
Abstract
West Nile virus (WNV) infection has been detected in many species of birds and mammals, but scant information is available about the disease in small ruminants. West Nile virus was diagnosed in 6 sheep with neurological signs and encephalitis, in California between 2002 and 2014. All sheep had severe lymphoplasmacytic meningoencephalitis. Lymphoplasmacytic myelitis was also detected in 2 sheep where the spinal cord was examined. Brain tissue was positive for WNV detected by polymerase chain reaction in 6 of 6 sheep and by immunohistochemistry (IHC) in 5 of 6 sheep. Viral antigen was not detected by IHC in extraneural tissues in the 3 sheep examined. West Nile virus RNA was sequenced from 2 of 6 sheep, and each one clusters closely with WNV isolated from mosquito pools from nearby locations at similar times. West Nile virus was the most common cause of viral encephalitis in sheep diagnosed at this laboratory between 2002 and 2014, accounting for 6 of 9 sheep.
West Nile virus (WNV) is an arbovirus belonging to the genus Flavivirus, family Flaviviridae. In California, the first human case of WNV infection occurred in 2002, the virus was first isolated in July 2003 and has been detected since then in humans and in a variety of birds and domestic and wild animals. 7 West Nile virus has a wide host range and has been detected in at least 48 species of mosquitoes, 320 species of birds, 2 species of reptiles, and 25 mammalian species. 6 The virus is mainly maintained by bird–mosquito–bird cycle, and corvids (eg, crows) are the main amplifying host. 3
Infected wild birds typically undergo a prolonged viremia which allows viral distribution to almost every organ. 3 Histological lesions of lymphoplasmacytic and histiocytic infiltrates with cellular degeneration, necrosis, and hemorrhage most commonly affect the central nervous system (CNS), heart, spleen, liver, and kidneys but may be minimal in acute cases. 3 Horses are the most susceptible domestic mammal. 1 Lesions in horses are limited to the CNS, mainly affecting gray matter of brain stem and thoracolumbar spinal cord, appearing grossly as hemorrhage and malacia and histologically as lymphoplasmacytic encephalomyelitis with gliosis, glial nodules, occasional neuronal degeneration, and necrosis. Ruminants are not considered significant hosts, and there have been only 3 individual case reports of meningoencephalitis associated with WNV infection in sheep. 4,8,9 In this study, we describe the clinical, histopathological, and immunohistochemical findings of naturally occurring WNV infection in 6 domestic sheep in California.
A retrospective search of the California Animal Health and Food Safety Laboratory System (CAHFS) from 2002 (the first year WNV was detected in California) to 2014 yielded 6 sheep with WNV-associated neurological disease.
The clinical history indicated that only a single animal was affected in each herd. The year, season, age, sex, breed, and relevant clinical signs of the 6 sheep are summarized in Supplemental Table 1. Briefly, all cases occurred in summer or fall, the more common clinical signs included fever (3 of 6), anorexia (3 of 6), and neurological signs (5 of 6) that included staggering, shaking, circling, drooling, head tilt, blindness, and tremors. All animals were found dead. There was no apparent age, breed, or sex predisposition.
Full necropsies were performed in all 6 sheep. Gross findings included variable pulmonary congestion and edema in all animals and foam in the trachea of 3 sheep. No gross lesions were observed in the CNS of any animal. Tissues examined histologically in all cases were obtained from brain, lung, heart, liver, spleen, kidney, skeletal muscle, rumen, abomasum, small intestine, and colon. The spinal cord was only examined in 2 sheep. All sheep had histologic lesions of severe lymphoplasmacytic meningoencephalitis (Fig. 1). Inflammatory changes were uniformly widespread in the brain and involved the cerebrum, cerebellum, brain stem, and thalamus. These changes were characterized by severe lymphoplasmacytic perivascular cuffing with occasional histiocytes and neutrophils. Scattered foci of gliosis, necrotic neurons, and neuronophagia were also detected. The lesions in cerebellum included occasional foci of gliosis and occasional necrotic Purkinje cells. In the 2 sheep in which the spinal cord was examined histologically, the lesions involved the gray and white matter but were more severe and widespread in the gray matter. In 1 sheep (sheep No. 3), where multiple sections of spinal cord were examined, the lesions were more severe in the cervical and thoracic spinal cord.
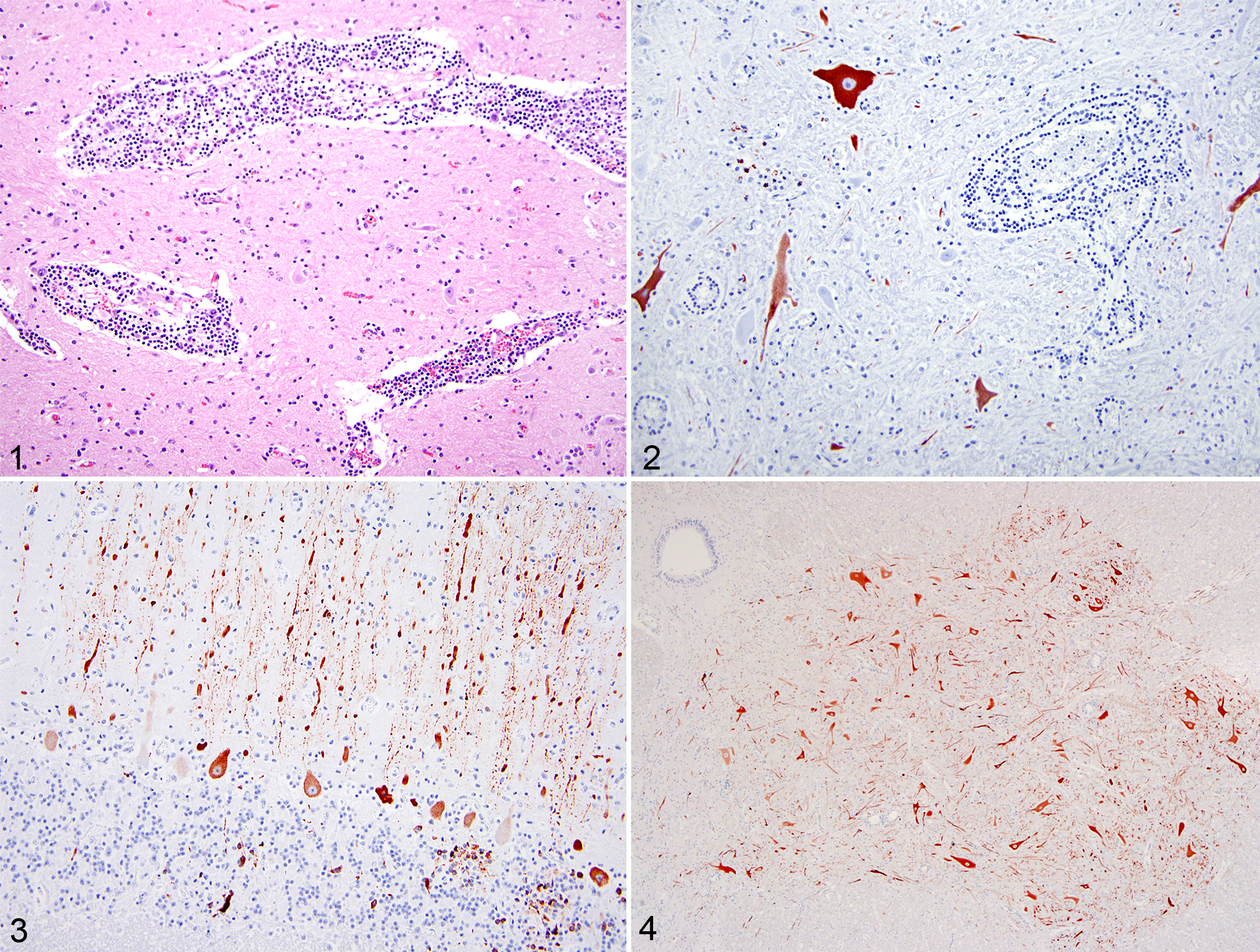
Central nervous system tissue samples from all 6 animals were positive for WNV detected by polymerase chain reaction (PCR). 5 Partial WNV genomes from 2 sheep detected in summer of 2014 (5 and 6) were sequenced (see Supplemental Materials for details). Each sequence was compared to a Californian reference sequence (WNV CA04, 2004), other California 2013–2014 WNV sequences from mosquitoes, other published sequences from 2003 to 2011, 2 and outgroup sequences Kunjin virus and St. Louis encephalitis virus from GenBank (National Institutes of Health, Bethesda, MD).
Immunohistochemistry (IHC) for WNV using a rabbit polyclonal antisera (VRL 81-015, VRL Laboratories, Centennial, CO), dilution 1:1000, after 8 minutes of antigen retrieval with proteinase K and using Envision + HRP (Dako K4003, Agilent Technologies, Santa Clara, CA) as detection system (negative control, VRL rabbit sera) was performed on CNS sections with lesions obtained from all 6 sheep and in extraneural tissues obtained from 3 sheep, including heart, lung, kidney, adrenal gland, liver and spleen (case Nos. 3, 5, and 6), small intestine in 2 sheep (case Nos. 3 and 6), rumen (case No. 6), abomasum (case No. 6), and spiral colon (case No. 5) in 1 sheep. Strongly positive immunolabeling was detected in the CNS obtained from 5 of 6 sheep (case Nos. 2, 3, 4, 5, and 6), whereas 1 (case No. 1) was negative. Large amounts of uniformly widespread viral antigen were detected within neuronal soma and axons, and, to a lesser extent, there was scattered granular staining of glial cells and segmental staining of dendritic processes located in areas of severe nonsuppurative meningoencephalitis (Fig. 2) and also in areas with no or minimal histological lesions (Figs. 3 and 4). Conversely, there was no IHC staining in some CNS sections that had significant nonsuppurative inflammation. All extraneural tissues examined were negative for WNV antigen as detected by IHC.
Other laboratory findings in these animals included mildly elevated hepatic copper levels (4 of 6), marginally low hepatic selenium levels (2 of 6), and abomasal Haemonchus sp. (2 of 6) and Ostertagia sp. (1 of 6). In 1 sheep (case No. 5), a mild chronic multifocal lymphocytic myocarditis (which was negative for WNV by IHC) and a chronic mild multifocal interstitial lymphocytic mastitis of undetermined cause were present. In all sheep, brain samples were negative for rabies detected by fluorescent antibodies technique, and no Listeria sp. was isolated from any sleep after 4-week incubation in cold-enrichment media.
Viral diseases causing lymphocytic encephalitis in sheep include bluetongue, small ruminant lentiviral encephalomyelitis [Caprine arthritis encephalitis (CAE)/ovine progressive pneumonia, (OPP)], rabies, pseudorabies, Borna disease, louping ill, and Nipah virus infection.
Between 2002 and 2014, a total of 2 083 ovine necropsy submissions were received at CAHFS. Encephalitis, meningoencephalitis, or encephalomyelitis was diagnosed in 53 live animals and 14 aborted fetuses, which were excluded from this study. Of these 53 cases, 22 (41.5%) were caused by bacterial infections, 9 (17%) by viruses, 3 (5.6%) by protozoa, 1 (1.9%) by a zygomycete fungus, and 18 (34%) were idiopathic. Of the 9 cases of encephalitis or encephalomyelitis with a definitive viral etiology, 6 were due to WNV, 2 were associated with bluetongue virus (both animals were positive serologically and 1 was positive by PCR in spleen tissue), and 1 with small ruminal lentivirus (CAE/OPP) based on positive serology. These 3 sheep were negative for WNV detected by IHC performed on brain tissue.
The 18 cases of idiopathic encephalitis or encephalomyelitis were retrospectively tested for WNV by PCR on frozen CNS samples (4 sheep), IHC (13 sheep), or both techniques (1 sheep), and results were all negative. The encephalitis lesions in these cases ranged from minimal to severe and were not always presumed as the cause of death.
These findings suggest that sheep may not only be more susceptible to WNV infection than previously suggested 4,8,9 but WNV was the most common viral cause of encephalitis or encephalomyelitis at CAHFS in the past 13 years. The diagnosis of WNV in these case series was confirmed by PCR in all 6 sheep and further supported by positive WNV IHC in 5 of the 6 sheep. Immunohistochemistry showed marked CNS tropism, with intense staining of neuronal soma and axons indicating a high antigenic load in the CNS. One of the 6 WNV PCR-positive animals was negative for WNV IHC on several sections of the CNS. The causes for this negative result are unknown but may be due to a different (possibly earlier) stage of the disease, low viral load within the CNS, lower test sensitivity, or inadequate sample preservation. In 3 sheep, several extraneural tissues were tested for WNV IHC, and all sections were negative, suggesting that the viral infection in sheep is restricted to the CNS. As in horses, WNV infection in sheep seems to be limited to the CNS. However, unlike horses and somewhat resembling CNS lesions in birds, large amounts of viral antigen may accumulate in the CNS of infected sheep. The intermittent detection of WNV-associated disease in sheep (3 in 2005, 1 in 2012, and 2 in 2014) does not correlate well with the more consistent detection of WNV-associated disease in a variety of avian species and horses received at our laboratory during that time. The reason for this is unclear but may be related to a lower susceptibility of sheep to the disease and/or to the fact that WNV may be underdiagnosed in this species. In summary, WNV should be considered an important differential diagnosis in sheep with encephalitis or encephalomyelitis, and both PCR and IHC are useful diagnostic tools to confirm the disease.
Footnotes
Acknowledgements
The authors are thankful to the histology team, Wendy Kerns, Alisha Curtis, Rosa Manalac, Patrick Pong, and Karen Sverlow.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Kelly Symmes was supported by a Students Training in Advanced Research Fellowship at the School of Veterinary Medicine in summer 2015. WNV sequencing was also partially supported by a University of California Agriculture and Natural Resources Grant to Andrea Townsend.