
Abstract
H5N1 highly pathogenic avian influenza (HPAI) virus causes high mortality of infected birds, with infection in multiple organs, including in feathers. Feathers have been proposed as samples for diagnosis of HPAI infection in birds, and this study is part of a broader investigation validating the use of feathers for diagnostic purposes. To understand and characterize the morphological basis for feather infection, sections from 7 different skin tracts of ducks and chickens infected with 3 different clades of H5N1 HPAI virus from Indonesia and Vietnam were examined histologically. Results showed that in ducks, lesions and viral antigen were mainly detected in the epidermis of feathers and follicles, whereas in chickens, they were mostly found in the dermis of these structures. Abundant viral antigen was found in nearly all the feathers examined from chickens, and there was no apparent difference between virus isolates or skin tracts in the proportion of feathers that were antigen positive. By immunohistochemistry, the majority of feathers from most skin tracts from ducks infected with a Vietnamese H5N1 HPAI virus contained abundant levels of viral antigen, while few feathers were antigen positive from ducks infected with 2 Indonesian viruses. These results support and inform the use of feathers for diagnostic detection of H5N1 HPAI virus in birds.
Since its emergence in 1996 in Guangdong province, China, 17 H5N1 highly pathogenic avian influenza (HPAI) virus has spread and circulated widely around the world and has caused significant losses to affected countries. 4 In addition, several new reassortant HPAI viruses have appeared in Asian poultry populations, a concerning trend indicating their active evolution and continuous threat to poultry and human populations. 5,16,22,23
HPAI viruses produce a systemic infection in chickens, affecting multiple organs, including the cardiovascular, nervous, lymphoid, and integumentary systems, leading to severe disease. 13 The outcome of HPAI infection in ducks is variable, ranging from asymptomatic infection to severe fatal disease, and is dependent on viral strain and age. 2,9,14 The different clinical picture observed between the 2 species reflects the different tissue tropism and pathology of HPAI viruses in chickens and ducks. In chickens, H5N1 HPAI has a marked tropism for endothelial cells, with variable spread to other cell types, 7,10,12,14 and it is the endothelial tropism that is thought to account for the severe acute disease in that species. In ducks, viral antigen occurs in a wide range of tissues, including myocardium and skeletal muscle, brain, and epithelial surfaces of the respiratory tract, but rarely in endothelium. 2,6,14 Variability of infection in ducks appears to be related to amount of antigen, rather than in differences of tropism.
Histological lesions and antigen in feather structures have been noted, 20 and our group previously reported necrosis and hemorrhage in the feather pulp of chickens infected with Indonesian HPAI viruses, along with immunohistochemistry (IHC) staining of endothelial cells in that tissue. 14 In ducks in the same study, we detected antigen in feather epidermis and feather pulp, and in ducks infected with a Vietnamese HPAI virus, we reported the presence of antigen in feather follicle epithelium and feather pulp, often associated with necrosis of these tissues. 2
The detection of HPAI viruses in feathers of chickens, ducks, and other avian species infected with HPAI viruses 2,3,14,19 –21 led to suggestions that feathers could be a potential sample for diagnosis of HPAI infection. Oropharyngeal and cloacal swabs are the recommended samples for detection of HPAI viruses and have been used extensively for both diagnostic and surveillance purposes. Virus detection in swabs is performed by various virological methods, including virus isolation, reverse transcriptase polymerase chain reaction (PCR) (conventional and real time), and rapid antigen detection tests. Feathers provide an alternative sample for HPAI diagnosis and have been shown to have higher viral titers than either swab type in both chickens and ducks. 8 Considering the low levels of virus-positive samples that are obtained from ducks infected with some strains of HPAI virus, our previous study showed that feathers offer an important additional sample type that can help to maximize the likelihood of determining the true infection status of an individual duck, particularly in the later stages of infection. 8
Although there have been a number of studies on the use of feathers for diagnosis of HPAI infection, 3,8,19 –21 there have been no comprehensive studies examining the distribution of virus in feather structures of different skin tracts of chickens or ducks, which could determine the most appropriate region of the body from which to collect diagnostic samples in these species. There have also been limited detailed studies on the cellular distribution of viral antigen within the feather, feather follicle, and skin. Finally, although previous studies have used virological methods to detect HPAI virus in feather samples, 3,8,19 –21 none have assessed histological methods as an approach for HPAI diagnosis. To address these deficiencies in the literature, we conducted a comprehensive histological study of H5N1 HPAI virus in skin of experimentally infected ducks and chickens.
Materials and Methods
H5N1 HPAI Virus Strains
Two Indonesian isolates of highly pathogenic avian influenza H5N1 virus were used: A/duck/Sleman/BBVW-1003-34368/2007 (IDN 34368, clade 2.1.1) (GenBank reference CY091949) and A/duck/Sleman/BBVW-598-32226/2007 (IDN 32226, clade 2.1.3) (GenBank reference CY091864). These isolates were collected from ducks in Central Java Province, Indonesia, during active surveillance in a collaborative project between the Indonesian Ministry of Agriculture and the Australian Centre for International Agriculture Research (ACIAR). 15 The infectivity and pathogenesis of these viruses have been reported elsewhere, where it was found that both cause severe peracute disease in chickens and are generally asymptomatic in ducks. 14 The Vietnamese isolate was A/Muscovy duck/Vietnam/453/2004 (VN 453), and this virus was previously reported to cause disease in ducks and chickens. 2,8
Animal Trial Protocol
The samples used in this study were taken from birds used in other experiments, and therefore the trial protocols were predicated on those studies. The animal infection studies for the Indonesian viruses have been described previously. 14 Feather samples and oropharyngeal and cloacal swabs were collected from each of the birds and analyzed for virus, as documented elsewhere. 8 For the study described here, feathered skin samples from the same birds were collected into formalin for additional histopathological analysis. To ensure that the animal experiments conformed to national animal welfare legislation, all protocols implemented were approved by the AAHL Animal Ethics Committee. We intended to euthanize birds if they reached moderate clinical signs (huddling together, fluffed-up feathers, and reluctance to move when disturbed) before the scheduled sampling time point (outlined below); however, due to the rapid progression of the infection, particularly in chickens, this was not always possible, leading to deaths in some birds.
Briefly, the 7-week-old Pekin ducks (Anas platyrhynchos) and 3-week-old broiler chickens (Gallus domesticus) used in the study were both sourced from commercial farms and were clinically healthy and sero-negative. The virus was suspended in phosphate-buffered saline and given in a 0.5-mL equivalent to 107.4, 108.4, and 105.7 median egg infectious doses (EID50) per duck and in a 0.2-mL equivalent to 107.0, 108.0, and 105.3 EID50 per chicken for isolates IDN 34368, IDN 32226, and VN 453, respectively. For all birds, 1 drop of the viral suspension was administered onto each eye and into each nostril, and the remainder was released into the mouth. Fourteen ducks and chickens were inoculated with IDN 34368, 13 ducks and chickens with IDN 32226, and 5 of each species with VN 453.
In the current study, samples from a total of 14 chickens and 30 ducks were used. Three randomly selected chickens were euthanized and sampled from both of the Indonesian virus groups at 24 hours postinoculation (hpi); an additional 3 birds were sampled at 30 hpi only from the IDN 34368 group, as by then all the IDN 32226 chickens had died. All 5 VN 453–infected chickens were euthanized and sampled at 24 hpi. In ducks, samples were collected from 14 and 12 birds in the IDN 34368 and IDN 32226 virus groups, respectively, at designated times of 2, 4, 7, and 18 days postinoculation (dpi), or if clinical signs warranted euthanasia. Four ducks in the VN 453 group were euthanized and sampled at various times postinoculation on the development of moderate clinical disease (reluctance to move on disturbance, recumbency).
Skin Samples
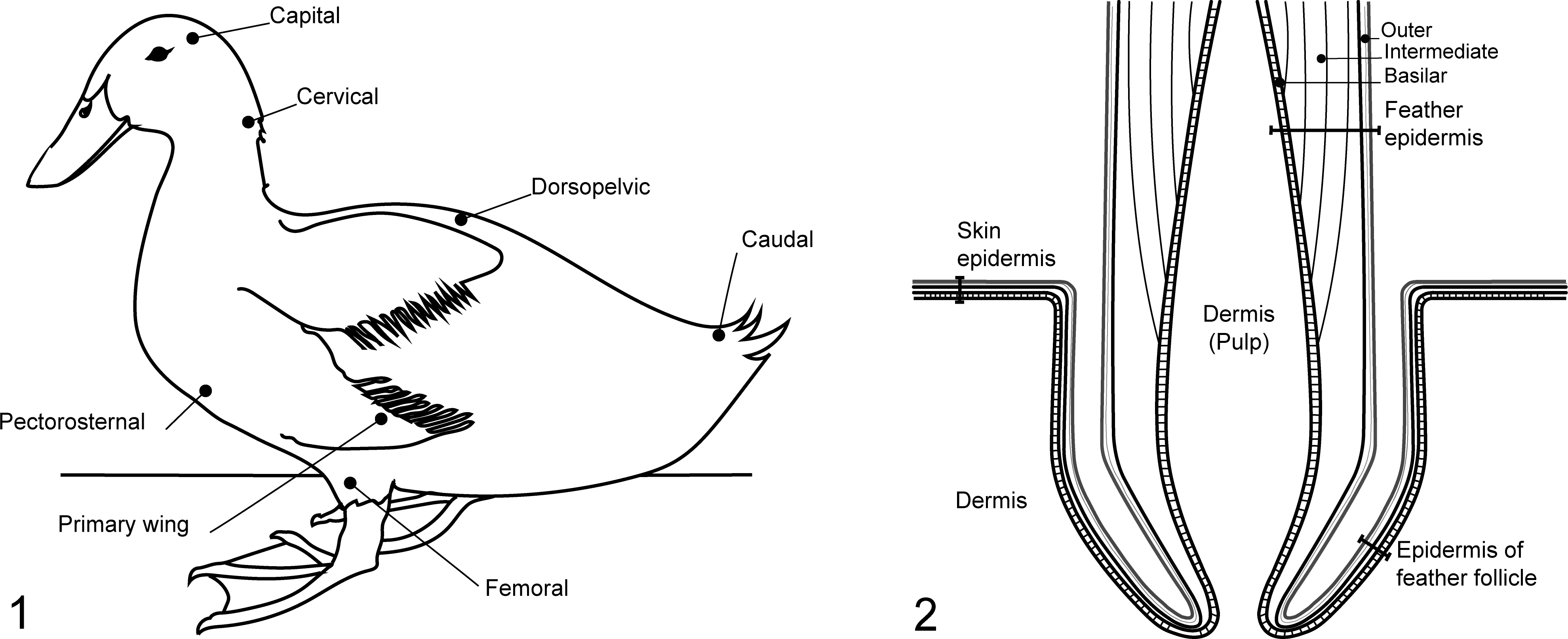
Samples were taken from 5 skin tracts (capital, cervical, dorsopelvic, pectorosternal, and femoral) containing typical contour feathers, as well as primary wing tracts (containing remiges) and caudal tracts (containing retrices) (Fig. 1). Pieces of skin approximately 3 to 4 cm in diameter, with feathers, were removed from the carcass and placed into 10% neutral phosphate-buffered formalin for fixation. Samples were fixed for 2 to 3 days before histological processing.
Histology and Immunohistochemistry
Seven skin tracts were processed from each bird. Skin samples were trimmed such that the longitudinal aspect of feather follicles was sectioned. Excess feather vanes 2 to 3 mm over the surface of the skin were trimmed off. The samples were processed and embedded into wax by routine histological processes. One section of each tract was cut by standard microtomy methods, and consecutive 4-μm-thick sections were stained with hematoxylin and eosin (HE) or by immunohistochemistry (IHC) techniques. The latter method has been described in detail in a previous report. 14 Briefly, the primary antibody used for the IHC was a rabbit antiserum raised against purified recombinant influenza A nucleoprotein expressed in Escherichia coli and was used at a dilution of 1:400. Antigen was visualized by horseradish peroxidase–conjugated secondary polymer antibody and aminoethylcarbamide chromogen (Envision Flex-HRP; Dako, Glostrup, Denmark). Sections were counterstained with hematoxylin. Sections of uninfected chickens and ducks were tested using the IHC, and no nonviral staining was found in these sections (Suppl. Fig. S1).
The IHC method was used to determine the proportion of virus-positive feathers by counting virus-positive feathers (feathers with viral antigen observed in any cell or structure) and total feathers in each IHC-stained section. This was assessed in each of 7 skin tracts (as above, Fig. 1) from a total of 14 chickens and 31 ducks. One IHC section was examined per tract, and in these sections, viral staining was assessed in the various layers of the feather, follicle, and skin (Fig. 2). Between 1 and 64 and 1 and 159 feathers per tract were examined for chickens and ducks, respectively. The range in feather numbers examined per tract was due to differences in the size of feathers: the primary tracts had only few large feathers per section, while the capital and cervical tracts had many small feathers.
Statistical Analysis
Within each virus-inoculated group of chickens or ducks, the mean tract percentage of virus-positive feathers per bird was compared statistically for each infection group using nonparametric repeated-measures 1-way analysis of variance (ANOVA) and Friedman’s test followed by Dunn’s multiple-comparison test. These methods were used because feathered skin tracts were collected from the same (dependent) bird, while we also compared samples from different (independent) birds. This analysis was carried out using GraphPad Prism 5 software (GraphPad Software, San Diego, CA).
Results
Clinical Outcomes
The clinical and pathological outcomes of infection for the Indonesian virus trials have been described elsewhere, 14 and the clinical and pathological signs for the Vietnamese virus in ducks were similar to those described elsewhere for this virus. 2 All inoculated chickens developed clinical signs and were euthanized or died acutely. The IDN 32226– and VN 453–infected chickens were all euthanized by 24 hpi. Three chickens infected with IDN 34368 were euthanized and sampled with mild clinical signs at 24 hpi; all other chickens in the group were euthanized or died with definite clinical signs at 30 hpi. Apart from 1 death in the IDN 34368–inoculated group (duck No. 9), only the ducks inoculated with the Vietnamese H5N1 HPAI virus showed clinical signs. Four of 5 ducks infected with the Vietnamese virus developed disease and were euthanized for welfare reasons, and the remaining duck was euthanized on day 10; this latter duck was not sampled for this study. One duck (No. 27), challenged with IDN32226 and euthanized on 7 dpi, did not have evidence of infection 14 and was therefore not included in the data set. All other birds in the study either seroconverted or yielded at least 1 positive sample by virus isolation.
Lesions in the Integument Caused by HPAI Virus Infection in Chickens and Ducks
All feathers observed in chicken skin sections were growing (immature) with highly vascularized pulp. In chickens, lesions in infected feathers consisted of acute focal necrosis mainly in the feather pulp (Fig. 3A) but also in the dermis of feather follicles and skin. No inflammatory or other tissue responses were observed. Lesions were associated with antigen staining by IHC (Fig. 3B). The type and prevalence of lesions were similar for chickens infected with all 3 H5N1 HPAI viruses.
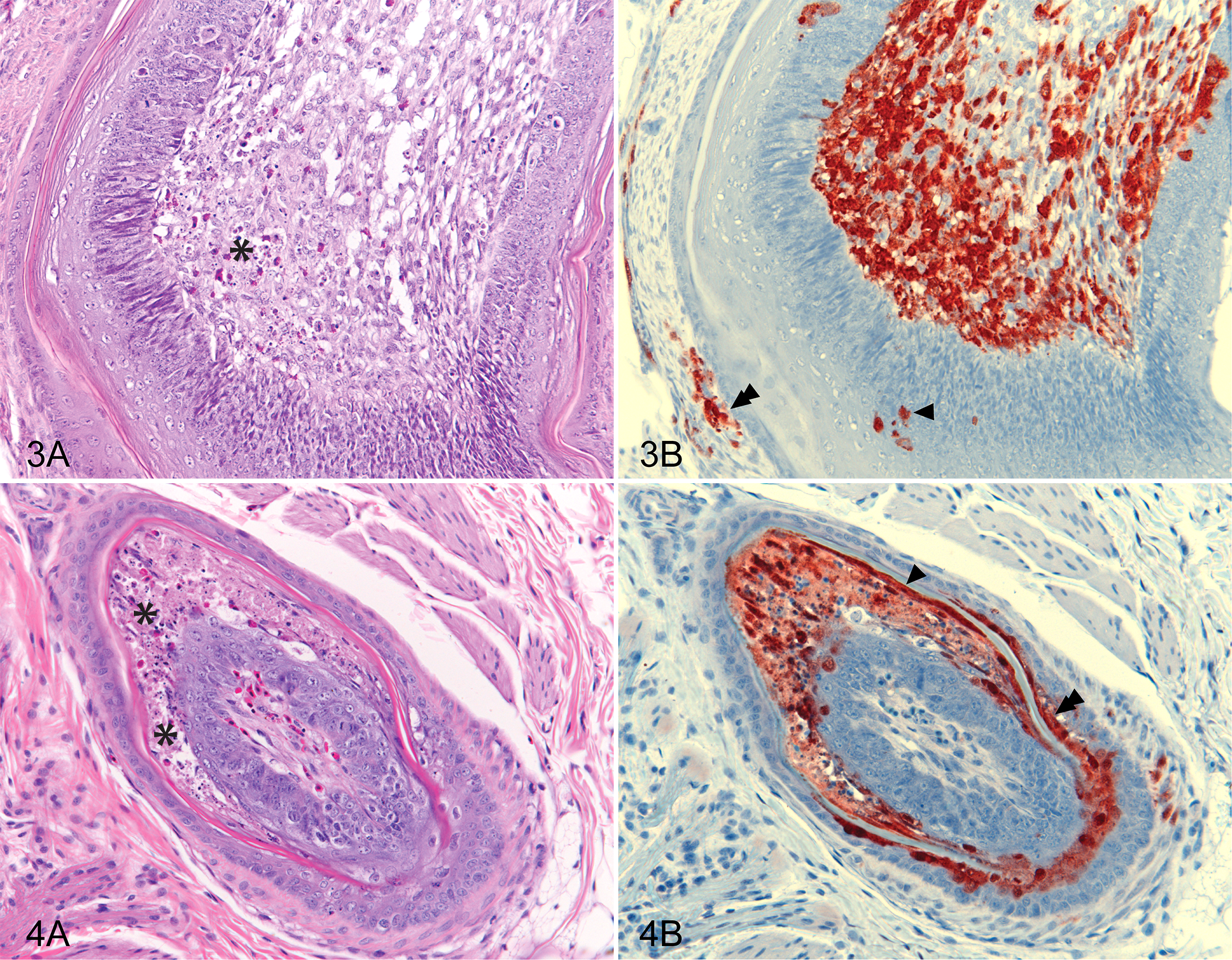
In duck skin sections, there was a mixture of immature and mature feathers. Lesions, where they occurred, consisted of focal necrosis, occurring mainly in the epithelium of the immature feathers and follicles (Fig. 4A). Usually, there were no associated inflammatory responses. Such lesions were associated with viral antigen on IHC. Major differences between viruses were detected in ducks, with frequent and severe lesions occurring in ducks infected with VN 453, while in ducks infected with either of the Indonesian H5N1 HPAI viruses (IDN 34368 and IDN 32226), viral lesions and antigen were detected rarely.
Distribution of Viral Antigen
The distribution of viral antigen in feathers, feather follicles, and skin was consistent across all the chicken samples examined. Viral nucleoprotein antigen was mainly detected in dermal structures (including feather pulp), and less antigen was found in the epidermal structures, the latter being confined to the intermediate and basilar layers of feather epidermis (see Fig. 2 for a schematic diagram of the integument structures and cell layers). Viral antigen was detected mainly in the endothelial cells of the small vessels of the dermis and dermal structures and in the tissues immediately surrounding capillaries. Therefore, antigen was particularly dense in the highly vascularized tissues of the pulp of the immature feathers (Fig. 3B) but also associated with capillaries throughout the dermis.
The distribution of viral antigen in the duck skin sections showed a different pattern from that in the chicken. Abundant viral antigen was observed in the outer and intermediate layers of the feather epidermis (Fig. 4B), with less antigen detected in feather pulp and basilar layer of the feather epidermis. Antigen was also found in the epidermis of feather follicles. No viral antigen was detected in the epidermis of skin, and viral antigen was present, but generally rare, in the dermis. In this species, the antigen quantity varied in some tissue structures (such as pulp and basilar layer) far more than appeared to be the case in chickens, even in the same infection group.
In all chickens of all groups, 100% of feathers from all but the capital tract were viral antigen positive. In the capital tract, 62.7% of feathers were positive in early disease (24 hpi, IDN 34368), while 93.3% to 99.5% of feathers from this tract were positive during clinical disease (Table 1, Suppl. Table S1). There was no significant difference in the mean percentage of antigen-positive feathers in all tracts evaluated for all virus groups (P > .05) (Suppl. Table S1).
Summary of the Proportion of Viral Antigen-Positive Feathers in Skin Sections From 7 Different Regions of Skin (Tracts) in Chickens Inoculated With IDN 34368, IDN 32226, or VN 453 H5N1 HPAI Viruses.
Skin sections were stained by immunohistochemistry to detect influenza A nucleoprotein. Sections from all tracts of all individual birds contained at least 1 viral antigen-positive feather. No significant differences were detected in the mean percentage of antigen-positive feathers between any of the skin tracts within each virus group. See Supplemental Table S1 for individual bird data. EID50, median egg infectious doses; HPAI, highly pathogenic avian influenza; hpi, hours postinoculation.
aTotal number viral antigen-positive feathers/total number of feathers observed in skin sections for each feather tract from all birds in the group at that time point.
In ducks, the prevalence of antigen-positive feathers varied with the infecting virus (Tables 2 and 3; Suppl. Table S2). Of the 26 ducks demonstrated to be infected with the Indonesian viruses, viral antigen was observed in feathers of only 4 ducks infected with IDN 34368 and 1 duck infected with IDN 32226. Of these 5 ducks, antigen-positive feathers were found in the primary tract in 3 ducks (16.7% to 33.3% of feathers), the capital tract in 2 ducks (1.5% to 3.5% of feathers), and the caudal tract in 2 ducks (16.7% of feathers in both) (Suppl. Table S2). Where it did occur, the antigen in these IHC sections was sparse. Statistical analysis could not be conducted on these data from the Indonesian virus duck groups because the majority of data points were zero.
Number of Ducks With Viral Antigen-Positive Feathers and Proportion of Antigen-Positive Feathers in Skin Sections From 7 Different Regions of Skin (Tracts) in Groups of Ducks Inoculated With Indonesian H5N1 HPAI Virus Isolates IDN 34368 and IDN 32226.
Skin sections were stained by immunohistochemistry to detect influenza A nucleoprotein. See Supplemental Table S2 for individual bird data. dpi, days post infection; EID50, median egg infectious doses; HPAI, highly pathogenic avian influenza.
aNumber of birds with antigen-positive feathers for each tract.
bTotal number antigen-positive feathers/total number of feathers observed in skin sections for each feather tract from all birds in the group at that time point.
cThis duck died suddenly with evidence of virus in ependyma and meninges of brain. 14 All other ducks presented in this table were healthy at euthanasia.
Proportion of Viral Antigen-Positive Feathers in 7 Different Regions of Skin (Tracts) in Ducks Inoculated With 105.3 Median Egg Infectious Doses of Vietnamese H5N1 HPAI Virus Isolate VN 453 and Euthanized During Clinical Disease.
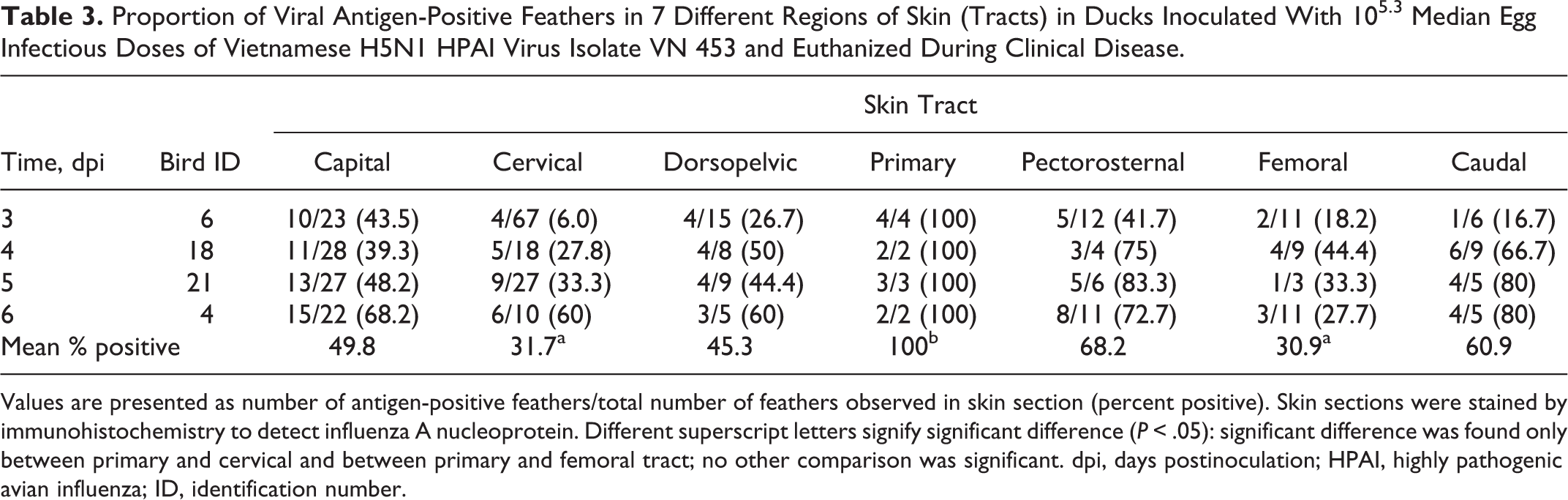
Values are presented as number of antigen-positive feathers/total number of feathers observed in skin section (percent positive). Skin sections were stained by immunohistochemistry to detect influenza A nucleoprotein. Different superscript letters signify significant difference (P < .05): significant difference was found only between primary and cervical and between primary and femoral tract; no other comparison was significant. dpi, days postinoculation; HPAI, highly pathogenic avian influenza; ID, identification number.
In contrast to the Indonesian viruses, viral antigen was detected in feathers of all 7 tracts of all 4 ducks infected with VN 453 that were examined. Antigen was abundant in almost all sections from these ducks. The prevalence of antigen-positive feathers in individual ducks ranged from 6% to 100% in the various tracts (Table 3), with the lowest antigen prevalence in each tract generally occurring in the duck that was sampled at 3 dpi and the highest in ducks that were sampled at 5 and 6 dpi. Across the sample period, the highest mean prevalence of antigen-positive feathers was in the primary tract (100%), the second being the pectorosternal tract feathers (68%) and the lowest the femoral and cervical tracts (31% and 32%, respectively). A significant difference in the mean proportion of virus-positive feathers for VN 453 occurred only between the primary tract and both the cervical and femoral tracts (P < .05).
Viral Antigen May Persist in Feather Epidermis
An interesting finding was that, in the IDN 34368 trial, viral antigen was detected in the epidermis of a retrice of duck No. 8 at 18 dpi (Fig. 5). In this individual, virus was detected in oral swabs only on 2 and 3 dpi; 14 it did not develop clinical signs but had seroconverted by 8 dpi and was euthanized on 18 dpi when the skin sample was taken (Suppl. Table S2).
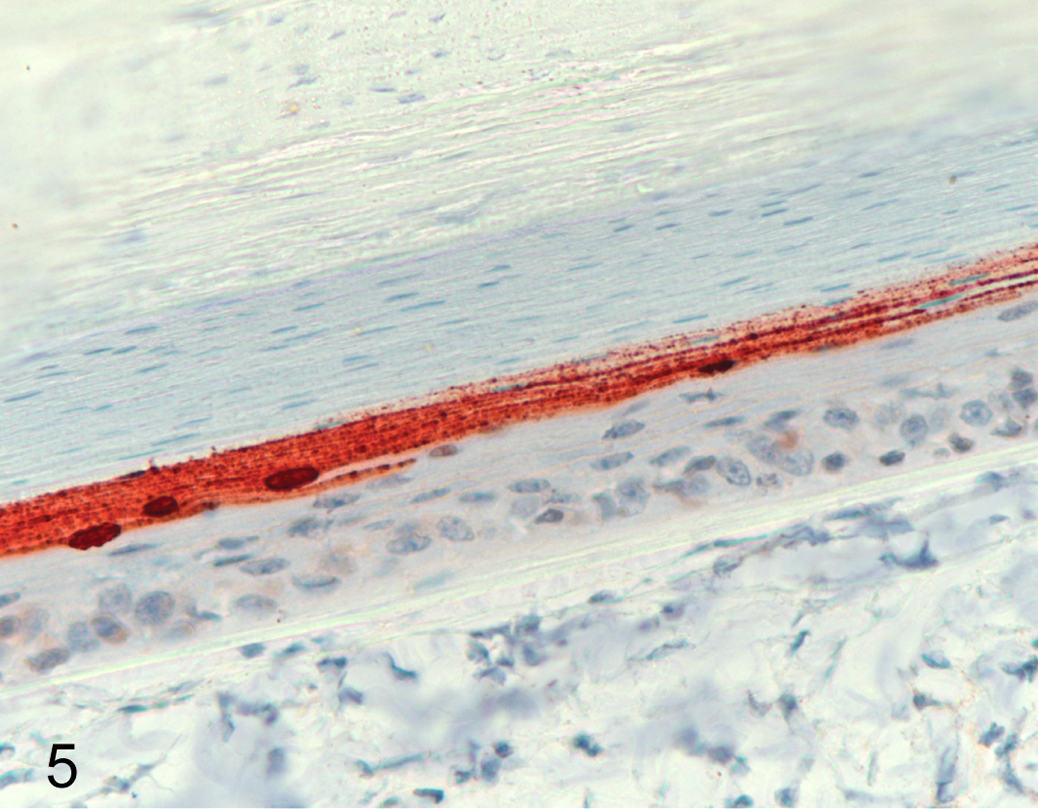
A feather from the primary tract of a duck (No. 8) infected with IDN 34368 taken 18 days postinfection. There is viral nucleoprotein antigen in the corneous layer. Immunohistochemistry for influenza A nucleoprotein antigen using AEC chromogen.
Discussion
Several studies have identified feathers as structures in which HPAI viruses replicate, 2,20 and some have evaluated them as diagnostic samples for avian influenza infection. 1,3,8,11,19 The present study characterized the pathology of HPAI infection in the integument and the morphological basis for viral tropism in feathers in ducks and chickens. Our data support previous findings that HPAI virus infection causes pterylitis and that it replicates to high levels in feather structures and therefore indicates they may be appropriate samples for virus detection.
Morphologically, we found distinct species differences in the infection pattern of feathers between ducks and chickens. Unlike in infected chickens where viral antigen and lesions were detected mainly in the dermis (pulp) of feathers and follicles, in ducks, H5N1 HPAI viruses seem to have a stronger tropism for the epidermis of feathers and follicles. This is consistent with previous reports showing that H5N1 HPAI virus replicated and caused varying degrees of necrosis in feather epidermis of infected ducks. 2,18,20 The reason for the difference in tropism between chickens and ducks is unclear. The observation that virus replication in chickens takes place predominantly in the pulp and not the epithelium of the feather would suggest that this is the tissue that should be preferentially retained for diagnostic purposes, if it was necessary to separate the two. In ducks, it will be necessary to use the wall of the feather, which may necessitate homogenization of this structure.
In chickens, this virus replicates in the endothelium, 14 and the rich vascular network in the pulp of growing feathers would therefore be expected to support viral replication. The density and frequency of viral antigen in feather pulp indicated that immature feathers could be a very reliable tissue type for the detection of infection. This was consistent across different viral strains and different feather tracts, reaching 100% positivity across all but the capital tract. Furthermore, feather pulp contained virus from early in the disease course (24 hpi).
In chickens, we have recommended that feathers be plucked from the pectorosternal tract 8 : these feathers are relatively large, they are abundant and accessible, and their removal does not cause undue pain and bleeding (unlike those of the primary and caudal tracts). This current study supports our recommendation as there was no difference in viral antigen density or prevalence of infection between pectorosternal feathers and the other feather tracts of chickens.
Our data indicated more marked variation within ducks compared with chickens. There was more variation in the proportion of antigen-positive feathers between different virus groups, between different feather tracts within some of the virus groups, and among individual ducks within the groups. There was also more variation in antigen quantity within the morphological regions of skin and feathers in ducks compared with chickens. In ducks infected with the Indonesian viruses, which, apart from 1 sudden death, caused asymptomatic infection, viral antigen was detected in feathers of only 5 of 26 infected birds and from only 3 feather tracts of these birds. In these 5 birds, only a small number of feathers were antigen positive (1–2 positive feathers per duck), and in these feathers, the antigen density was sparse. It is not clear which particular tract in ducks is the best candidate for virus detection, although the retrices from the primary tracts must be a contender. Conversely, the Vietnamese virus, which is known to be pathogenic in ducks, 2 infected a higher proportion of feather structures (up to 100%) in all 7 tracts of all 4 infected ducks, and the antigen density was generally abundant. It appears that the amount of viral antigen in feathers and the proportion of feathers that are antigen positive are correlated with viral pathogenicity, being highly abundant and prevalent in chickens infected with any of the viral strains and least abundant in ducks infected with mild Indonesian-strain viruses.
The immunohistochemistry test of this study appeared to perform as well with chicken skin samples as virus isolation on plucked feathers, 8 as both tests detected 100% of the same infected chickens. In ducks infected with the Vietnamese virus, the immunohistochemistry test on pectorosternal skin samples detected 100% (4 of 4) of infected ducks, compared with virus isolation, which detected virus positivity in 68% of pectorosternal feather samples from these ducks. 8 However, the latter samples were taken over the course of the infection period, not just at euthanasia, where the disease was more advanced and positivity would be expected to be higher. For ducks infected with the Indonesian viruses, immunohistochemistry on skin detected only very few infected ducks. It would appear that immunohistochemistry on skin may be reasonably reliable during clinical HPAI disease but is not sensitive in birds with subclinical infection.
We reported previously that compared with traditional sample types of oropharyngeal or cloacal swabs, feathers were a superior sample for detection of virus in ducks in postacute stages of infection, from about 4 dpi. 8 This is consistent with a previous study that found that virus in feathers persisted 1 to 2 days after its disappearance from oral swabs. 19 The current study provides further evidence for this observation, with the proportion of antigen-positive feathers in most skin tracts of the VN 453–infected ducks increasing between 3 and 6 dpi. This suggests that feather samples may be more reliable for detection of virus in ducks in later stages of infection than in early infection.
An interesting finding was the detection of viral antigen in a feather at day 18 postchallenge from a duck that did not develop clinical signs and from which virus isolation from concurrent swab and feather samples was negative (data not shown). As it was only 1 isolated focus of antigen, its significance is unclear. Although it is not possible to verify that the antigen was associated with infectious virus, it seems reasonable to assume that live virus was present in the feather after the acute infection period. This finding provides information on the potential role of feathers in the ecology of H5N1 HPAI virus, as it shows that virus may persist in feathers to day 18 postchallenge. Also, this shows the potential of feathers for use as diagnostic samples for prolonged periods following H5N1 HPAI infection. Further study is required to determine the prevalence of feathers containing viral antigen up to 3 weeks postchallenge and also to investigate whether the virus is live and therefore constitutes a risk.
Of concern is the finding that in all chickens infected with the 3 HPAI viruses and in ducks infected with the Vietnamese virus, almost all feathers contained viral antigen. With thousands of feathers on just 1 chicken, feathers have a high potential to be carriers for H5N1 HPAI virus transmission from poultry to humans. In infected ducks, H5N1 HPAI virus was predominantly detected in the epidermis of feathers and follicles, including the outer layer of these structures. This suggests that duck feathers have the potential to readily transmit virus, because the outer layer is the part that has direct contact with the environment. Therefore, the potential of feathers in transmitting H5N1 HPAI viruses warrants further research.
Our results support the role of feathers for diagnostic detection of HPAI in chickens and indicate that further investigation into their diagnostic use in ducks is warranted, particularly in prolonged postinfection periods. These findings also raise concerns regarding the role of feathers in the spread of H5N1 HPAI viruses among poultry and the transmission of virus to humans.
Footnotes
Acknowledgements
We thank the Animal Studies Team at AAHL for assistance with animal trials, Manabu Yamada for assistance with histopathology, the Bioassays R&D Team at AAHL for the supply of the primary antibody used in the immunohistochemistry test, and Viviana González-Astudillo for the artwork of Figures 1 and ![]() .
.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially funded by the Australian Centre for International Agricultural Research (ACIAR) (project AH/2004/040). H.N. and H.W. were supported by ACIAR John Allwright Fellowships. This work was performed at AAHL, which is a National Collaborative Research Infrastructure Strategy (NCRIS)–funded facility.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.