
Abstract
Horses affected by chronic piroplasmosis may develop poor performance and muscle atrophy. Here we investigate the pathological and immunopathological aspects of myopathy occurring in chronic equine piroplasmosis. The study included 16 horses serologically positive for equine piroplasms presenting with clinical signs and supporting serum biochemical evidence of a myopathy. Skeletal muscle was evaluated by histopathology, immunohistochemistry, indirect immunofluorescence, and molecular detection of piroplasms and inflammatory cytokines in skeletal muscle. Histologic lesions included muscle fiber atrophy (100% of cases), degenerative changes (13/16, 81%), and perivascular perimysial and endomysial lymphocytic infiltrates (81% of cases). In 15 cases (94%), muscle fibers had strong immunostaining for major histocompatibility complex classes I and II. T lymphocyte populations were mainly CD3+, CD8+, and CD4+ in equal proportions, with a lower number of CD79α+ cells. The serum from affected horses was tested by indirect immunofluorescence for binding of IgG, IgM, or IgA to sections of normal equine muscle to detect circulating autoantibodies against muscle antigen(s). In all cases, distinct sarcolemmal staining was detected in sections incubated with serum from affected horses, in contrast to sections incubated with phosphate-buffered saline or equine control sera. Reverse transcription polymerase chain reaction (RT-PCR) testing of muscles from affected animals revealed a significant increase of interferon-γ, interleukin-12, and tumor necrosis factor–α gene expression compared to healthy controls. Theileria equi or Babesia caballi was not detected in samples of affected muscle by RT-PCR. Thus, inflammatory myopathy associated with equine piroplasmosis may involve an autoimmune pathogenesis with upregulation of inflammatory cytokines that may cause myofiber atrophy and degeneration.
Equine piroplasmosis is a tick-borne protozoal disease of horses, donkeys, mules, and zebra. 19,57,59 The etiological agents are 2 apicomplexan hemoprotozoa, Theileria equi 47 and Babesia caballi, 50 that are mainly transmitted by ixodid ticks. Equine piroplasmosis (EP) occurs in tropical, subtropical, and temperate regions, and it is maintained within equine populations as long as competent arthropod vectors are present. 8,10,19,31,59,71 In Italy, especially in the central-southern regions, EP can be considered endemic, and high rates of seropositive animals are reported. 6,29,41,48
Clinically, piroplasmosis can cause acute to chronic forms in adult equids, 19,57,59 as well as neonatal syndromes and abortion in mares. 19,66 With acute T. equi infection, clinical signs are usually related to marked hemolysis and varying degrees of thrombocytopenia, hypophosphatemia, anemia, and hyperbilirubinemia that coincide with high levels of parasitemia. 18,19
Chronic T. equi or B. caballi infection can present with nonspecific signs, including lethargy, anorexia, weight loss, and poor performance. 69 Anemia may be minimal or absent in equids with chronic or persistent infection; these animals are termed inapparent carriers, have no appreciable signs of disease, and are reservoirs for tick and iatrogenic transmission. 61 Among racehorses, seropositive animals can show decreased performance compared with seronegative horses and may be at risk for developing overt clinical disease or even sudden death. 32,61
The response of the equine immune system to piroplasm infection is considered complex and multifaceted. 69 No cross-protection between T. equi and B. caballi is documented, as horses can be infected with both parasites simultaneously. A humoral response is induced 9 and correlates with control of parasitemia. 12 The function of cell-mediated immunity in piroplasmosis has yet to be fully determined, 5,69 although it is considered essential as in other hemoprotozoan infections such as Babesia bovis; it is generally accepted that CD4+ T lymphocytes play an important role in immune defense against apicomplexan parasites, including Plasmodium, Toxoplasma, and Babesia. 24 Development of antierythrocyte antibodies has been implied in the pathogenesis of anemia in human, murine, canine, and bovine piroplasmosis. 1,28
Causes of inflammatory myopathy in horses include bacterial (Actinobacillus equuli, Clostridium spp., Streptococcus equi infections) and parasitic (Sarcocystis fayeri) infections and immune-mediated (purpura hemorrhagica, idiopathic immune-mediated myositis) conditions. 2,20,43
In the present study, we describe for the first time an inflammatory myopathy in horses affected by chronic piroplasmosis and provide insight into the pathogenesis.
Materials and Methods
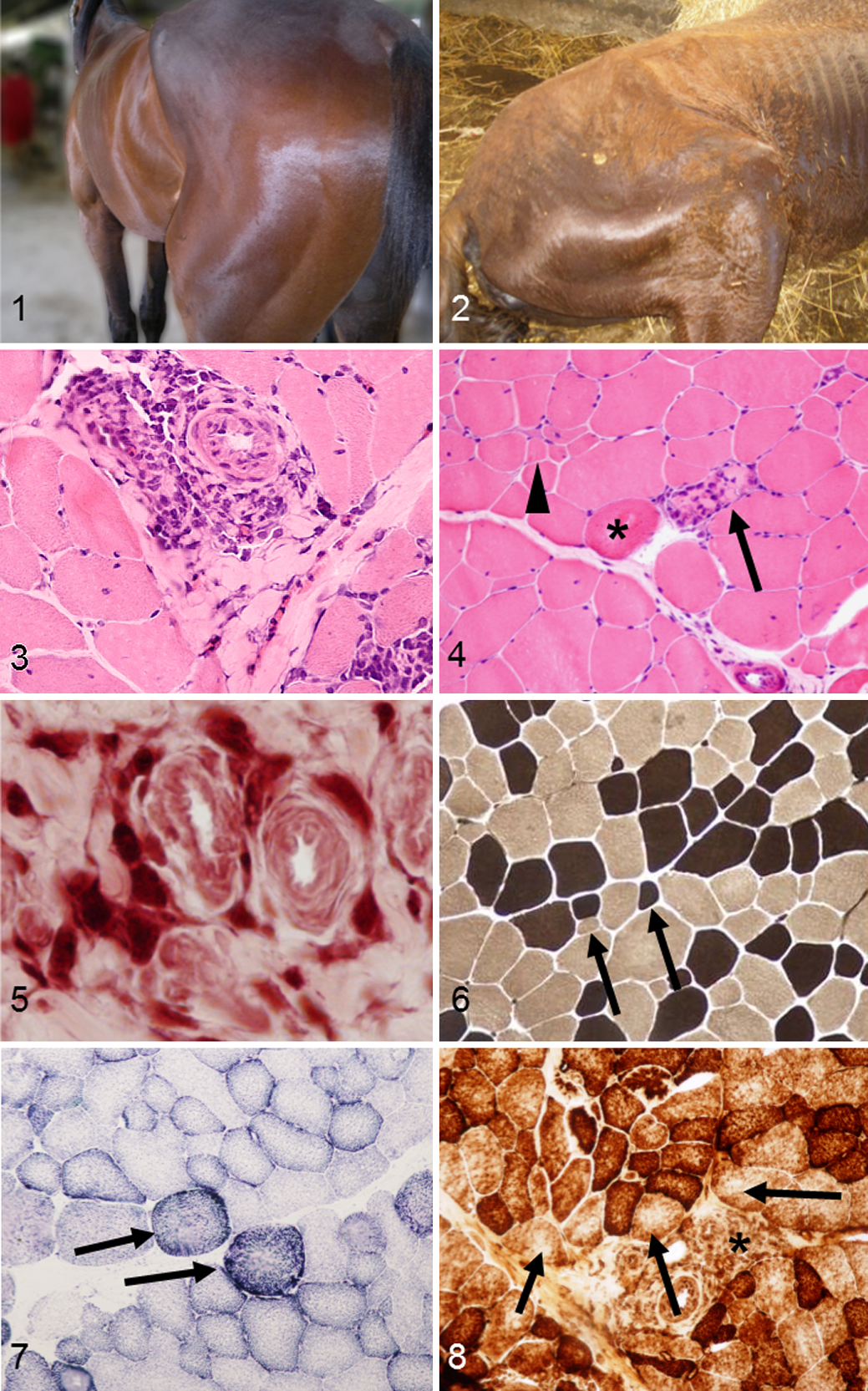
The study included 16 horses, 10 females and 6 males, aged from 3 to 28 years, referring with mild to severe clinical signs of myopathy and that were serologically positive by indirect fluorescent antibody test (IFAT) for one or both agents of piroplasmosis (cutoff of 1:80). More information about the animals’ signalment and clinical signs are reported in Table 1. Clinical signs included mild to severe muscle atrophy (Figs. 1, 2), poor performance, fatigue, and weight loss. None of the studied animals showed hematologic or serum biochemical alterations consistent with acute piroplasmosis or clinical signs such as fever, anemia, or hemoglobinuria. Horses seropositive for equine herpes virus type 1 or 4 or with a clinical history of strangles were excluded from the study.
Clinical, Serum Biochemical, and Parasitologic Findings in 16 Horses With Myopathy Associated With Chronic Piroplasmosis.
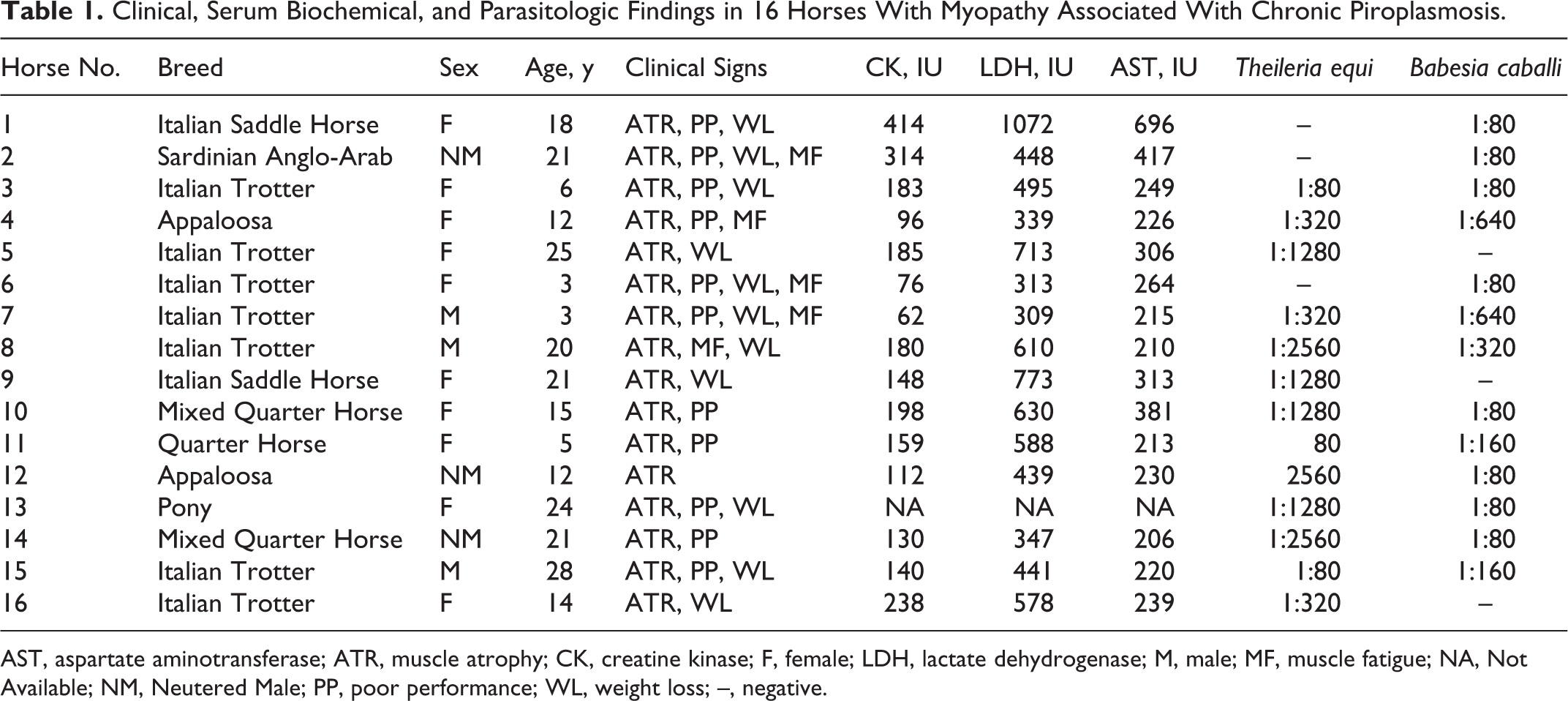
AST, aspartate aminotransferase; ATR, muscle atrophy; CK, creatine kinase; F, female; LDH, lactate dehydrogenase; M, male; MF, muscle fatigue; NA, Not Available; NM, Neutered Male; PP, poor performance; WL, weight loss; –, negative.
Normal control specimens of equine muscle (taken from semitendinosus muscle) for immunohistochemical and molecular studies were selected from the archive of the Laboratory of Neuromuscular Pathology of the Department of Veterinary Medicine and Animal Production of Naples, from horses serologically negative for both agents of piroplasmosis and without histological evidence of myopathy.
All sampling procedures from animals were performed for diagnostic purposes; thus, the study did not require consent or ethical approval according to European Directive 2010/63/EU. Animal sampling, including muscle biopsies, was performed by a veterinary surgeon (M.P.P.) after receiving the consent of the owner. Each owner approved the use of tissues for research purposes, according to the internal rules of Diagnostic Service of the Department of Pathology and Animal Health of the University of Naples Federico II.
Hematology and Serum Biochemistry
Blood samples were collected from the jugular veins into vacuum tubes (Vacutainers; Becton Dickinson, Franklin Lakes, NJ), one with EDTA for complete blood count (CBC) and the other one without anticoagulant for serum biochemistry. Samples were sent to the laboratory on ice packs within 2 hours from collection, immediately centrifuged, and processed. CBC was performed using a cell counter analyzer (Cell Dyn 3700; Abbott, Chicago, IL), and a complete biochemical profile (sodium, potassium, chloride, iron, ionized calcium, urea, creatinine, bilirubin, aspartate aminotransferase [AST], alanine aminotransferase [ALT], lactate dehydrogenase [LDH], creatine kinase [CK], γ-glutamyltranspeptidase [GGT], total proteins, albumin, albumin-globulin ratio, globulin) was performed on samples of parasitized animals using an ADVIA 1650 Chemistry System (Siemens Healthcare, Tarrytown, NY). Serum from 2 healthy horses that were serologically negative to both agents of EP protozoa were also collected.
Parasitological Diagnosis
Sera were screened at a dilution of 1:80 for antibodies against B. caballi and T. equi using an IFAT, according to the manufacturer’s instructions (Laboratory Dr. Bose GmbH, Harsum, Germany). Each sample was run in duplicate. Ten microliters of diluted serum was placed in wells on masked slides, which contained fixed horse erythrocytes infected with B. caballi and T. equi. Positive and negative controls were supplied from the manufacturer. The slides were incubated in a humid chamber at +37°C for 30 minutes, successively rinsed, and soaked in buffer for 10 minutes.
Ten microliters of fluorescein-conjugated rabbit anti-horse IgG (Sigma Chemical, St Louis, MO) was applied to each well, and then incubation and rinse-soak steps were repeated. Finally, the slides were dried and mounted with 50% glycerol and 50% carbonate-bicarbonate buffer (pH 9). The slides were observed under an epifluorescent microscope (DM 2500; Leica, Wetzlar, Germany) using a xenon light source. A positive reaction appeared as peripheral clusters of distinct apple-green inclusion bodies within the infected erythrocytes (see Suppl. Fig. S1).
Histopathology and Immunohistochemistry
Muscle biopsies from the semitendinosus muscle were collected and snap frozen in isopentane precooled in liquid nitrogen.
Muscle samples from the same muscle of 5 healthy horses were used as control for immunohistochemical and molecular analysis. Specimens from both affected and control animals were frozen within 2 hours after sampling.
Frozen sections (10 μm thick) were subjected to a standard panel of histochemical stains, 54 including hematoxylin and eosin (HE), Engel’s trichrome (ET), NADH-tetrazolium reductase (NADH-TR), succinate dehydrogenase (SDH), cytochrome oxidase (COX), ATPase at pH 9.4 and 4.3, and periodic acid–Schiff (PAS) reaction.
A scoring system was designed to assess the degree of fiber atrophy as follows based on assessment of 100 fibers at 200× magnification: mild (score 1), <10% atrophic fibers; moderate (score 2), 10% to 50% atrophic fibers; and severe (score 3), >50% of atrophic fibers.
A scoring system was also defined for perivascular and/or endomysial lymphocytic inflammation based on light microscopy: no inflammation (score 0); mild inflammation (score 1), 5 to 25 lymphocytes/plasma cells per high-power field (HPF) (400×); and moderate inflammation (score 2), 26 to 50 lymphocytes/plasma cells per HPF.
At least 10 fields at 400× magnification were evaluated for each section by 2 independent pathologists (T.B.P. and O.P.) under an optical microscope (Nikon E600; Nikon, Tokyo, Japan), with a concordance rate of 95%.
For immunohistochemistry (IHC), frozen sections (8 μm thick) were processed with the MACH1 Universal HPR-Polymer Detection Kit (Biocare Medical LLC, Concord, CA). Briefly, the sections were dried for 1 hour at room temperature and fixed in acetone at 4°C for 3 minutes; peroxide block was applied for 15 minutes at room temperature, and then the sections were incubated for 30 minutes with background sniper (Biocare Medical LLC). The primary antibodies were diluted in phosphate-buffered saline (PBS) and incubated overnight at 4°C. MACH1 mouse probe was applied for 20 minutes at room temperature. Horseradish peroxidase (HRP)–polymer was added for 30 minutes at room temperature. After every step of the procedure, the sections were washed in 0.01 M PBS (pH 7.2–7.4). The reaction was revealed by using 3,3′-diaminobenzidine (DAB) chromogen diluted in DAB substrate buffer. Finally, sections were counterstained in Carazzi’s hematoxylin. Primary antibodies were directed against major histocompatibility complex I (H58A, mouse monoclonal antibody, dilution 1:200; VMRD, Pullman, WA), major histocompatibility complex II (H42A, mouse monoclonal antibody, 1:200; VMRD), CD3 (IS503, rabbit polyclonal antibody, 1:50; Dako, Milan, Italy), CD79α (HM57, mouse monoclonal antibody, 1:50; Dako), CD4 (HB61A, mouse monoclonal antibody, 1:50; VMRD), and CD8 (HT14A, mouse monoclonal antibody, 1:50; VMRD).
The percentage of muscle fibers with sarcolemmal positivity to major histocompatibility complex I and II (MHC I and I) was scored as follows: absent/none (score 0), 0%; mild (score 1), 1% to 25%; and moderate (score 2), 26% to 50%. For evaluation of MHC I and II, fibers directly adjacent to the inflammatory infiltrate were avoided, since positive endomysial inflammatory cells can make evaluation of sarcolemmal positivity difficult. The positive staining for MHC I and II varied from a continuous staining throughout the sarcolemma to a discontinuous sarcolemmal pattern.
Endomysial blood vessels, which are normally positive for MHC I and II, were used as positive internal controls for immunohistochemistry with anti–MHC I and MHC II antibodies. Frozen section of normal horse lymph nodes was used as positive controls for immunohistochemistry with anti-CD3, CD79α, CD4, and CD8.
The total inflammation score (IS) was obtained by summing the scores for lymphoplasmacytic inflammation, MHC I, and MHC II.
Indirect Immunofluorescence
To determine if IgG, IgM, or IgA autoantibodies directed against an unknown muscle antigen may be present in the bloodstream, we analyzed the serum from affected horses by using indirect immunofluorescence on sections of normal equine muscle (the same samples used as controls for histology and immunohistochemistry) as previously described in canine polymyositis. 35 Frozen sections (8 μm thick) were dried for 1 hour at room temperature and fixed in acetone at 4°C for 3 minutes. Protein block was performed using rabbit normal serum diluted 1:50 for 30 minutes at room temperature. Then, the sera from selected affected animals (horse Nos. 8, 10, 13, 14, and 15) were serially diluted in PBS (undiluted, 1:100, 1:300, 1:1000, 1:3000) and applied as primary antibody overnight at 4°C.
Slides were then washed 3 times in PBS and incubated with a fluorescein isothiocyanate (FITC)–conjugated rabbit anti-horse secondary antibody (equine IgG/IgM/IgA [FITC], SA 136092; Thermo Fisher Scientific, Rockford, IL) diluted 1:100 in PBS for 2 hours at room temperature. Slides were rinsed with PBS and mounted with a solution of 1 part glycerol/1 part PBS. For scanning and photography, a laser scanning microscope (LSM 510; Zeiss, Göttingen, Germany) was used with illumination at 488 nm and read using a 505- to 560-nm band pass filter.
As a negative control, serial sections of muscle were incubated with PBS or with serum from 2 healthy horses that were serologically negative for piroplasms, rather than serum from affected horses.
To determine if the presumed autoantibodies were specific for equine muscle, the same sera were tested on sections of normal muscle from dog, cat, and sheep.
Reverse Transcription Quantitative Polymerase Chain Reaction
Total RNA, isolated from skeletal muscle tissue from 10 affected animals (horse Nos. 1–3, 6–10, 12, and 15) and 3 controls, was extracted using TRIzol Reagent (Invitrogen Biotechnologies, Carlsbad, CA), according to the manufacturer’s instructions. Cases were selected based on the availability of frozen material for molecular analysis. Complementary DNA (cDNA) was synthesized using a reverse transcription kit (Maxima First Strand cDNA Synthesized Kit; Fermentas, Ontario, Canada) from 2 μg total RNA. Polymerase chain reactions (PCRs) were performed with a Bio-Rad CFX96 Connect Real-time PCR System instrument and software (Bio-Rad Laboratories, Hercules, CA). Primer and probe sequences used were as follows: tumor necrosis factor (TNF)–α (F: TTACCGAATGCCTTCCAGTC, R: GGGCTACAGGCTTGTCACTT), interferon-γ (IFNγ) (F: TGGACACCATCAAGGAGGAC, R: GGACCTTCAGATCATTTACCG), interleukin (IL)–10 (F: GTCATCGATTTCTGCCCTGT, R: GCTTCGTTCCCTAGGATGC), IL-12 (F: GACGCTGTGCCTTAGC, R: TCTGCCTCTGAGGATCTATCAACA), IL-4 (F: CAAAACGCTGAACAACCTCA, R: CTGTTGAAGCACCTTTGCAG), and IL-6 (F: AGCAAGTGTGAAAACAGAAG, R: CAT CAGGCAGGTCTCCTGAT) as previously described. 60,63 The PCR conditions were 10 minutes at 95°C followed by 40 cycles of 2-step PCR denaturation at 95°C for 15 seconds and annealing extension at 60°C for 60 seconds. Each sample contained 1 to 100 ng cDNA in 2× Power SYBRGreen PCR Master Mix (Applied Biosystems, Foster City, CA) and 200 nmol/l of each primer (Eurofins MWG Operon, Huntsville, AL) in a final volume of 25 μl. The relative amount of messenger RNA (mRNA) for each cytokine was normalized to GAPDH as a housekeeping gene, and the data were analyzed according to the 2–ΔΔCT method.
Reverse Transcription Quantitative PCR for T. equi and B. caballi DNA
Reverse transcription quantitative PCR (RT-qPCR) for T. equi 18S and B. caballi was performed on 11 of 16 frozen samples of affected horses (horse Nos. 1–4, 6–10, 12, and 15). Cases were selected based on the availability of frozen material for molecular analysis. Remaining 5 cases could not be evaluated due to lack of sufficient muscle tissue.
Muscle tissue (100 mg) was disrupted in 1 ml PBS using FastPrep1 FP120 Cell Disrupter Instrument (Qbiogene, Montreal, Quebec) in Lysing Matrix D tubes (MP Biomedicals, Santa Ana, CA). DNA was extracted from 200 μl of the homogenized tissue (QIAcube; Qiagen, Hilden, Germany) using the QIAampcador Pathogen Mini Kit according to the manufacturer’s instructions. Each sample was extracted in duplicate, and the average nucleic acid concentration was measured using a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE). 6
RT-qPCR for T. equi primers (F: Be18SF; R: Be18SR) and TaqMan probe (VIC-TAMRA, Be 18SP) were those previously reported 39 and amplified an 81-bp fragment outside the V4 hypervariable region of the 18S ribosomal RNA (rRNA) gene. For B. caballi, primers (F: Bc-18SF402; R: Bc-18SR496) and TaqMan MGBTM probe (FAM-MGB, Bc-18SP) were those reported 7 and amplified a 95-bp fragment in the V4 hypervariable region of the 18S rRNA gene of B. caballi. TaqMan Universal PCR Master Mix Kit (Applied Biosystems) was used, and the reactions were carried out using the ABI PRISM 7900 HT Sequence Detection System (Applied Biosystems). Internal positive controls were RT-qPCR products of B. caballi and T. equi, obtained from EDTA blood samples of seropositive symptomatic horses, certified by the Office International des Epizooties (OIE) Reference Laboratory for Babesiosis of the Istituto Zooprofilattico Sperimentale della Sicilia and cloned in the plasmid vector PCRII-TOPO (Invitrogen). The negative control was RNAase-free water.
Statistical Analysis
All data were imported into a program for statistical analysis (JMP 8.0; SAS Institute, Cary, NC). The level of significance was set at P ≤ .05. Data of cytokine mRNA levels are presented as mean ± standard error of the mean (SEM), whose statistical analysis was performed by analysis of variance test for multiple comparisons followed by Bonferroni’s test, using GraphPad Prism (GraphPad Software, San Diego, CA). Statistical significance was set at P < .05. For each animal, an inflammation score was obtained summing the scores of lymphocytic inflammation, MHC I, and MHC II. Data were checked for normality of distribution with a Shapiro-Wilk’s W test. The relationship between IS, histological score, and fiber atrophy was evaluated by a Spearman rank correlation test (rs).
Results
Parasitological Diagnosis, Hematology, and Serum Biochemistry
Signalment, clinical signs, selected serum biochemical findings, and serologic titers are provided in Table 1. By IFAT, 10 of 16 horses were positive for serum antibodies against both parasites, 3 of 16 were positive for B. caballi, and 3 of 16 were positive for T. equi (see Table 1).
Hematocrit was lower than 0.3 in 5 of 16 horses. The mean corpuscular volume (MCV) was lower than 45 fl in 2 of 16 horses. White blood cell (WBC) count was higher than 9.5 × 109/L in 2 of 16 horses, and in 11 of 16, platelets were lower than 1.6 × 1011/L. In 11 of 16 horses, β1-globulins were above the normal range (30–110 g/l), and albumin was reduced (<45 g/L) in 2 of 16 horses. In 3 of 16 horses, raised levels of serum bilirubin (>50 μmol/l) were observed. In 2 of 16 horses, hemoglobin (Hb) was lower than normal range (<68.3 μmol/L).
Serum activity of CK was mildly elevated in 10 of 16 horses, AST in 5 of 16, and LDH in 8 of 16 (Table 1), and SDH and γ-glutamyltransferase (γ-GT) were both elevated (>4 IU and 35 IU, respectively) in 1 of 16. Those minor serum biochemical abnormalities were consistent with a chronic form of equine piroplasmosis.
Histopathology and Immunohistochemistry
Pathological changes in muscle from all affected horses were similar, although they varied in severity (Table 2). In 11 of 16 cases (94%), the muscle contained small lymphocytes (Figs. 3, 4) with fewer macrophages (Fig. 5), plasma cells, and rare eosinophils.
Histologic Findings in the Muscle of in 16 Horses With Myopathy Associated With Chronic Piroplasmosis.
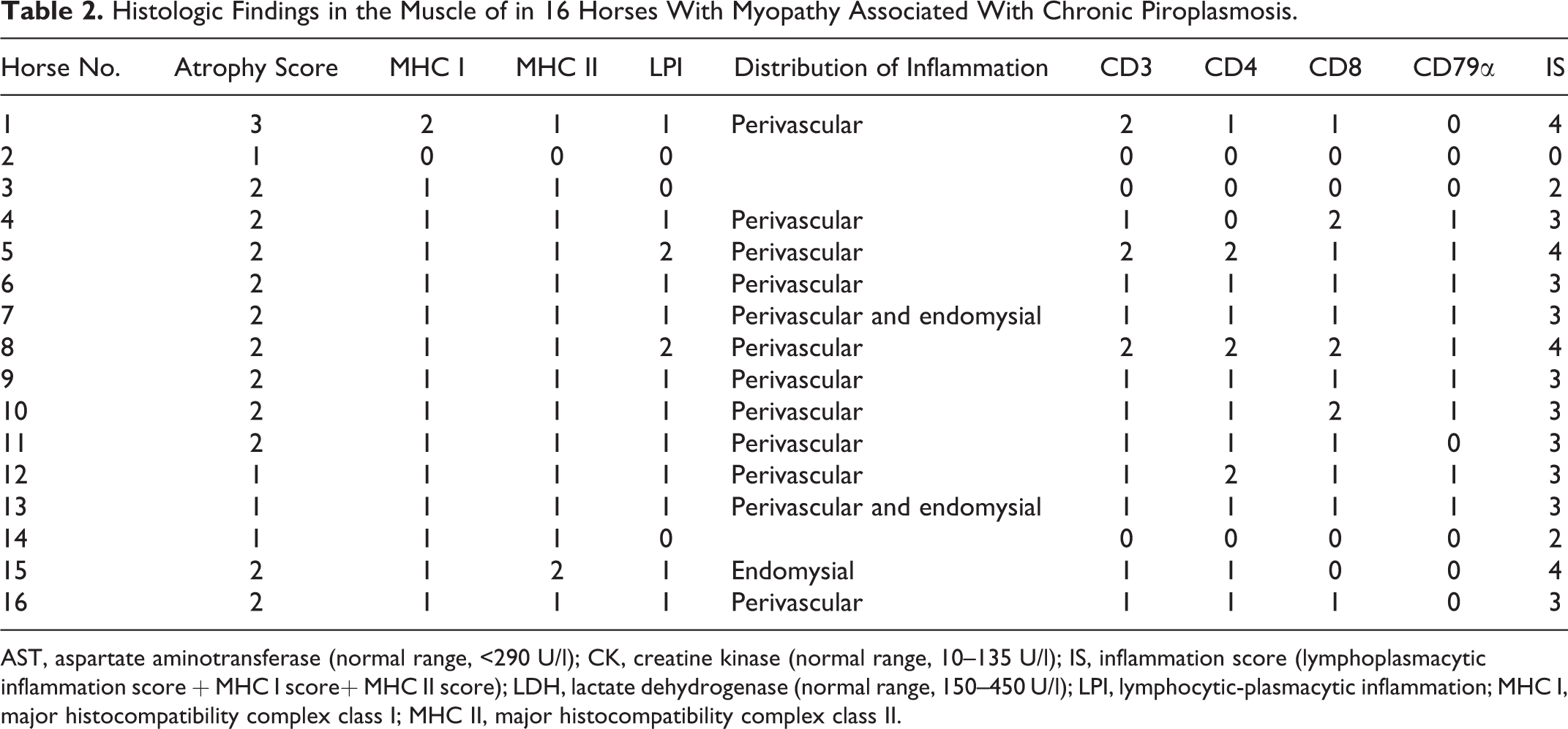
AST, aspartate aminotransferase (normal range, <290 U/l); CK, creatine kinase (normal range, 10–135 U/l); IS, inflammation score (lymphoplasmacytic inflammation score + MHC I score+ MHC II score); LDH, lactate dehydrogenase (normal range, 150–450 U/l); LPI, lymphocytic-plasmacytic inflammation; MHC I, major histocompatibility complex class I; MHC II, major histocompatibility complex class II.
In 12 of 16 cases, lymphocytic infiltrate formed cuffs around perimysial and endomysial blood vessels (perivasculitis; Fig. 4) with small numbers of lymphocytes scattered in the endomysium; 3 of 16 cases had mild lesions only in the endomysium. Only in 1 case with evident inflammation was perivascular cuffing not observed. In addition to lymphocytes, macrophages were also present in 5 of 16 cases (31%) (Fig. 5).
Various degrees of nonangular fiber atrophy was observed (Table 2), scored as mild in 4 of 16 cases (25%), moderate in 11 of 16 cases (69%), and severe in 1 case (6%). No fiber type selectivity was detected by ATPase staining at pH 9.4 and 4.3 (Fig. 6). Furthermore, mitochondrial abnormalities were identified using histoenzymatic stains in 13 of 16 cases (82%), including ragged blue fibers with SDH (Fig. 7) and fibers with a moth-eaten appearance with COX (Fig. 8) and NADH stains. Importantly, mitochondrial alterations were more prominent in muscle fibers adjacent to inflammatory infiltrates (Fig. 8). Additional morphological findings were as follows: necrotic fibers associated with sarcoclastosis (phagocytes invading necrotic fibers) in 11 of 16 cases (69%); hypereosinophilic, round, and swollen degenerated fibers in 12 of 16 cases (75%) (Fig. 4); and mild perimysial fibrosis in 5 of 16 cases (31%). No parasites or PAS-positive accumulations related to polysaccharide storage myopathy were found. The control biopsies did not contain histologic lesions or abnormalities in serum CK, LDH, or AST.
The immunophenotype of inflammatory cells in skeletal muscle was identified by immunohistochemistry. All cases contained CD3+ T-cell populations with CD4+ and CD8+ equally represented in 7 of 16 cases (43.7%; Figs. 9, 10). Only in 2 cases (horse Nos. 5 and 12) were CD4+ lymphocytes more abundant than CD8+. Scattered CD79α+ cells were present in 9 of 16 cases (56%), especially where the degree of inflammatory change was more severe. Lymphocytic infiltrate was graded as follows (Table 2): mild (score 1) in 11 of 16 (69%), severe (score 2) in 2 cases (13%), and absent (score 0) in 3 cases (19%).
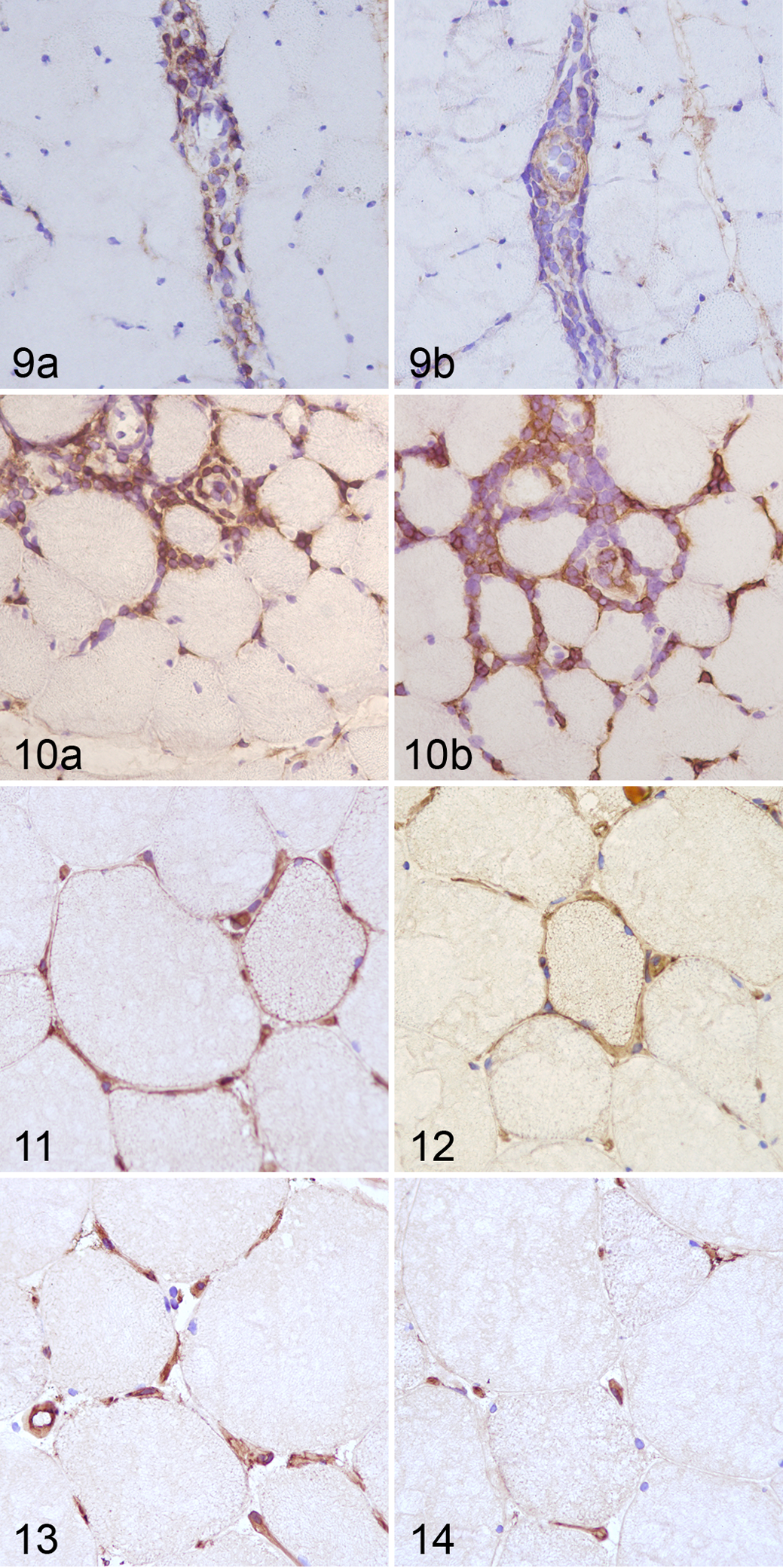
Furthermore, we evaluated the expression of MHC I and II antigens in myocytes. As expected, endothelial cells were positive in sections both from control and affected animals (Figs. 11–14) as well as many inflammatory cells (not shown). The majority of biopsies from affected animals (11/16 cases; 94%) showed a mild (score 1, 14/16 cases; 88%) to moderate (score 2, 2/16 cases; 13%) immunoreactivity to MHC I and II (Figs. 11, 12). The staining pattern was localized to the sarcolemma or the cytoplasm (sarcoplasm) in both the atrophic and normal-sized fibers. The staining pattern varied from a distinct, continuous sarcolemmal positivity, to discontinuous sarcolemmal positivity, to sarcoplasmic positive spots. Positive muscle fibers were even found distant from the inflammatory infiltrate. In control cases, MHC I and II expression was restricted to endothelial cells (Figs. 13, 14).
The histopathologic data were analyzed, and a statistically significant correlation between inflammation and atrophy scores was found (rs = 0.6; P = .02, Spearman rank correlation test). The atrophy and MHC I scores were also correlated (rs = 0.6; P = .008). No correlation was found between inflammation score and serum levels of muscular enzymes (CK, P = .24; LDH, P = .8; AST, P = .53) or between inflammation score and serum titers to etiological agents (T. equi, P = .09; B. caballi, P = .86; T. equi and B. caballi, P = .22).
Indirect Immunofluorescence
To detect muscle-specific antibodies in serum of affected horses, serum was applied to sections of muscle and then probed with antibody against equine IgG, IgM, and IgA. Distinct sarcolemmal labeling was detected in all sections subjected to serum from 4 affected animals even at high dilution (1:3000) of the conjugate (Fig. 15a). Sections incubated with serially diluted serum from normal control horses did not show the same dilution-dependent fluorescence pattern (Fig. 15b). Weak sarcolemmal staining was detected in sections incubated with serum from normal control horses and interpreted as nonspecific background, because its intensity did not vary significantly at different dilutions, whereas the labeling was dilution dependent for sections incubated with serum from affected horses. No staining was observed in sections incubated with PBS rather than equine serum (Fig. 15a,b) or sections treated with PBS rather than FITC-conjugated secondary antibody (not shown).
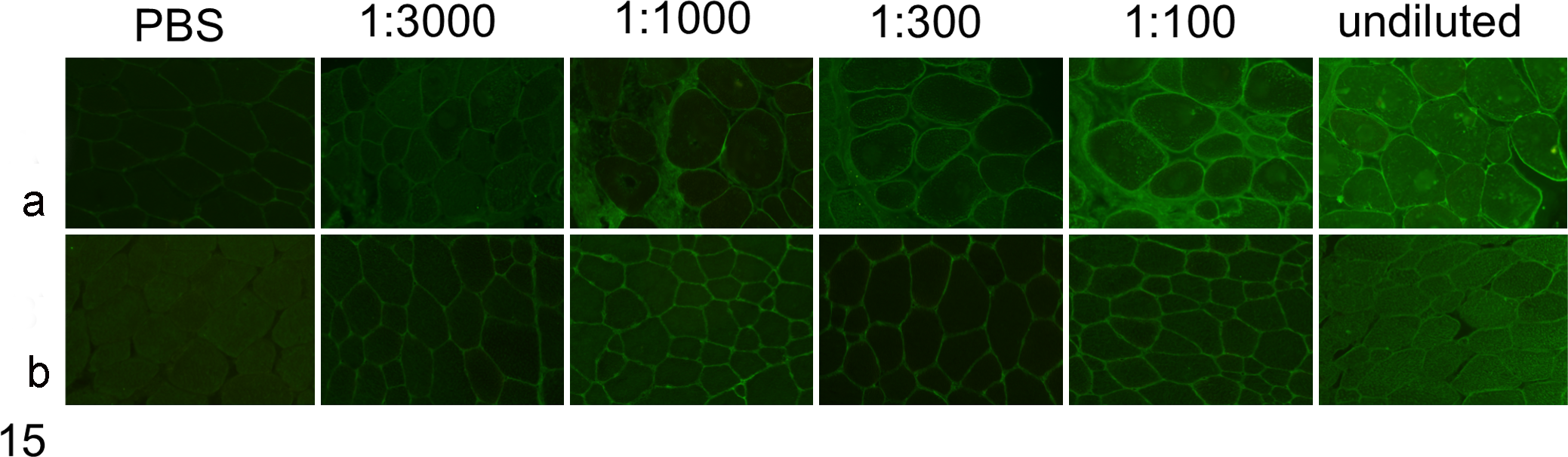
Indirect immunofluorescence to detect muscle-reactive antibodies in horse serum. Normal equine muscle cryosections were incubated with undiluted and serially diluted horse serum. (a) Strong dilution-dependent, sarcolemmal positivity is evident in sections treated with serum from the affected horse (horse No. 14). (b) In the section treated with serum from a control normal horse, sarcolemmal staining is weak, not dilution dependent, and considered nonspecific. Sections treated with phosphate-buffered saline (PBS) rather than serum are negative.
To investigate species specificity of the target antigens, equine serum was applied to sections of skeletal muscle from dog, cat, and sheep and detected as above. Using serum from affected horses, antibodies bound to dog, cat, and sheep skeletal muscle cryosections, with similar staining intensity (Fig. 16a). In contrast, normal control horse serum showed no immunolabeling of muscle from these species (Fig. 16b).
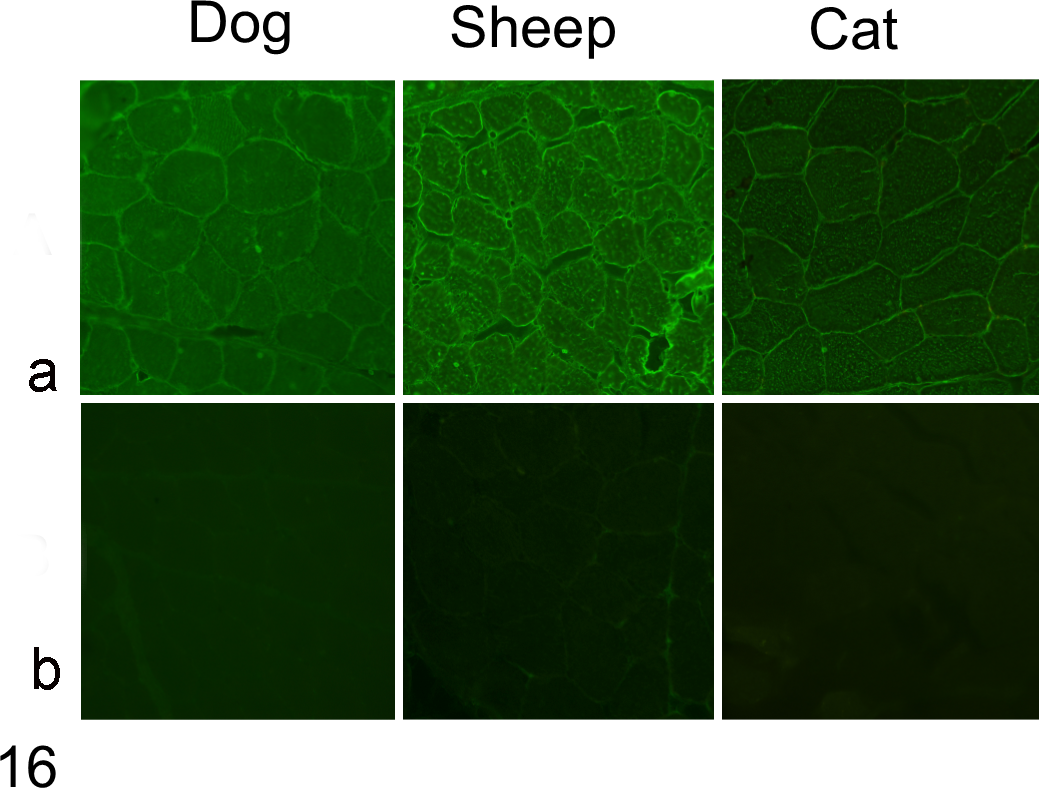
Indirect immunofluorescence to detect reactivity of antibody in horse serum to normal canine, feline, and sheep muscle. Serum-probed cryosections were labeled with FITC-conjugated rabbit anti-horse IgG, IgM, and IgA. (a) Strong sarcolemmal reactivity is evident in the section labeled with serum from horse No. 14 diluted 1:300. (b) The section treated with serum from a normal healthy horse is negative.
Molecular Findings
Using RT-qPCR, significantly increased mRNA levels of IL-12, TNF-α, and IFN-γ were found in muscle samples from affected animals compared to controls, while IL-10 mRNA levels were not different between affected and control animals (Fig. 17). No amplification of IL-4 and IL-6 was found either in specimens from affected animals or controls.
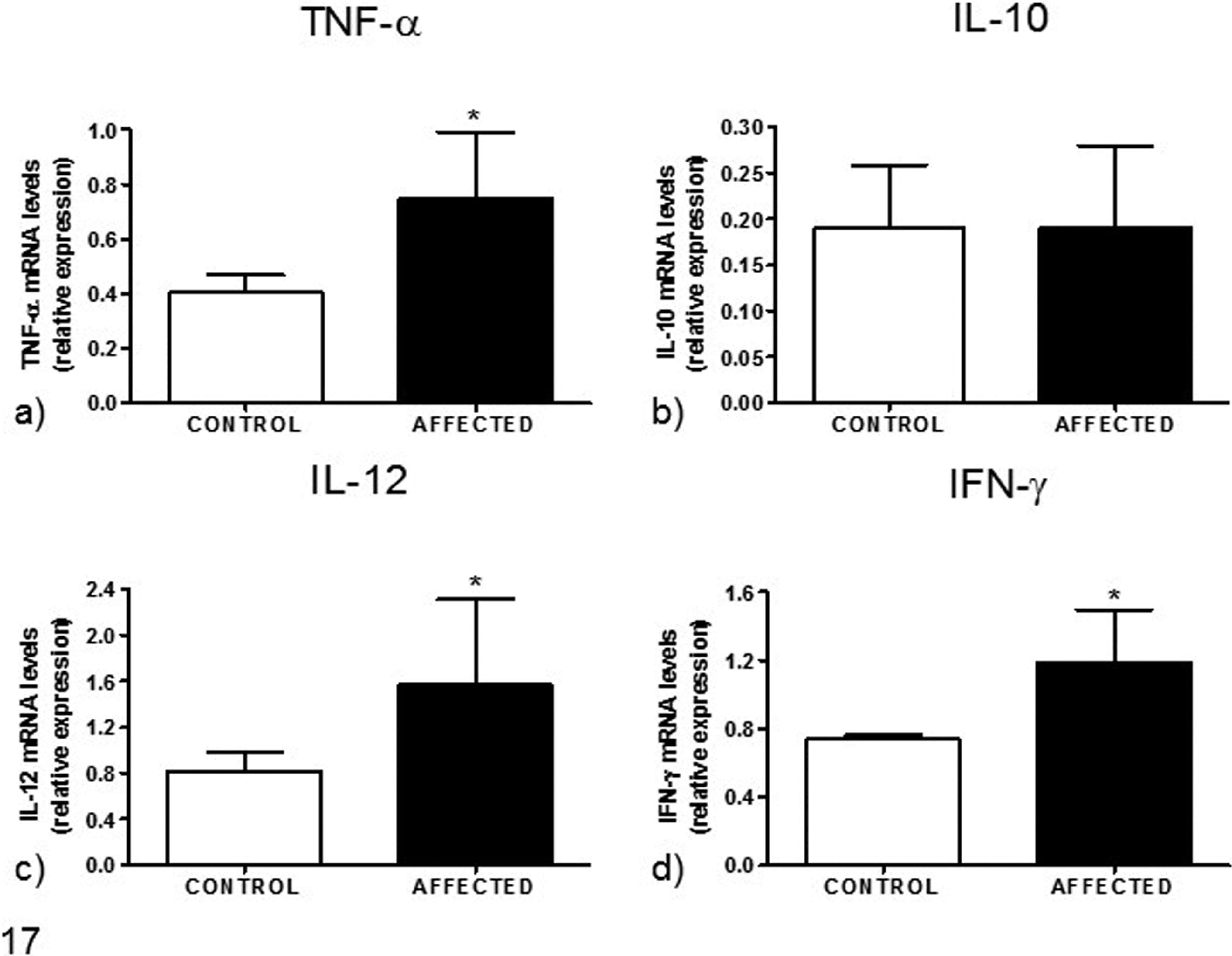
Analysis of (a) tumor necrosis factor (TNF)–α, (b) interleukin (IL)–10, (c) IL-12, and (d) interferon-γ (IFN-γ) gene expression based on reverse transcription quantitative polymerase chain reaction, from skeletal muscle of horses with myositis associated with chronic piroplasmosis compared to healthy controls. Data are mean ± SD of 5 samples per group. *P < .05 vs control. mRNA, messenger RNA.
T. equi and B. caballi DNA was not detected by RT-qPCR in any of the 11 muscle samples tested from horses with chronic piroplasmosis.
Discussion
Although clinical evidence of muscle impairment has been reported in chronic equine piroplasmosis, 69 the histopathological and immunological features have not been documented to date. This study provides the first evidence of muscle pathology in horses chronically affected by piroplasmosis and with a clinical history of muscle atrophy and poor performance.
Our results suggest that in chronic equine piroplasmosis, functionally important muscle damage may depend on an immune-mediated muscle damage involving circulating autoantibodies, and local cytokine expression may contribute to impaired muscle function. If so, this common protozoal disease should be included in the differential diagnosis of polymyositis and poor performance in horses as well as one of the possible causes underlying equine immune-mediated myopathy. Our work highlights the importance of the histopathological evaluation of muscle biopsies as a starting point for a more precise understanding of the pathogenesis of clinical signs of muscle impairment.
To date, histopathological features and pathogenetic aspects of idiopathic inflammatory myopathies have been extensively studied, especially in humans and dogs. 15,22,27,30,45,58,68 In humans, the main idiopathic inflammatory myopathies are polymyositis (PM), dermatomyositis (DM), immune-mediated necrotizing myopathy (NM), and sporadic inclusion body myositis (sIBM). 15 The disorders have primarily an autoimmune pathogenesis, mediated either by cytotoxic T cells, as in polymyositis and inclusion body myositis; by a complement-mediated microangiopathy, as in DM (dermatomyositis); or by macrophages and possibly autoantibodies, as in NM (necrotizing myopathy). 15 In dogs, the best-known immune-mediated myopathies are masticatory muscle myositis, 70 polymyositis, dermatomyositis, extraocular muscles myositis, 37 and inflammatory myopathy associated with Leishmania infantum infection. 53 Masticatory muscle myositis and extraocular muscle myositis involve an immune response directed against unique antigens of this muscle group, while polymyositis and dermatomyositis share many pathogenetic aspects with their human counterparts. 22,58 Systemic infectious diseases have been proposed as triggering factors of inflammatory myopathies and cardiomyopathies in humans, dogs, and hamsters. 11,15,22,36,56 At least 3 main mechanisms have been proposed to explain the role of infectious factors as a trigger for autoimmune disease involving skeletal muscles: (1) polyclonal B- or T-cell activation, (2) molecular mimicry, or (3) immunogenicity of muscle auto-antigens secondary to infection-mediated inflammation. 3,26,62
Based on anamnestic and histopathological data, we described an equine inflammatory myopathy that seems to involve an immune-mediated pathogenesis. A similar pattern of cellular infiltration has been described in equine immune-mediated myositis, 20,43 with the only exception that CD4+ cells were the predominant lymphocyte in immune-mediated myositis but were rare in the myopathy associated with chronic piroplasmosis (only 2 of 16 cases). Given the perivascular pattern of the mixed T-lymphocytic infiltrate (CD4+ and CD8+ cells), we cannot exclude an immune reaction against blood vessels as the primary target as in human and canine dermatomyositis. 3,26 However, the absence of a true vasculitis (ie, inflammatory cells invading and causing damage to vessel walls) and binding of serum antibodies to myocytes identified by indirect immunofluorescence are findings that argue against an immune reaction against blood vessels. A more likely explanation for the inflammatory myopathy caused by piroplasmosis seems to be an immune reaction to a protozoal antigen priming molecular mimicry, with subsequent development of autoantibodies against muscle antigens that result in damage to myocytes. The sarcolemmal upregulation of MHC I, the presence of cytotoxic CD8+ T lymphocytes that are the histological hallmark of human and canine PM (polimyositis), 15,16,22 and the presence of sarcolemmal staining following incubation with infected horse sera all support this hypothesis. Moreover, the similar findings when serum of infected horses was incubated with muscle tissue from 3 other species (dog, cat, and sheep) make it likely that such autoantibodies are not species specific, resembling autoantibodies in canine PM. 35 In this study, we did not examine binding of serum antibodies to other normal tissues to determine if equine piroplasmosis autoantibodies are muscle specific.
Regarding the immunohistochemical detection of sarcolemmal MHC I and II, it is noteworthy that MHC upregulation has been found in immune-mediated idiopathic myositis. The diagnostic value of detection of sarcolemmal MHC I and II in human and canine inflammatory myopathies 54,55,65 has been recently confirmed in equine immune-mediated myositis. 20 Upregulation of MHC I and II has been correlated to the active role of muscle fibers in antigen presentation and in initiating and maintaining pathological events in myositis independently of inflammatory infiltrates. 21,55 It is conceivable to speculate that during chronic piroplasmosis, an unknown stimulus could activate a series of inflammatory pathways resulting in sarcolemmal overexpression of MHC I and II, which in turn would attract T lymphocytes that exacerbate and perpetuate the inflammatory myopathy.
RT-qPCR for direct identification of piroplasmal DNA in skeletal muscle gave negative results in our study. Like in other immune-mediated myopathies, 15 chronic infections do not cause muscle damage through a direct replication of the agent in the muscle but instead trigger a persistent inflammatory response with activated T cells. Indeed, in humans and monkeys infected with human immunodeficiency virus that develop PM or sIBM (sporadic Inclusion Body Myositis), sensitive PCR studies have repeatedly failed to confirm the presence of the virus in muscle. 13,14,42 Further studies are needed to definitely establish a direct link between chronic piroplasmosis and immune-mediated myositis, such as the identification of clonally activated T lymphocytes stimulated by piroplasm antigens.
For the inflammatory cytokines IL-12, TNF-α, and IFN-γ, the levels of gene expression within muscle were increased in affected compared to control animals. IL-12 is a proinflammatory cytokine produced by both immune and nonimmune cells 49,64 that play a key role in autoimmune diseases and food allergies. 64 Indeed, it stimulates the production of IFN-γ and TNF-α from Th1 cells and natural killer (NK) cells, respectively. TNF-α (also named cachectin) is involved in the pathogenesis of muscle wasting in human sarcopenia and cachexia. 33 TNF-α effects are mediated by the transcription factor nuclear factor (NF)–κB, which upregulates the ubiquitin-proteasome pathway and increases the expression of inducible nitric oxide synthase (iNOS), leading to oxidative stress 33,44 and decreased protein content in muscle fibers. In horses, TNF-α was demonstrated to stimulate granulocyte diapedesis and their adherence to the endothelium. 4 Notably, TNF-α is able to antagonize the biological effects of insulin-like growth factor 1, 25 thus contributing to the development of muscle atrophy. TNF-α has a synergistic role with IFN-γ in inducing muscle atrophy in human muscle cell culture and animal models of cachexia. 38,46 TNF-α and IFN-γ are also considered strong inducers of MHC I antigens on muscle fibers. 15 Furthermore, previous studies demonstrate that cytokine stimulation induces muscle fibers to also secrete proinflammatory cytokines, such as IFN-γ, in an auto-amplificatory mechanism that may facilitate the recruitment of activated T cells contributing to the self-sustaining nature of myositis and disease chronicity. 23,45 TNF-α, IFN-γ, and IL-12 are indicative of a typical Th1-mediated response, 40 and their involvement in the immune response in equine piroplasmosis 34 could explain the prominent muscle atrophy in our cases considering their aforementioned biological function.
Furthermore, a statistically significant correlation between inflammation score and muscle atrophy was found, suggesting a common pathogenic mechanism related to the chronic release of cytokines such as TNF-α or IFN-γ. Cytokines have been demonstrated to be a promising immunotherapeutic target. 17,51
Mitochondrial abnormalities are frequently associated with muscle inflammation, as reported in several human inflammatory myopathies where they are generally associated with damage to the mitochondrial DNA. 52,67 In 82% of muscle biopsies from affected horses, prominent mitochondrial histochemical alterations were found, possibly contributing to clinical muscle impairment.
In conclusion, we described a clinically significant inflammatory myopathy associated with chronic equine piroplasmosis. The condition is characterized by infiltration of CD4+ and CD8+ T lymphocytes, evidence of binding of serum antibodies to muscle antigens, and increased gene expression of inflammatory cytokines, suggesting a likely immune-mediated pathogenesis.
Footnotes
Acknowledgements
We gratefully thank Raffaele Ilsami for his technical support in histology.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.