
Abstract
Encephalitic flaviviruses are important arthropod-borne pathogens of humans and other animals. In particular, the recent emergence of the West Nile virus (WNV) and Japanese encephalitis virus (JEV) in new geographic areas has caused a considerable public health alert and international concern. Among the experimental in vivo models of WNV and JEV infection, mice and other laboratory rodents are the most thoroughly studied and well-characterized systems, having provided data that are important for understanding the infectious process in humans. Macaca monkeys have also been used as a model for WNV and JEV infection, mainly for the evaluation of vaccine efficacy, although a limited number of published studies have addressed pathomorphology. These animal models demonstrate the development of encephalitis with many similarities to the human disease; however, the histological events that occur during infection, especially in peripheral tissues, have not been fully characterized.
Keywords
The genus Flavivirus comprises more than 70 viruses, many of which are arthropod-borne human pathogens. 59 The genome of flaviviruses is a single-stranded RNA of approximately 11 kb. Based on antigenic and genetic characteristics, arthropod-borne flaviviruses have been classified into four groups: the Japanese encephalitis virus (JEV) complex, the tick-borne encephalitis virus complexes, the Dengue virus, and the yellow fever virus. 4,10,20,44,57,63 The JEV and tick-borne encephalitis complexes are neurovirulent and can cause encephalitis in humans. In contrast, yellow fever virus and Dengue virus are more viscerotropic and can cause hemorrhagic fever. 40 Among the mosquito-borne flaviviruses, current major global concerns include Dengue virus, JEV, West Nile virus (WNV), and yellow fever virus. 60 JEV and WNV, which belong to the JEV complex, are serologically cross-reactive with each other. 64,115
Development of flaviviral encephalitis in infected hosts has been thought to occur mainly via hematogenous spread of the virus to the central nervous system (CNS). 13,79 After injection of virus-containing saliva from mosquito, the virus replicates in local tissues and regional lymph nodes. This results in a primary viremia and subsequent infection of extraneural tissues (eg, the reticuloendothelial system) where further replication leads to a more sustained secondary viremia, which causes CNS infection. The mechanism underlying the crossing of the blood–brain barrier by flaviviruses during natural infection remains uncertain.
Although a variety of animal models of JEV and WNV exists, the rodent models are the most thoroughly characterized of these and are thus the focus of our review. A considerable body of knowledge concerning the pathogenesis of flaviviral encephalitis has been generated from experimental infection studies using rodent models (as reviewed in Charlier et al 14 and Nalca et al 78 ). However, the pathomorphological changes that occur in those models during infection have not been fully characterized.
Pathological Changes in Mice Infected With WNV
WNV is maintained in an enzootic cycle between mosquitoes and wild birds, but it can also infect humans, horses, and other vertebrate animals. 43 Human infection may result in a febrile illness that can progress to lethal encephalitis with cognitive dysfunction and poliomyelitis with flaccid paralysis. 12,88,101 WNV infection was previously recognized as being endemic to Africa, Europe, and Asia; 76 however, WNV outbreaks occurred in Romania, Russia, Israel, and the Americas in the 1990s with fatal encephalitis as a significant feature. 19,60 Seroprevalence studies of the outbreaks suggest that approximately 20% of infected individuals develop clinical manifestations characterized as WNV fever. 88,122 About 1 in 150 people infected with WNV develop a neuroinvasive disease. 89,123 The mortality rate following neuroinvasive infection is approximately 10%. 81,88 Pathologic evaluation of human cases with WNV encephalitis reveal that patients exhibit perivascular lymphocytic infiltrates, microglial nodules, loss of neurons, and viral antigens predominantly within the brainstem and anterior horn of the spinal cord, although the cerebral cortex, cerebellum, and posterior horn of the spinal cord are also involved. 39 The WNV antigen has been detected in the cytoplasm of neurons and neural processes. 39 In immunosuppressed individuals, infection can disseminate throughout the CNS. 54
Animals that have been used to model experimental WNV infection include mouse, rat, 28 hamster, 117,127 monkey, 37,91,94 horse, 8 pig, 116 dog, 3 and cat. 3 Among the models, those based on the mouse are the most intensively studied and have fostered a significant understanding of WNV pathogenesis, as previously reviewed. 22,96 After intradermal inoculation with WNV, initial viral replication is thought to occur in Langerhans dendritic cells of the skin. 6,50 The infected Langerhans cells migrate to draining lymph nodes, 9,49 and the virus is considered to enter the bloodstream through the lymphatic and thoracic ducts. 62 This primary viremia disseminates the virus to peripheral tissues, including the spleen, where further viral replication occurs. 23,108 It is conceivable that most viral particles in viremic blood are free and exist in the plasma. 61 By the end of the first week after infection, WNV is largely cleared from the serum and peripheral organs, and infection of the CNS is observed. 105 Mice that succumb to infection develop CNS pathology similar to that observed in human WNV cases. 29,85 Mouse lesions described in the literature include necrosis of neurons, perivascular mononuclear infiltrates mainly composed of macrophages and lymphocytes, glial/microglial nodules, neuronal satellitosis, and neuronophagia in the brainstem, cerebral cortex, hippocampus, cerebellum, and spinal cord. 13,31,106
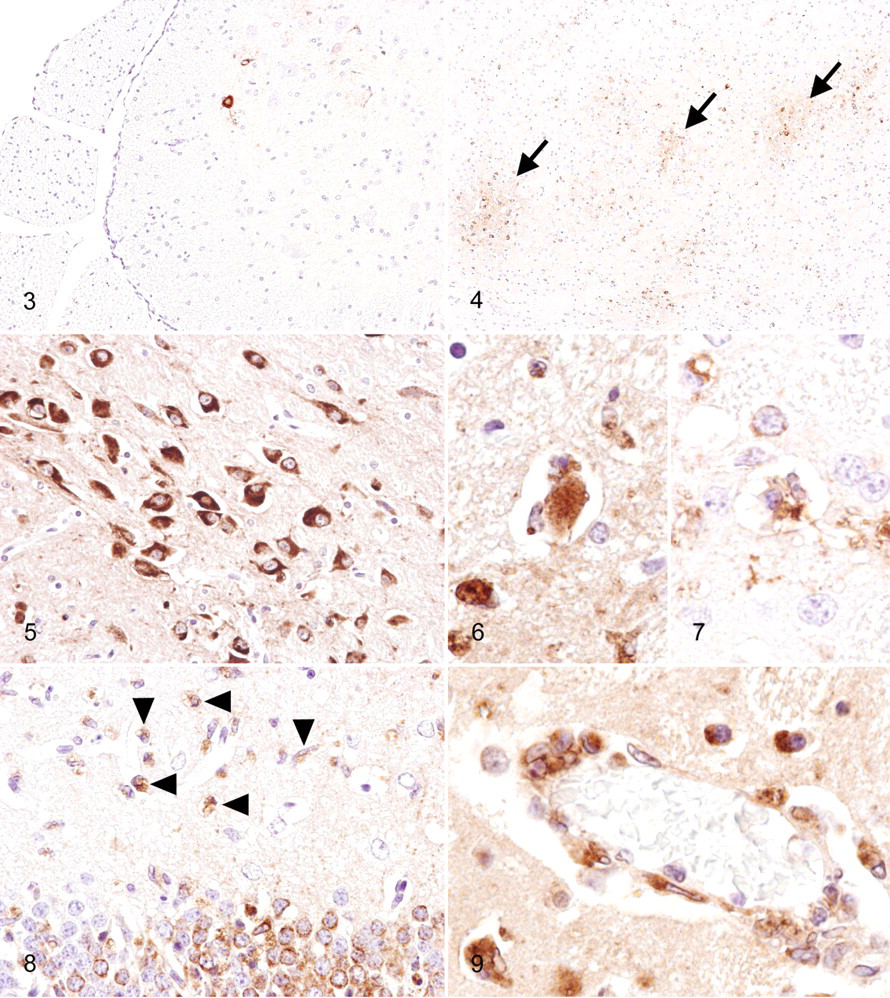
Many studies recently performed on the pathogenesis of North American WNV isolates used the C57BL/6 mouse strain and genetically engineered immunocompromised mice in the C57BL/6 background. C3H/HeN mice showed higher morbidity and mortality than that of C57BL/6 mice after infection with the recombinant virus derived from a 2000 New York isolate, although viral tropism, viral load, and kinetics did not differ substantially between these strains. 6 Our unpublished observations from 6-week-old C3H mice that received footpad inoculation of 1,000 plaque-forming units of a NY99-6922 strain-derived plaque isolate revealed that infected mice developed neuronal necrosis (Fig. 1 ) with infiltration of reactive microglia, neuronophagia, and perivascular infiltration of macrophage/microglial cells and lymphocytes (Fig. 2 ) into the gray matter of the CNS from day 9 postinfection. These lesions were distributed preferentially in the brainstem, cerebral cortex, caudate putamen, thalamus, and spinal cord; however, the hippocampus and cerebellum were involved in many cases. Mild meningitis was also seen. Small hemorrhagic foci were occasionally observed. These histological findings are consistent with those reported by Garcia-Tapia et al 31 using 8-week-old C57BL/6 mice that were inoculated in the footpad with 100 plaque-forming units of a 2002 Missouri isolate. Immunohistochemistry (IHC) revealed that viral antigens were present in the CNS, at first in a few neurons of the ventral horn of the lumbar spinal cord on day 7 postinfection (Fig. 3 ). Infected neurons were located on the ipsilateral side of inoculation, which is consistent with the findings of previous studies 97,120 and suggests transneural spread of the virus via the sciatic nerve. Foci of viral antigen were then seen in multiple regions, including the brainstem, cerebral cortex, caudate putamen, and cervical spinal cord (Fig. 4). Viral antigens were mainly detected in the cytoplasm of neurons and neuronal processes (Fig. 5), although a few cells morphologically suggestive of astrocytes were positive. Neuronophagia was common (Fig. 6, 7). In heavily infected foci, proliferation of microglia was prominent, and many macrophage/microglial cells contained viral antigens (Fig. 8). These macrophage/microglial cells also accumulated in perivascular areas as a morphological indication of the involvement of microglial cells in viral clearance and/or antigen presentation (Fig. 9). Vascular endothelial cells in the infected foci sometimes showed positivity for WNV antigens, and in most cases they were associated with the infiltration of virus antigen-containing macrophages/microglia. Diffuse severe inflammation and antigen distribution occurred in a small number of mice.
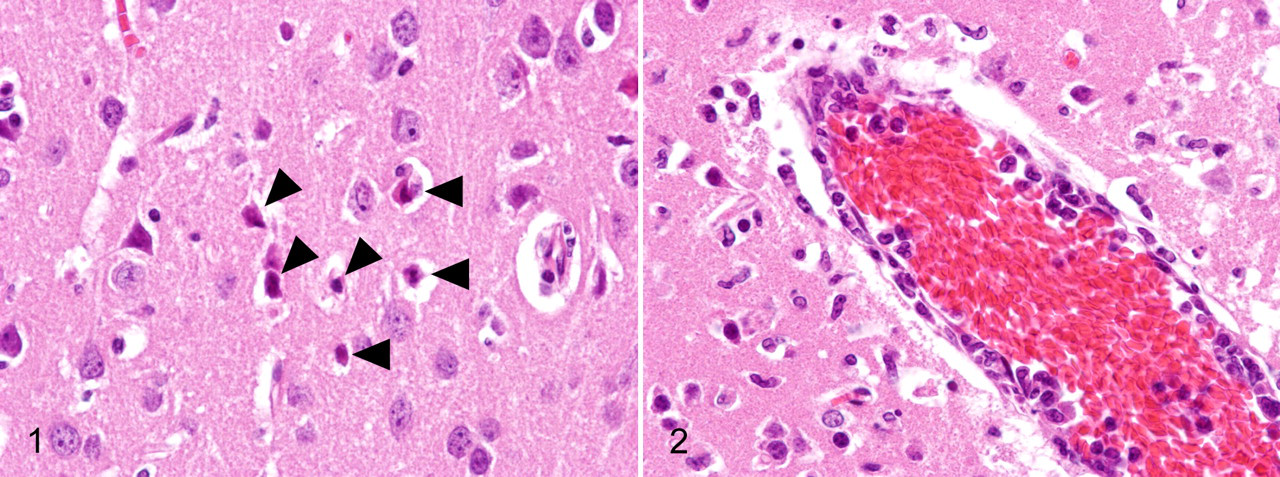
Several laboratories have analyzed the mode of lymphocyte recruitment into the CNS of WNV-infected mice. CD45 (common leukocyte antigen)-positive leukocytes were detected in the inflamed parenchyma of the CNS of WNV-infected mice. 106 Trafficking of CD8+ cells to the brain is more predominant than that of CD4+ T cells after WNV infection in mice. 105,107,121 In response to WNV infection, neurons secrete the CXCL10 chemokine, which in turn recruits effector CD8+ T cells via an interaction with the CXCR3 chemokine receptor. 53 The CCR5 chemokine receptor of leukocytes participates in the promotion of leukocyte recruitment into the brain of WNV-infected mice and plays a crucial role in host survival. 36 Other chemokines, such as CCL5 (RANTES), CCL3 (MIP-1α), and CCL4 (MIP-1β), are also upregulated in the brains of WNV-infected mice. 103
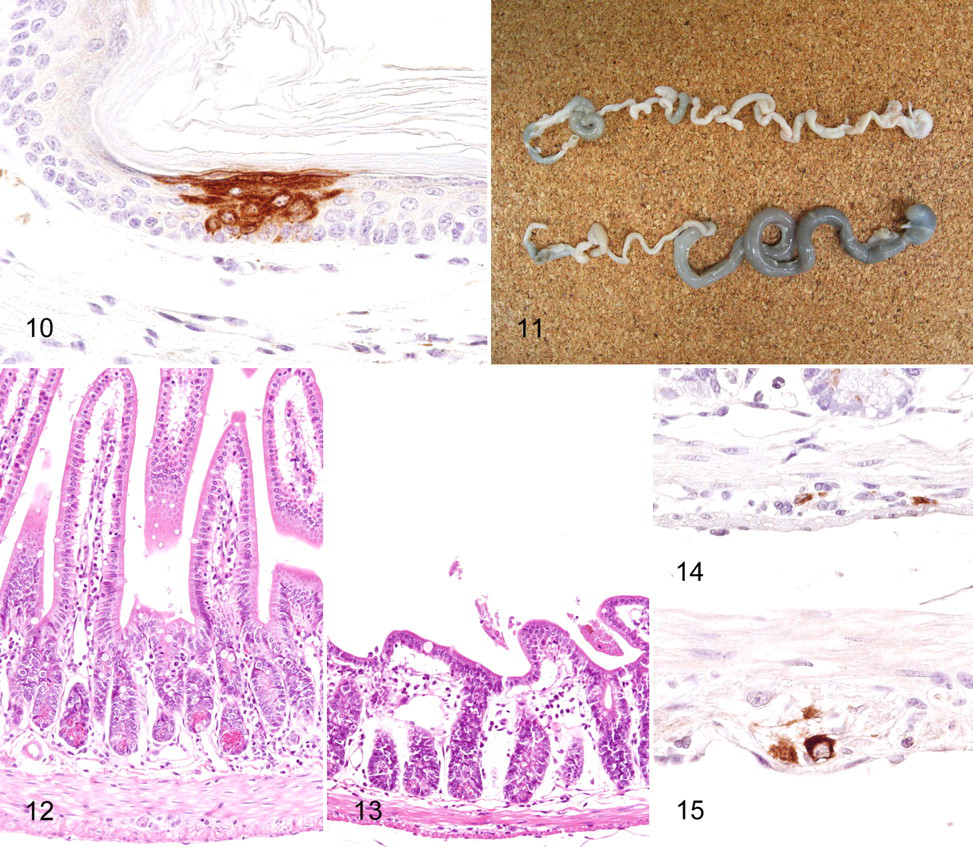
The pathological changes that occur in peripheral tissues infected with WNV remain incompletely characterized. In human cases, WNV has been detected in renal and splenic tissues, with cell culture and reverse transcriptase polymerase chain reaction; 85 in renal tubular epithelium of an AIDS patient, with IHC; 39 in testis of a transplant recipient, with electron microscopy; 110 and in macrophages, intravascular mononuclear cells, and vascular endothelia of the skin, stomach, lung, liver, kidney, spleen, and bone marrow, with IHC. 86 Systemic distribution of WNV in severely immunocompromised transplant recipients has been reported in whom the epithelial cells of the lung, pancreas, thyroid, intestine, stomach, esophagus, bile duct, skin, prostate, and testes were positive for viral antigen, based on IHC. 1 In mice, the WNV antigen has been detected immunohistochemically in macrophages and fibrocytes of the skin, in a few macrophages of the spleen, and in some renal tubular cells with no signs of inflammation. 31 Our unpublished observations from C3H mice that received footpad inoculation of WNV, as described above, revealed that viral antigens are rarely present in the squamous epithelial cells of the footpad inoculation site (Fig. 10 ). This finding seems to be consistent with that of a previous report in which the skin, duodenum, and pancreas of C3H mice were identified as novel sites of viral replication. 6 Reportedly, monocytes and a subset of lymphocytes are able to support the productive replication of WNV after infection. 32 However, using IHC on paraffin-embedded sections, we did not detect specific WNV immuno reactivity in the spleen and lymph nodes of infected mice, both of which are major organs of extraneural viral replication. 6,108 It may be that the level of viral replication in each mononuclear cell is below the threshold of detection of the conventional immunohistochemical technique. Significant non-CNS lesions associated with viral antigens were frequently observed in the digestive tracts of C3H mice from day 7 postinfection, in which the upper small intestines and sometimes the stomachs were abnormally dilated and filled with a greenish watery content (Fig. 11). Villi of the upper small intestines of WNV-infected mice were markedly shortened (Fig. 12, 13). Degeneration and necrosis of myenteric ganglia cells with WNV immunoreactivity were a consistent feature (Fig. 14). The myenteric plexi of the upper small intestine were most severely affected, whereas ganglia cells in other regions were positive for WNV (Fig. 15). The lesions observed in the intestines of WNV-infected C3H mice are not features that are relevant in human WNV cases. As reported previously, 31 thymic atrophy was common in moribund mice.
In surviving immunocompetent mice, WNV cleared from all tissues within 2 to 3 weeks after infection. Innate immunity, humoral immunity, and T cell–mediated immunity are all involved in viral clearance and in the protection of mice against WNV infection, as previously reviewed. 96 Among them, humoral immunoglobulin M response is the most critical for averting lethal outcomes in acute WNV infection. 23,24 In rodent models, persistent infection has been reported in the brains of CD4+ T cell–deficient, CD8+ T cell–deficient, or perforin-deficient mice and in the brain and kidney of infected hamsters. 105,107,109,117,127 Persistent infection in WNV-infected immunosuppressed patients has been reported in whom viremia was detected for more than 60 days. 5
Wild-type mice are resistant to flavivirus infection, whereas laboratory mouse strains exhibit varying degrees of susceptibility to flaviviruses. 17,98 The susceptibility of inbred mouse strains to WNV has been mapped to a mutation in the 2′-5′-oligoadenylate synthase 1b gene (OAS1b), which results in the expression of a truncated OAS isoform. 65,87 The OAS1b gene is a member of an interferon-regulated gene family that is involved in the degradation of viral RNA. Knocking in the OAS1b resistance allele into a susceptible mouse strain generated mice with the flavivirus resistance phenotype. 100 Although the detailed mechanisms of how OAS gene alleles affect WNV pathogenesis remain uncertain, pathologists should keep in mind this genetic phenomenon that renders inbred mice highly susceptible to flavivirus infection when compared with animals in other models.
Pathological Changes in Other Animal Models of WNV
Hamsters inoculated intraperitoneally with a NY isolate of WNV develop viremia and subsequent meningoencephalitis. 127 Infectious viral particles were cultured from the brains of convalescent hamsters up to 53 days after the initial infection, which indicates the establishment of a persistent viral infection. 127 In hamsters that were chronically infected with WNV, the antigen was found in macrophages and vascular endothelial cells within renal interstitia. 117
Studies are limited that describe the histopathological changes that occur in WNV-infected monkeys. Pogodina et al 91 reported that intracerebral inoculation of rhesus monkeys with the African, Asian, and European isolates of WNV results in the establishment of persistent viral infection in the CNS and in other organs (ie, spleen, lymph nodes, and kidney) up to 167 days after the initial infection. Infected monkeys that experienced encephalitis or febrile or asymptomatic infection exhibit a protracted pathological process in their CNS. The histopathological findings in the acute stage of WNV-induced fatal encephalitis in monkeys include marked perivascular and diffuse inflammatory infiltrates, neuronal degeneration and necrosis, and glial reactions in the gray matter—especially in the cerebral cortex, subcortical ganglia, thalamus, medulla oblongata, midbrain nuclei, cerebellum, and spinal cord. A protracted pathological process was observed in the CNS of monkeys examined at various points of convalescence or in animals with asymptomatic infection. Lesions were mainly found in the cerebellum, brainstem, and anterior horns of the spinal cord. Loss of cerebellar Purkinje cells and spinal motoneurons was characteristic. Fresh foci of inflammation with neuronal death and formation of neuronophagic nodules were observed up to 2 months after inoculation, simultaneously with older lesions that exhibited a reparative glial reaction. Replacement of the dead nerve cells by astrocytes became prominent at 167 days after infection. Monkeys that were intracerebrally challenged with the NY99 strain of WNV exhibit a lethal outcome, which is preceded by fever and tremors that progress to ataxia and spasticity. 2 Generally, intradermal or subcutaneous infection of monkeys with WNV does not cause clinical disease, as it does in humans, 90,94 although a low level of viremia develops postinfection, which can be monitored via serial blood collections. 58,90 The prevention or inhibition of the development of viremia by prior vaccination is a demonstration of the protective efficacy of the vaccine, regardless of clinical presentation. 58,90 A case of severe nonsuppurative meningoencephalitis due to naturally acquired infection with WNV was reported in a Barbary macaque (Macaca sylvanus) housed in a zoo. 84
Wild birds infected with WNV develop prolonged high levels of viremia, thus acting as the primary amplifying hosts. The WNV strains that circulate in North America and the related strains from Israel are considered more virulent for birds than those previously isolated in Africa, Asia, and Europe. 55 WNV reportedly infects all major organ systems and a variety of individual cell types in wild birds; 113 thus, it yields pathological features that are different from those present in human WNV cases. The most prominent lesions observed in wild birds (27 birds representing 8 orders and 14 species) during the 1999 WNV outbreak in New York city were gross hemorrhage of the brain, splenomegaly, meningoencephalitis, and myocarditis. 113 The WNV antigen was detected most frequently in the kidney, heart, and brain of these animals but was also found in the spleen, liver, adrenal gland, intestine, pancreas, lung, and ovary. Infected cells included neurons and glial cells of the brain, spinal cord, and peripheral ganglia; myocardial fibers; macrophages and blood monocytes; renal tubular epithelium; adrenal cortical cells; pancreatic acinar cells and islet cells; intestinal crypt epithelium; fibroblasts; and smooth-muscle cells. 113 High mortality associated with WNV is prominent among American crows and other American corvids. 55,80 The most common histological findings reported in naturally infected American crows are multifocal necrosis of the spleen and bone marrow. 126 The WNV antigen was consistently detected in the heart and kidney of these animals but was also found in the bone marrow, duodenum, proventriculus, liver, lung, spleen, pancreas, and brain. 126 Monocytes/macrophages appear to be important targets for virus replication and contribute to the systemic dissemination of the virus. 113,124 WNV is not likely to cause significant disease in domestic chickens (Gallus gallus), whereas chickens that were inoculated subcutaneously with the NY strain of WNV developed relatively low levels of viremia, myocardial necrosis, nephritis, and pneumonitis. 102
Mouse Models of JEV Infection
Japanese encephalitis is a major viral encephalitis in Southeast Asian countries, with 30,000 to 50,000 cases reported annually. The disease affects mostly children, but all age groups are affected in areas where the virus was recently introduced. 119 Although the ratio of apparent to unapparent infection may vary across populations, it is about 1 in 300 in rural Asia. 119 Case fatality rates range from 5 to 40%, and most survivors who develop severe disease have neurological and/or psychiatric sequelae. 40 Pigs and birds serve as effective amplifying hosts in the transmission cycle of JEV. 7
Patients who succumb to Japanese encephalitis usually die within 1 week of hospital admission for deepening coma and respiratory arrest. 48 Pathological studies of Japanese encephalitis in children 48 have reported moderate meningeal inflammatory response, marked perivascular cuffing, infiltration of cells into the parenchyma, neuronophagia, and glial nodule formation in the cerebral cortex, thalamus, and brainstem. Polymorphonuclear cells are present in moderate numbers in the meninges and perivascular cuffs. JEV antigens are localized in neurons (IHC), with no evidence of glial cell infection. The highest concentration of infected neurons is found in the thalamus and brainstem. In perivascular cuffs, T cells are more predominant than B cells and macrophages. CD8+ T cells are a minority among the T-cell population. The majority of the inflammatory cells infiltrating the parenchyma are macrophages. In a recent study of human cases, acellular necrotic foci (“punched out” appearance), vascular endothelial cell damage, and prominent astrocytic activation were seen. 33 A significant degree of antigen localization in the hippocampus, temporal cortex, and cerebellum, in addition to the thalamus and brainstem, was reported. 21 Flaccid paralysis is occasionally seen, and pathological studies that included the spinal cord reported extensive involvement of the anterior horn. 25,27,128 Conscious patients with polio-like acute flaccid paralysis have been reported. 16,111,112
Histological findings of the brain lesions in JEV-infected mice include neuronal degeneration and necrosis, neuronophagia, microglial proliferation forming glial nodules, mild perivascular hemorrhage, and perivascular cuffing. 33,40 Although the severity of the inflammation and the distribution of viral antigens are influenced by the phenotype of the inoculated viruses and by the route of inoculation, respectively, the histological changes occur mostly in the gray matter and are predominantly found in the cerebral cortex and hippocampus; 30,33,42 however, they are also frequently found in basal ganglia, thalamus, brainstem, and other regions.
In the pathogenicity of neurotropic viruses, the outcome of intracerebral inoculation reflects viral neurovirulence, whereas the outcome of peripheral inoculation (subcutaneous or intraperitoneal) reflects neurovirulence and neuroinvasiveness. 118 Intraperitoneal inoculation is a preferential route of peripheral virus delivery in rodent models of JEV infection, as opposed to subcutaneous inoculation (eg, footpad), which mimics the bite of an infected mosquito vector. This is due in part to the observation that mice, especially older animals, are somewhat more resistant to JEV infection when the virus is administered subcutaneously, 46,71 although several studies have reported an absence of significant differences in mortality and infectivity among intraperitoneal, intravenous, subcutaneous, and intradermal inoculations. 38,70
Suckling mice are highly susceptible to JEV, and their brains have been used for efficient virus propagation. JEV infection in infant mice results in fatality, either by intracerebral or peritoneal inoculation. Weanling mice are also susceptible to intracerebral JEV inoculation; however, there is considerable variation in the development of fatal encephalitis when the virus is administered via peripheral inoculation. 45,46 This variation stems from the titer of the virus, the viral strain used, and the age of the mice. In mice inoculated subcutaneously, the early phase of viral replication occurs in the peripheral tissues, which include the liver, spleen, heart, and kidney, and is followed by the secondary phase, which occurs in the brain. 47 It remains unclear whether this scenario is applicable to all cases that develop CNS infection after peripheral inoculations; that is, JEV sometimes establishes CNS infection without conclusive evidence of prior viral proliferation in the peripheral tissues. In the hamster model of the St. Louis encephalitis virus, which is a flavivirus antigenically related to both JEV and WNV, the possible involvement of the olfactory pathway in natural infection has been proposed because of the observation of initial CNS involvement of the olfactory bulbs after intraperitoneal infection. 72 However, the role of this pathway in JEV neuroinvasion remains unknown. Infectious viral particles are usually detected in the brain as early as 5 days after intraperitoneal inoculation.
Intraperitoneal inoculation of the low-neuroinvasive JEV strain SA-14 generates a more marked inflammatory response than that of intracerebral inoculation, 42 and intranasal inoculation of this strain does not lead to fatality, 11 suggesting the route of inoculation may influence the pathological phenotype. Mice that received intraperitoneal inoculation with the neuroinvasive strain JaTH160 have a prominent infiltration of T cells that are mainly CD8+ and have the Th1/Tc1 phenotype in the brain. 30 Activation of microglial cells may play a significant role in the development of Japanese encephalitis in mice by producing proinflammatory mediators (ie, inducible nitric oxide synthase, cyclooxygenase-2, monocyte chemotactic protein-1, interleukin-6, tumor necrosis factor–alpha) and inducing neuronal death. 35 The protective role of inducible nitric oxide synthase during JEV infection has been suggested on the basis of results from a mouse model. 99
Peripheral organs, which include the lung, liver, spleen, heart, and kidney, are possible sites for viral replication after peripheral inoculation. 47 However, little is known about the pathological changes in these extraneural tissues during infection with JEV. After intraperitoneal inoculation, the nonneuroinvasive JEV strain 78668A replicates first in peritoneal macrophages; the viral antigen then appears in the splenic macrophages of the perifollicular region on day 3 postinfection and later in cells of the periarteriolar lymphoid sheath. 68 Productive infection occurs both in macrophages and in T cells. Morphologically, the spleen shows proliferative changes with an enlarged periarteriolar lymphoid sheath, which is followed by the appearance of a prominent germinal center and an increase in the number of macrophages and polymorphonuclear leukocytes in the perifollicular region. 68
Transplacental JEV infection in swine causes fetal encephalitis, abortion, or stillbirth. Pregnant mice inoculated intraperitoneally with the 78668A strain transmit the virus to the fetus, which results in an increased incidence of abortion. 66 Pregnant mice infected with this strain for 6 months transmit the virus to fetuses, which is indicative of the establishment of latent infection in mice. 66 The virus can be reactivated from latently infected mice via cyclophosphamide treatment or via induction of pregnancy. 67 Latent infection in T cells of infected mice was demonstrated using the cocultivation technique. 69
Other Animal Models of JEV
As described in the previous section, older mice are more resistant than younger mice to the lethal effects of JEV administered peripherally 46 and, to some extent, the lethal effects of JEV administered intracerebrally. 38 However, the age-dependent resistance to intracerebral infection is more striking in rat models, 26,82 in which the resistance is closely associated with neuronal maturation. JEV binds to and infects developing neurons among the neural cells in primary fetal rat brain cultures, which suggests that specific molecules that strongly bind JEV may be expressed on the surface of neurons at certain stages of development. 52 Several studies have reported the possible involvement of host molecules—such as glycosaminoglycans, 114,125 a 57-kDa molecule derived from BHK-21 cells, 15 a 74-kDa molecule derived from Vero cells, 51 and a 74-kDa heat shock cognate protein 70 derived from C6/36 cells 95 —in the attachment and/or entry of JEV into cells. A recent study demonstrated that the heat shock protein 70 serves as a putative receptor for JEV in the mouse neuronal cell line Neuro2a. 18 The relationship, if any, between these molecules and the age-dependent resistance to infection remains unclear. It has also been reported that suckling rats intracerebrally inoculated with JEV developed Parkinson’s disease 12 weeks after infection because of the selective depletion of tyrosine hydroxylase–positive neurons in the substantia nigra. 83 This experimental finding is consistent, to some extent, with the clinical findings indicating that a fraction of patients with Japanese encephalitis may have lesions predominantly in the substantia nigra and may manifest Parkinsonian features after recovery from acute encephalitic illness. 92
Several review articles have reported that rabbits and guinea pigs develop asymptomatic infection regardless of the route of inoculation. Additionally, hamsters reportedly die after intracerebral or intranasal inoculation but develop asymptomatic viremia after peripheral infection. 40,78
The experimental intracerebral challenge of monkeys using JEV leads to uniformly lethal encephalitis. 74 Intradermal inoculation of JEV into rhesus monkeys does not cause fatal encephalitis, although the infected monkeys develop viremia. 75 In contrast, Taiwan macaques (Macaca cyclopis) experimentally infected with JEV via an intranasal route develop viremia, followed by clinically evident encephalitis and death. 56 Harrington et al 41 described the pathological features of encephalitis in rhesus (Macaca mulatta) and cynomolgus (Macaca fasicularis) monkeys intranasally inoculated with JEV. The neuropathology of rhesus and cynomolgus macaques is similar. Diffuse meningeal and perivascular lymphocytic infiltrations were observed both in the brain and in the spinal cord. Neuronal degeneration, necrosis, and neuronophagia were most common in the spinal cord and medulla of these animals but were also present in other CNS areas. Glial cell proliferation was pronounced in the gray matter of the spinal cord, brainstem, and thalamus. In contrast, the olfactory lobes were normal, with the exception of the presence of minimal meningeal lymphocytic infiltrates and occasional perivascular cuffing. Myint et al 77 reported the distribution of viral antigens in the CNS of rhesus monkeys infected intranasally with JEV. The JEV antigen was found in the cytoplasm of neurons but not in glial cells, meningeal cells, and vascular endothelium. The JEV antigen was disseminated throughout the CNS, with greater intensity in the thalamus, brainstem, and cerebellum. The preferential localization of JEV antigens in the thalamus and brainstem was also reported in cases of human Japanese encephalitis. 21,48 Infection of Purkinje and granule cells of the cerebellum of monkeys with JEV is similar to what was described by Desai et al for human cases; 21 in contrast, a study performed by Johnson et al with human cases did not identify infection of Purkinje cells. 48 The widespread pattern of lesion and virus distribution observed in the CNS of the macaque models suggests the hematogenous spread of the virus after intranasal inoculation; however, the possible transneural spread of the virus via the olfactory neurons, the trigeminal nerve, and other nerves involved in the development of the CNS lesions should also be considered. The JEV strain that exhibits relatively low neurovirulence and neuroinvasiveness does not reportedly cause encephalitis in monkeys via intranasal challenge; however, intracerebral delivery of this strain causes lethal encephalitis. 74 The rhesus macaque has been proposed as a potential model for the evaluation of vaccines 73,93 and antiviral treatments. 34,41
Summary
Studies of rodent models have provided a body of information about flaviviral pathogenesis, including the identification of genetic determinants and immune reactions that influence disease outcome. Recent developments in the generation and characterization of genetically engineered mice have rendered mouse models a more powerful tool for the dissection of flaviviral pathogenesis. Although intradermal or subcutaneous inoculation mimic the natural route of viral inoculation and subsequent systemic dissemination, a variety of viral and host factors influence the course and outcome of infection. The elucidation of the molecular mechanisms that underlie the influence of host age on disease outcome remains one of the important issues of this field of research. Regarding viral neuroinvasion, the mechanisms via which viruses cross the blood–brain barrier warrant additional investigation. Intracerebral inoculation into suckling mice or hamsters is a more sensitive system for the purpose of infectivity assays and virus isolation. WNV and JEV are strongly neuronotropic, in rodents and in humans, and infection can cause nonsuppurative polioencephalitis or polioencephalomyelitis. Basic histological findings in the CNS lesions observed in WNV-infected mice are similar to those described in JEV-infected and other encephalitogenic flavivirus-infected mice; hence, it is difficult to microscopically differentiate these lesions. A precise diagnosis should be based on serological tests or via the isolation and identification of the virus involved in each case. The neuropathology of macaques intranasally inoculated with JEV resembles that of humans with encephalitis that results from naturally acquired infection, whereas intradermal or subcutaneous inoculation of WNV and JEV into macaques generally results in asymptomatic infection. In view of the close genetic and immunological relationships between nonhuman primates and humans, the macaque models provide useful information that can be used for the evaluation of vaccines and antiviral therapies.
Footnotes
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The original work of the authors was supported in part by Grants-in-Aid for Scientific Research (C) (TK), for Challenging Exploratory Research (TK), and from the Program of Founding Research Centers for Emerging and Reemerging Infectious Diseases (TK, MO, and HS) from the Ministry of Education, Culture, Sports, Science and Technology, Japan.