (Fig. 1). The duodenum contained a large mass and had severe hypertrophy of the wall. The spleen was discolored and enlarged. The oviduct contained a large mass with yolk-like material on the cut surface. Significant macroscopic lesions were not observed in quail Nos. 2 through 41 at necropsy.
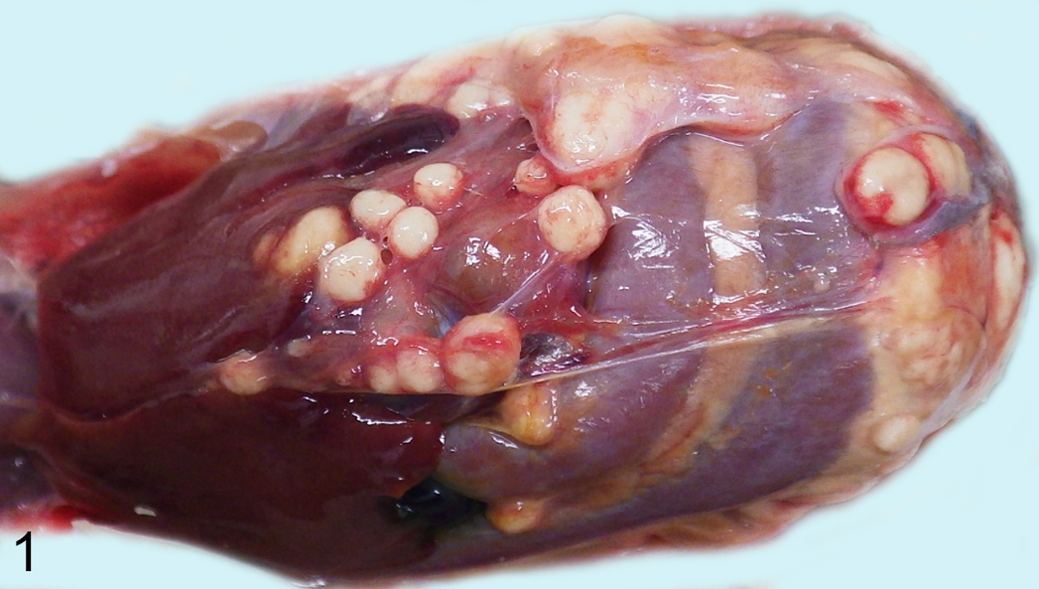
Amyloid A amyloidosis, Japanese quail, No. 1 in group A. Multiple masses are present on the peritoneum and serosa of the liver and intestine.
The distribution of amyloid deposition in quail Nos. 1 to 41 is shown in Figure 2. In quail No. 1, slight to severe amyloid deposition was observed histologically in all of the organs examined, including the lamina propria of the intestine, interstitium of the pancreatic acinus, and nearly all of the splenic parenchyma. In quail Nos. 2 through 41, mild amyloid deposition was frequently observed in the small vessel walls of the duodenum and pancreas and peribronchial interstitium and infrequently in the splenic vessel walls.
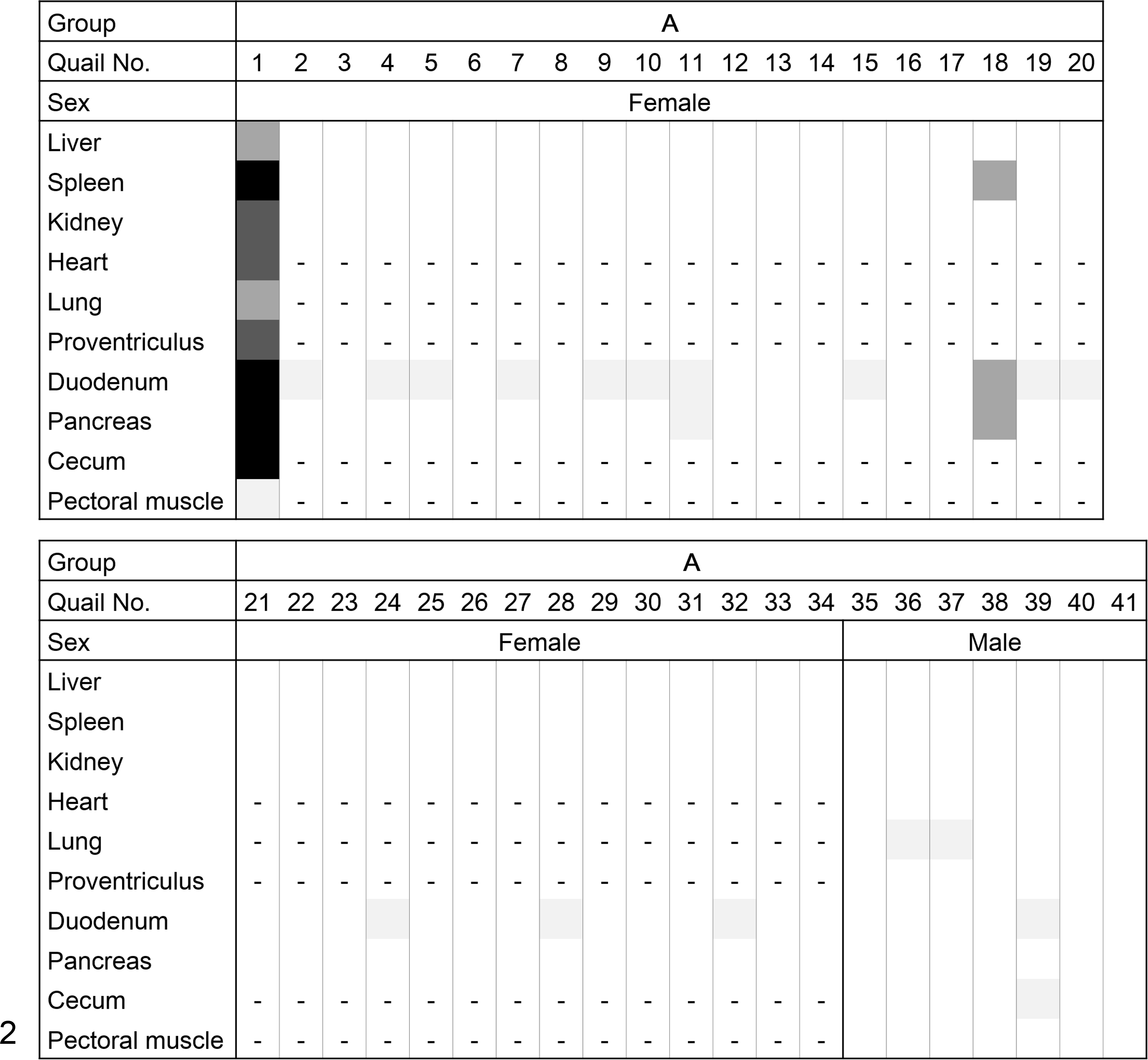
Heat map of the distribution and intensity of amyloid deposition in group A (spontaneous amyloidosis in adult quail). The severity grade of amyloid deposition is represented as: 4, black; 3, dark gray; 2, mid gray; 1, faint gray; 0, white; and -, no data.
All amyloid deposits stained positive for CR (Fig. 3a, b) but were negative after pre-incubation with potassium permanganate. By immunohistochemistry, amyloid deposits were negative using anti-chicken SAA1 antiserum but positive using anti-human SAA antibody (Figs. 4, 5). However, the cytoplasm of hepatocytes showed granular immunoreactions to the anti-chicken SAA1 antiserum.
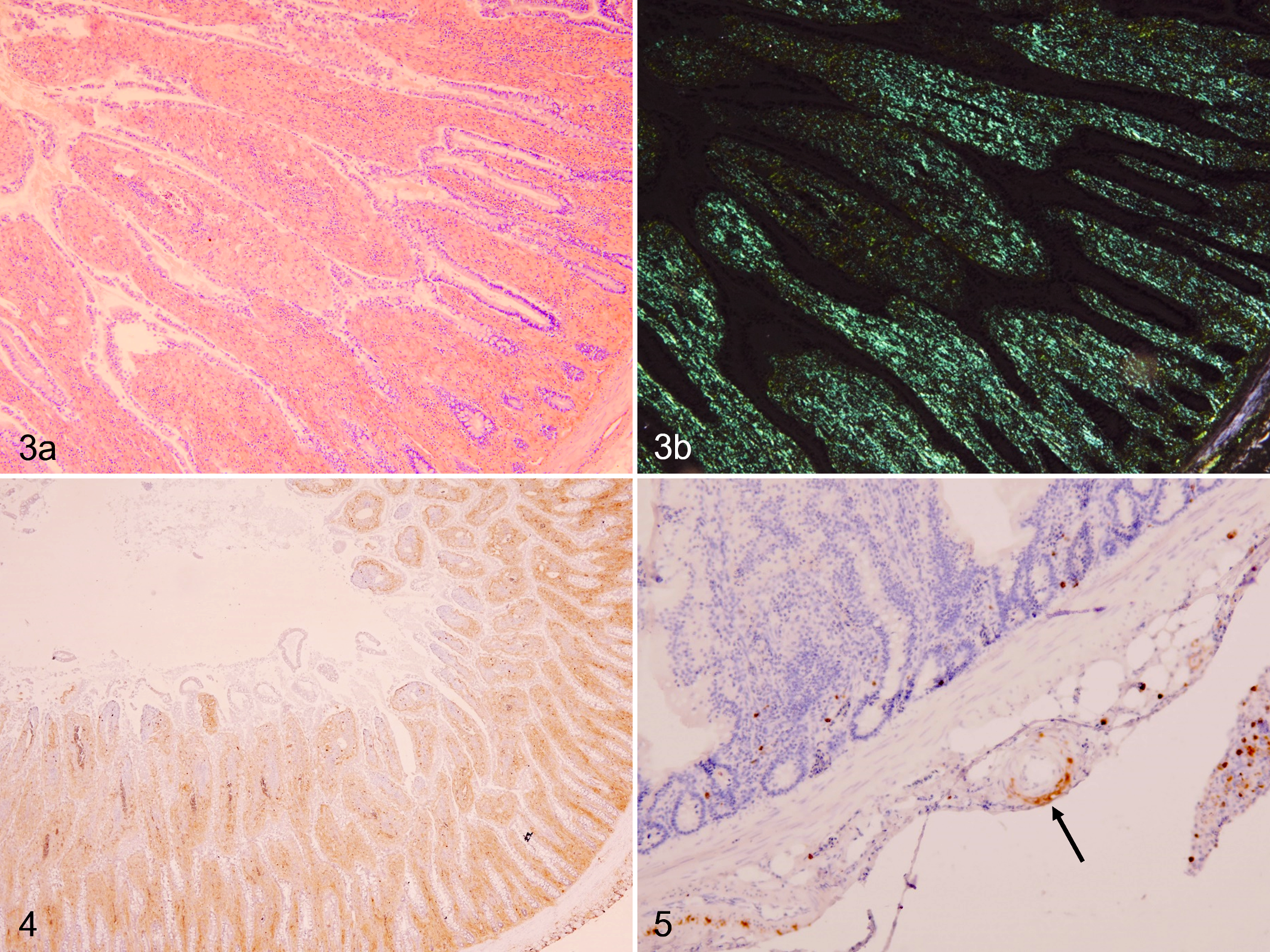
In quail No. 1, the peritoneal masses were composed of abscesses or multifocal necrosis. Many short Gram-negative bacilli that were negative for Ziehl-Neelsen and immunoreactive to anti-E.coli LPS antibody (Fig. 6) were observed in the peripheral zone of the abscesses.
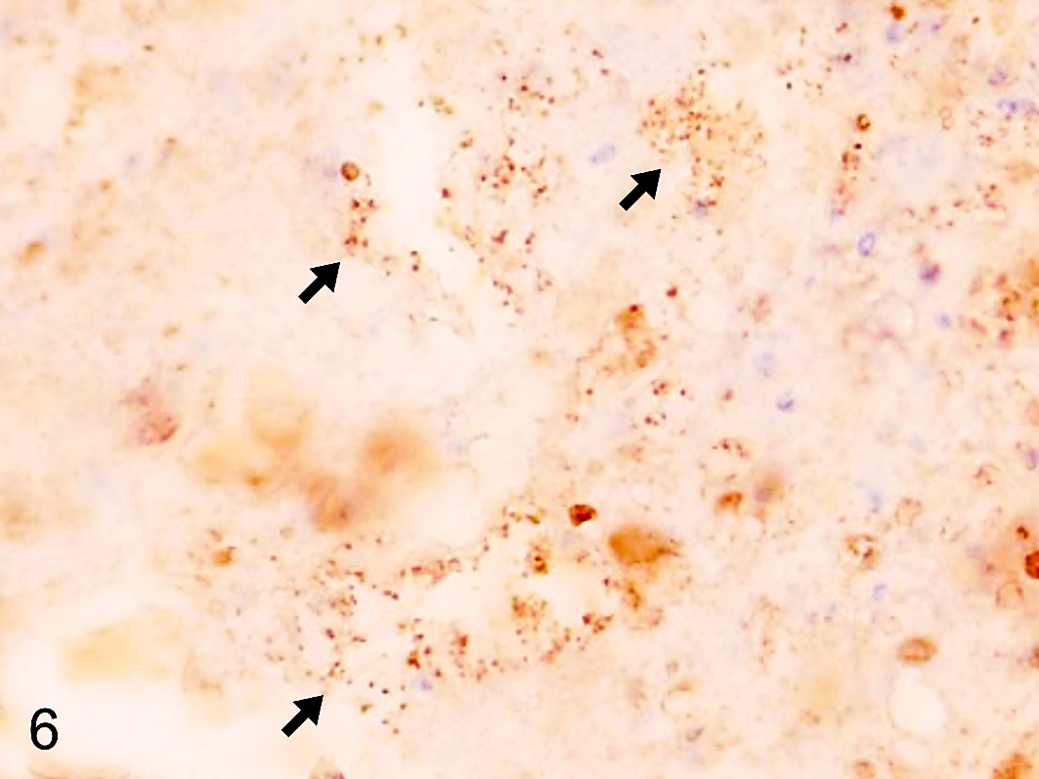
Peritoneal mass, quail. Group A, quail No. 1. Multiple immunolabeled short bacilli are observed at the peripheral zone of the mass (arrows). Immunohistochemistry using an antibody to E.coli lipopolysaccharide.
Ultrastructurally, the amyloid fibrils in the lamina propria mucosae of the duodenum were nonbranched and straight-chained (Fig. 7).
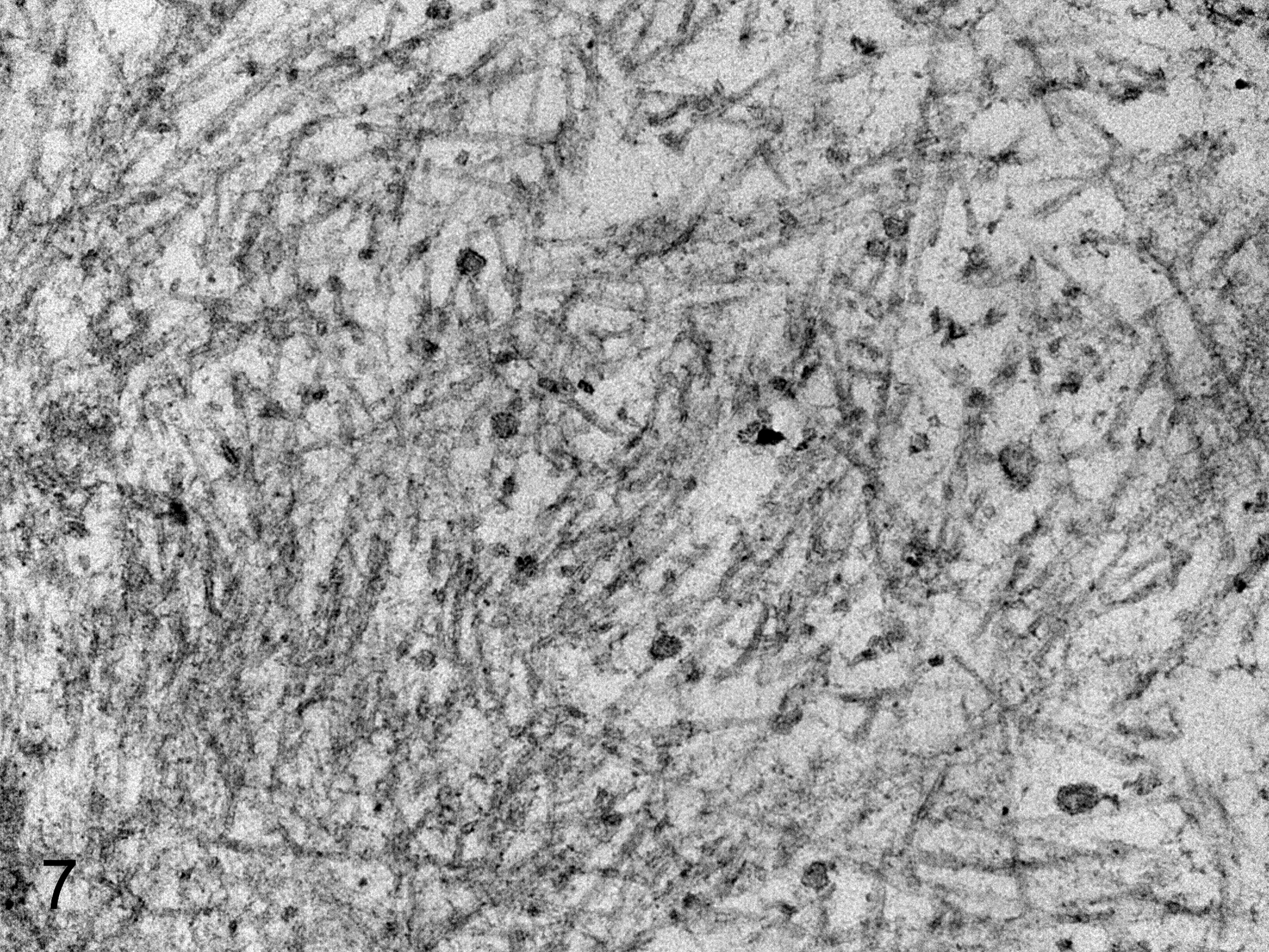
Amyloidosis, duodenum, quail. Quail No. 1. Amyloid fibrils are observed as nonbranched and straight-chained fibrils in the lamina propria. The widths of the amyloid A fibrils ranged from 7.7 to 10.8 nm. Transmission electron microscopy.
MS-based proteomic analysis
MS-based proteomic analysis provided reliable MS/MS spectra and identified a precursor ion with a m/z value of 552.27, corresponding to the tryptic peptide of EANYIGADKYFHAR identified as chicken SAA (Suppl. Fig. S1). This sequence corresponded perfectly to residues 48 through 61 of quail SAA (Fig. 8). From these results, the extracted amyloid was identified as AA.

Sequence alignment of serum amyloid A (SAA) in quail and chicken. White bar, predicted signal peptide (residues 1–18). Gray bars, predicted AA-forming fragment (residues 19–99). Cross-hatched bar, amino acid sequence detected by LC/MS/MS (residues 48–61). Black bar, antigen for anti-chicken SAA antiserum (residues 105–127).
Sequence alignment of SAA proteins
Figure 8 shows the sequence alignment of SAA in quail and chicken. By SignalP 4.1 analysis, the signal peptide of quail SAA was predicted as residues 1 through 18 quail SAA. By sequence alignment, residues 19 through 99 of quail SAA corresponded to residues 1 through 76 of human mature SAA1 6 and were predicted as the AA-forming fragment. The amino acid residue EANYIGADKYFHAR that was detected by MS-based proteomic analysis was within the predicted AA-forming fragment.
Experimental Induction of Quail AA Amyloidosis
Experiment 1: Experimental induction of AA amyloidosis in adult quail
The distribution and intensity of amyloid deposition in each quail in groups B through G is shown in Figure 9. Amyloid deposits in each tissue were stained with CR and presented emerald-green birefringent streaks under polarized light (Suppl. Figs. S2–S5). Deposited amyloid was defined as AA by IHC analysis using anti-human SAA antibody. Quail Nos. D-1, G-1, and G-2 died after the 9th, 7th, and 7th LPS injections, respectively, and were not used for statistical analysis. Quail in group B showed minor to mild amyloid deposition in the duodenum and lung. In contrast, quail injected with >2 mg/kg BW of LPS frequently showed severe amyloid deposition, especially in the digestive system. The mean AI scores of group C were significantly higher than group B (P < .01), and those of groups D through G also showed an upward tendency compared with group B (Fig. 12a). Spearman’s rank correlation coefficient did not reveal a clear dose relationship between groups C through G.

Heat map of the distribution and intensity of amyloid deposition in groups B–G (adult quail injected with lipopolysaccharide). The severity grade of amyloid deposition is represented as: 4, black; 3, dark gray; 2, mid gray; 1, faint gray; 0, white; and -, no data.
Experiment 2: Experimental transmission of AA amyloidosis in adult quail
The distribution and intensity of amyloid deposits in each quail in groups H through M is shown in Figure 10. Adult quail injected with only LPS showed a high rate of amyloid deposition from 1-week post-injection (WPI), and deposition levels became more severe over time. Adult quail in qAEF-injected groups showed remarkable amyloid deposition in the spleen at 3 and 5 WPI, and their AI scores also increased over time (Fig. 12b). The AI scores of group L were significantly higher than group I (P < .05). There were no obvious sex differences in amyloid deposition levels in any of the groups.

Heat map of distribution and intensity of amyloid deposition in groups H through M (adult quail injected with lipopolysaccharide; groups K–M were also injected with quail amyloid-enhancing factor). The severity grade of amyloid deposition is represented as: 4, black; 3, dark gray; 2, mid gray; 1, faint gray; 0, white; and -, no data.
Experiment 3: Experimental induction and allogeneic or xenogeneic transmission of AA amyloidosis in young quail
The distribution and intensity of amyloid deposits for each quail in groups N through T is shown in Figure 11. There were no amyloid deposits in the control (N and O) and LPS-injected (P and Q) groups until 5 WPI (Suppl. Fig. S6). In contrast, young quail injected with qAEF showed remarkable amyloid deposition in the spleen from 2 WPI, which became systemic and severe at 5 WPI (Suppl. Fig. S7). The AI scores of the qAEF-injected groups increased over time (P < .01, Fig. 12c). Mild to moderate amyloid deposits in the spleen and digestive organs were observed in birds injected with cAEF (group T). The AI score of young quail injected with cAEF (group T) was significantly lower than those injected with qAEF (group S) (P < .01, Fig. 12c). There were no obvious sex differences in amyloid deposition levels between the groups. The schematic overview of experiments 1, 2, and 3 is shown in Figure 13.
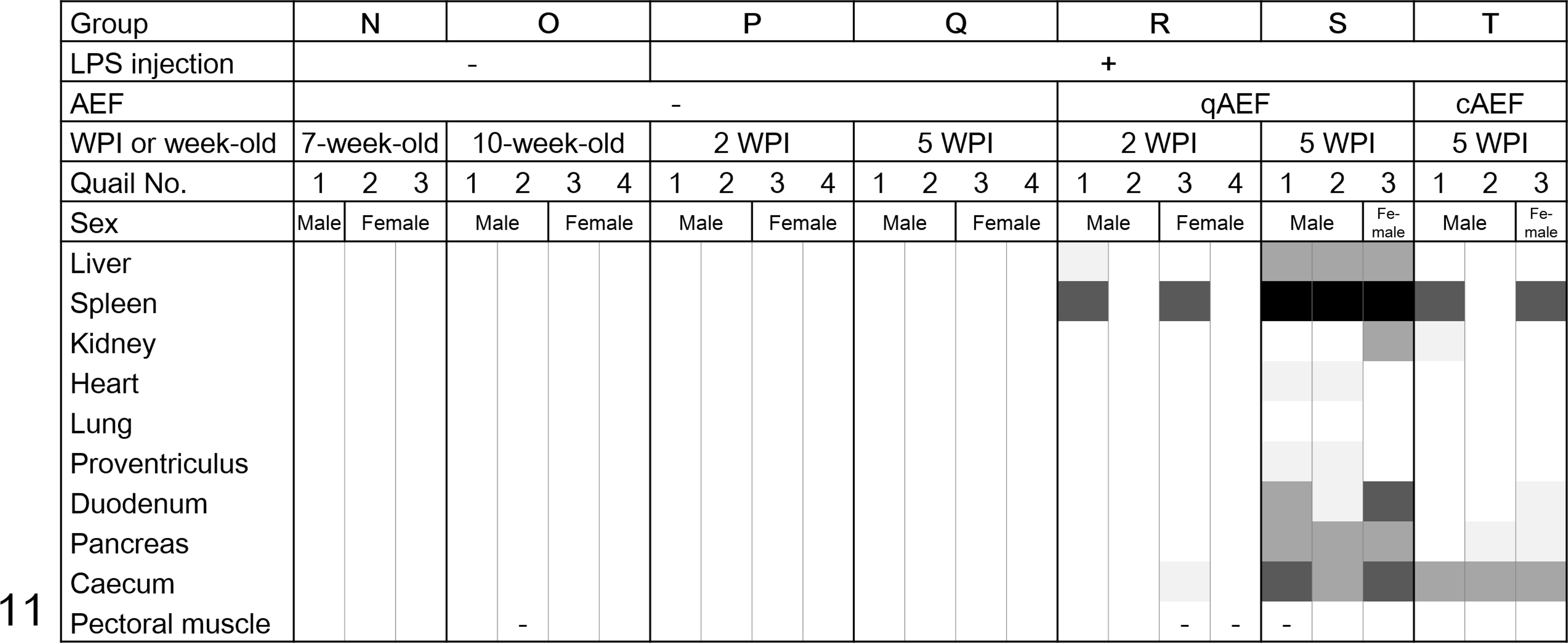
Heat map of the distribution and intensity of amyloid deposition in groups N through T (young quail injected with lipopolysaccharide; groups R and S and T were also injected with amyloid-enhancing factors derived from quail and chickens, respectively). The severity grade of amyloid deposition is represented as: 4, black; 3, dark gray; 2, mid gray; 1, faint gray; 0, white; and -, no data.
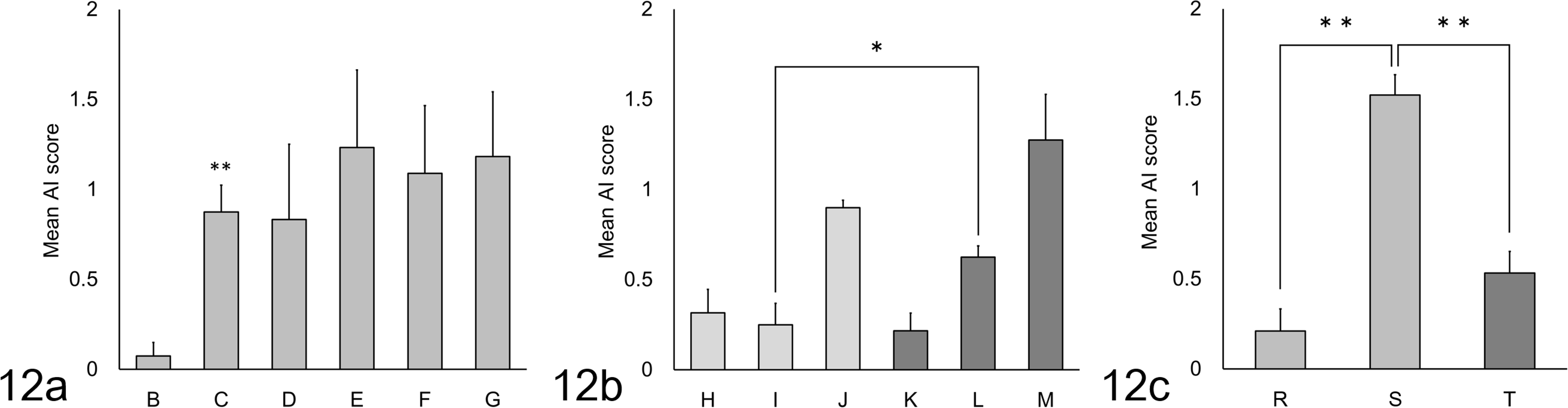
Mean amyloid index (AI) scores in Japanese quail. (a) Experiment 1. **Significantly different versus group B by t-test (P < .01). (b) Experiment 2. *Significantly different by t-test (P < .05). (c) Experiment 3. **Significantly different by t-test (P < .01). The groups shown on the x-axes are: B, adults injected intraperitoneally (IP) with saline; C-G, adults injected IP with 2, 4, 8, 16, or 32 mg/kg of LPS respectively; H-J, adults injected with 2 mg/kg LPS then euthanized at 1, 3 and 5 weeks respectively; K-M, adults injected with 2 mg/kg of LPS plus quail amyloid enhancing factor (AEF) then euthanized at 1, 3 and 5 weeks respectively; R, juveniles injected with 2 mg/kg of LPS and quail AEF then euthanized at 2 weeks; S, juveniles injected with 2 mg/kg of LPS and quail AEF then euthanized at 5 weeks; T, juveniles injected with 2 mg/kg of LPS and chicken AEF then euthanized at 5 weeks.
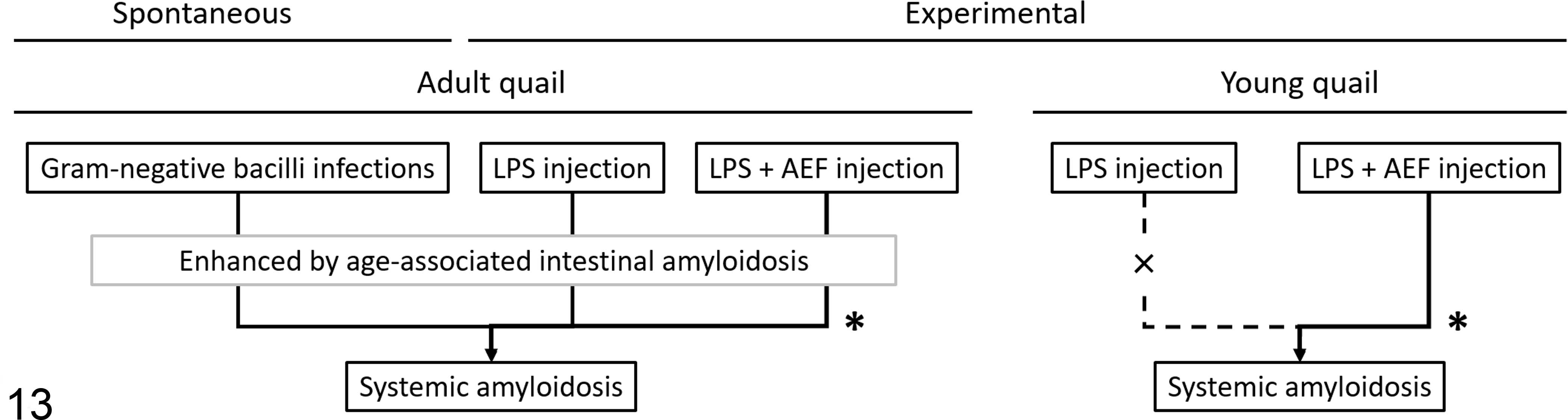
Schematic representation of spontaneous and experimental quail amyloid A amyloidosis. Adult quail develop systemic amyloidosis as a result of Gram-negative bacterial infection or from lipopolysaccharide (LPS) injection with or without amyloid enhancing factor (AEF) injection. Age-associated intestinal amyloidosis is thought to be an aggravating factor for systemic amyloidosis. Young quail develop systemic amyloidosis only as a result of injection of both LPS and AEF. *Accompanied by splenic amyloid deposition.
Discussion
AA amyloidosis was diagnosed in quail No. 1 in group A by immunohistochemistry and MS-based proteomic analysis. The bird may have developed AA amyloidosis secondary to chronic inflammation caused by peritonitis with internal ovulation because at necropsy, the oviduct was found to have been replaced by a yolk-like mass. Histological and immunohistochemical analysis revealed short Gram-negative bacilli that were immunoreactive to anti-E.coli LPS antibody around the intraperitoneal abscess. Because there was no outbreak of disease in other birds from the same flock, it is possible that this quail might have suffered from an opportunistic infection that caused the inflammation.
The anti-chicken SAA antiserum used in this study was produced by injection of a synthetic peptide (consisting of amino acid residues 105–127 of chicken SAA) into mice as an antigen. By immunohistochemistry, we found that this antiserum did not label amyloid deposits in quail but bound to the cytoplasm of hepatocytes, consistent with binding to quail SAA but not to quail AA. By sequence alignment of human SAA1 and quail SAA, residues 19 through 99 of quail SAA were predicted as the AA-forming fragment, which could explain the absence of any reaction with antiserum raised against the C-terminal side of SAA. SAA is usually detected in AA deposits as a conjugated protein. 23 The anti-chicken SAA antiserum used in this study has previously been shown to react positively with amyloid deposits in chickens. 11,20 In contrast, the antiserum was negative for amyloid deposits in quail, suggesting that they rarely contain precursor proteins.
Amyloid deposition was observed not only in quail No. 1 but also in apparently healthy quail in group A at an unexpectedly high rate (17/40). Amyloid deposits in these quail were immunoreactive to anti-AA antibodies, indicating AA amyloidosis. The low-level AA deposition was considered as an age-associated lesion as reported in other species, including Pekin duck. 2 In mice, it is known that SAA3s are located in the mucosa and provide mucosal protection. 27 It is possible that the precursor protein of quail AA could have a similar physiological function in the mucosa.
The amyloid deposition in quail injected with LPS occurred at a high rate and was severe compared with the control group, indicating that adult Japanese quail develop AA amyloidosis following repeated administration of LPS. Although the mean AI score was significantly higher in group C, a clear dose response was not identified, indicating that the administration of 2 mg/kg BW of LPS was sufficient to induce amyloidosis in adult quail. In previous studies of experimental avian amyloidosis, intravenous (IV) injection of bacterial liquid or intramuscular administration of inactivated vaccines were used as inflammatory stimuli to induce AA amyloidosis. 5,16 The experimental chicken transmission model of AA amyloidosis in previous research showed that amyloid deposition was induced by intramuscular injection of a Salmonella Enteritidis (SE) vaccine but not by a Mycoplasma gallisepticum vaccine. 20 Taken together with our finding that a quail developed AA amyloidosis secondary to Gram-negative bacilli peritonitis, we suggest that bacterial stimulation, especially exposure to Gram-negative bacilli, enhances the sensitivity of fowl to development of AA amyloidosis.
The amyloid deposition in the digestive system of quail became worse from 1 WPI in LPS-injected groups H and J. In contrast, amyloid deposition was never observed in young birds in groups P and Q despite injection of LPS. These results strongly suggest that amyloid deposition in the digestive organs and lungs of adult quail is an age-associated lesions. In the experimental model for chicken AA amyloidosis, amyloid deposits were primarily observed in spleen or liver. 20 However, in this study, amyloid deposition in quail was observed especially in the digestive system, suggesting that the spontaneous age-associated deposits of amyloid may act as a seed leading to severe amyloid deposition in the digestive system following a subsequent inflammatory stimulus such as LPS administration.
In experiment 2, the amyloid deposition at 3 WPI was significantly enhanced by IV administration of qAEF, suggesting that transmission of AA amyloidosis can occur in quail. In the experimental model of AA amyloidosis transmission, amyloid deposits were primarily observed in the spleen. 20,29 We also observed amyloid deposits in the spleen of qAEF-injected groups in this experiment. On the other hand, severe amyloid deposition was also seen in the digestive tract in both the LPS- and qAEF-injected groups, so we could not specifically determine the effect of AEF on each organ.
To assess the effect of AEF without age-associated lesions, experiment 3 was performed with young quail. There were no amyloid deposits in the untreated control and LPS-injected groups until 5 WPI. In contrast, qAEF-injected groups showed remarkable amyloid deposition in the spleen from 2 WPI and systemic and severe amyloid deposition at 5 WPI. These results indicate that amyloid primarily deposits in the spleen and spreads to other organs over time in experimental AA amyloidosis in young quail. By using a young quail model, experiments could be performed without the effect of age-associated lesions.
By sequence alignment, chicken SAA had high homology to predicted quail SAA. Furthermore, the amino acid residue EANYIGADKYFHAR that was detected from quail AA coincided with the chicken SAA sequence. Therefore, we examined cross-species transmission of AA amyloidosis between chicken and quail. Interestingly, the amyloid deposits were also seen in young quail injected with chicken AA fibrils as a xenogeneic AEF. Although chickens are closely related to Japanese quail and belong to the same Phasianidae family, 35 this is the first report of cross-species transmission of avian AA amyloidosis. These results suggest that avian AA amyloidosis could be transmitted to genetically heterogeneous birds.
In this study, we report the pathological features of AA amyloidosis in adult Japanese quail. We describe a spontaneous case of AA amyloidosis that occurred without vaccination in a Japanese quail. Mild amyloid deposition in apparently healthy birds was thought to be an age-associated lesion. Quail AA amyloidosis has rarely been reported despite the fact that quail are not a minor domestic poultry species. Furthermore, we developed an experimental model of quail AA amyloidosis by repeated administration of LPS. We revealed that quail AA amyloidosis was transmissible by the administration of LPS and allogeneic or xenogeneic amyloid fibrils. Because Japanese quail are less aggressive and smaller than chickens, they have high value as an experimental model of avian AA amyloidosis. Little is known about avian AA amyloidosis, and this quail model is expected to contribute to future studies of avian AA amyloidosis.
Footnotes
Supplementary material for this article is available online.
Acknowledgments
We would like to thank Dr Toshiaki Isobe for his assistance with mass spectrometry–based proteomic analysis. We also thank Dr Satoshi Nakaba for his excellent technical assistance in ultrathin sectioning.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was partially supported by JSPS KAKENHI, Grant Number 16H05027 and 17K17702.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.