
Abstract
Recurrent outbreaks of sudden death and bloody diarrhea were reported in March 2013 and February 2014 in a breeding colony of Papillon dogs. During the first outbreak, 1 adult dog and 2 eight-month-old puppies died. During the second outbreak, 2 ten-week-old puppies died. One puppy from the first outbreak and 2 puppies from the second outbreak were examined at necropsy. Histologically, all 3 puppies had severe segmental crypt necrosis of the small intestine and marked lymphoid follicle depletion in the spleen and Peyer’s patches. Real-time (RT) polymerase chain reaction (PCR) demonstrated abundant canine parvovirus (CPV-2) DNA (Ct<15) in the affected small intestine, and immunohistochemistry detected large amounts of CPV-2 antigen in intestinal crypt epithelium and Kupffer cells but few positive macrophages in lymphoid organs. All puppies had marked sinusoidal histiocytosis and multifocal granulomatous inflammation in mesenteric lymph nodes and spleen, prompting additional RT-PCR testing for canine circovirus 1 (CaCV-1). Very high levels of CaCV-1 DNA (Ct<13) were detected in small intestine, lymph nodes, and spleen. In situ hybridization for CaCV-1 detected rare positive nuclei of regenerating crypt epithelium but abundant amounts of CaCV-1 nucleic acid in the cytoplasm and nuclei of histiocytes in all lymphoid tissues, including granulomatous inflammatory foci and hepatic Kupffer cells. Significant levels of CaCV-1 DNA were detected in blood and serum (Ct as low as 13) but not feces from 3 surviving dogs at 2 months or 1 year after the outbreak, respectively. We hypothesize that CPV-2 infection predisposed dogs to CaCV-1 infection and ultimately resulted in more severe clinical disease.
Keywords
Circoviruses are nonenveloped, icosahedral viruses with a single-stranded circular DNA genome of ≈2 kb, classified under the genus Circovirus within the family of Circoviridae. 17,25 Currently, this genus consists of 11 recognized species, including porcine circovirus 1 (PCV-1), porcine circovirus 2 (PCV-2), canary circovirus (CaCV), beak and feather disease virus (BFDV), and other viruses of domestic and wild birds. 26,29,30
Porcine and avian circovirus infections are both characterized by clinical courses that may vary from asymptomatic infections to lethal disease. Circovirus infection in psittacine birds has been associated with dystrophy within the epidermis of the feathers and beak, as well as hyperplasia and necrosis of the integument, lymphoid depletion, and immunosuppression, resulting in psittacine beak and feather disease (PBFD). 28 In pigs, PCV-2 is the primary causative agent of several syndromes collectively known as porcine circovirus-associated disease (PCVAD). 1,5,20 Due to the common presence of PCV-2 nucleic acid in healthy pigs, PCV-2 is considered the causative agent of clinical disease when immunohistochemical or in situ hybridization (ISH) analysis demonstrates large amounts of PCV-2 antigen or nucleic acid in the cytoplasm of macrophages and dendritic cells in the depleted lymphoid follicles or foci of granulomatous inflammation. 12,24 Experimental and natural disease studies have indicated that PCV-2–associated diseases are often accelerated or exacerbated by concurrent viral or bacterial infections. 2,4,6,10 The stimulation of the immune system and induction of a proliferating target cell population for subsequent cirvoviral infection have been shown to be the pivotal effect for the development of circoviral disease in pigs. 13,27
Recently, canine circovirus 1 (CaCV-1, also reported as dog circovirus, DogCV), the second mammalian circovirus species, was identified in dogs. 9 A cohort study 15 using real-time polymerase chain reaction (PCR) revealed that CaCV-1 was detected in the feces from 14 of 204 healthy dogs (6.9%) and 19 of 168 dogs with diarrhea (11.3%). Among the dogs with diarrhea, 68% of those positive for CaCV-1 were coinfected with more than one enteric pathogen, including canine enteric coronavirus, Cryptosporidium spp, Clostridium perfringens α toxin, Giardia spp, Salmonella spp, Campylobacter jejuni, or Campylobacter coli. In addition, CaCV-1 nucleic acid was also found in 3.3% (16/480) of tested blood samples, a prevalence similar to that reported in the previous study. 9,15 Up to 11% of dogs with thrombocytopenia were PCR positive for CaCV-1 in peripheral blood.
CaCV-1 has been detected in a small number of cases of dogs with vasculitis, histiocytic inflammation, or a combination thereof. 3,15 According to the report published by Li et al, 15 tissues from all CaCV-1–infected animals revealed fibrinonecrotizing vasculitis. The location and severity of affected vessels varied among the cases, with most dogs presenting with abnormalities in the intestine and kidneys. Similar to circovirus infection in other species, lymph node necrosis was also observed in several cases, and all dogs had histiocytic infiltrates into lymphoid organs. Similar to reports of PCV-2 in pigs with PCVAD, CaCV-1 was most abundantly present in primary and secondary lymphoid tissues in affected dogs. 15 However, neither viral inclusions nor multinucleate giant cell formation, which are distinct histologic features of PCV-2 infection, were detected by routine histology in dogs infected with CaCV-1.
While there have been numerous anecdotal claims of CaCV-1 causing severe illness and mortality in the Midwestern United States, there have been no confirmed reports of CaCV-1 as the cause of a severe disease outbreak. However, CaCV-1 has been detected in a number of dogs both with and without clinical signs. 9,15 Considering the complexity of porcine circoviral diseases and the difficulties in elucidating the pathogenesis of PCV-2, it is essential to determine the pathogenic potential of CaCV-1 and to shed light on its potential role in canine disease complexes. The study presented here describes a recurrent outbreak of severe diarrhea and death associated with a concurrent infection with canine parvovirus (CPV-2) and CaCV-1 in a CPV-2 vaccinated papillon breeding colony in Michigan and investigates the association of both viruses with the pathologic findings.
Materials and Methods
Two severe disease outbreaks occurred in a Papillon breeding colony in Michigan in March 2013 and February 2014. During the first episode, 2 of 16 adult dogs and all 3 eight-month-old puppies developed anorexia, vomiting, and severe bloody diarrhea. All dogs had been up to date on routine vaccinations, including CPV-2. One of the 2 sickened adults and 2 of 3 sickened puppies died within 1 week of developing clinical signs. One puppy was submitted for necropsy. During the second outbreak, 3 of 4 ten-week-old puppies but none of the 11 adult dogs developed similar disease signs. Two of the affected puppies died and were submitted for necropsy. All 11 adult dogs had been in the breeding colony during the first outbreak. One of those adult dogs had previously developed clinical signs but survived.
A total of 3 puppies had been submitted for necropsy to the Diagnostic Center for Population and Animal Health (DCPAH), Michigan State University. Fresh tissues obtained from the lung, liver, spleen, and intestine were collected and submitted for virology and bacteriology testing. All major organs were collected for histopathologic examination and fixed in 10% neutral buffered formalin, paraffin-embedded, sectioned at a thickness of 5 μm, and stained with hematoxylin and eosin (HE).
In March 2014, blood and feces were collected from those puppies from the first or second disease outbreak that had survived. At the time of collection, 1 dog was 20 months old while the other 2 puppies were 3 months old.
Detection of CPV-2 and CaCV-1 DNA by Real-Time PCR
The presence of CaCV-1 DNA in the samples was analyzed by real-time PCR, according to the procedure described in Li et al, 15 with slight modifications. Sample DNA was extracted with the QIAGEN DNeasy Tissue Kit (Qiagen, Valencia, California). The target for amplification was a 66-bp fragment of the CaCV-1 replicase gene. The following probe and primers were used: probe, 6-FAM-5′-TCCGGAGATGACCACGCCCC-3′-BHQ-1; forward primer, 5′-CTT GCG AGA GCT GCT CCT TAT AT-3′; and reverse primer, 5′-CTC CAC TTC CGT CTT CCA GTT C-3′. The QIAGEN Taq Master Mix (Qiagen) was used according to the manufacturer’s protocol. Real-time PCR was performed using a Smartcycler II instrument (Cepheid, Sunnyvale, California). The cycling conditions were predenaturation at 95°C for 4 minutes, followed by 45 cycles of denaturation at 95°C for 15 seconds (step 1) and annealing and elongation at 60°C for 30 seconds (step 2). Fluorescence signals were collected at step 2. Nuclease-free water was used as the negative control template.
The detection of CPV-2 DNA was performed by SYBR Green–based real-time PCR, as previously described, with slight modifications. 16 The primer pair, (forward) 5′-AGC TGA AGG AGG TAC TAA CTT-3′ and (reverse) 5′-GCC TCA AAA GAA TAA TAT GGT-3′, targets a 153-bp segment of the VP2 gene. The QuantiTect SYBR Green PCR Kit (Qiagen) was used for the PCR reactions. The CFX96 real-time system (Bio-Rad Laboratories, Hercules, California) was used for PCR amplification with the following cycling parameters: predenaturation at 95°C for 15 minutes, followed by 40 cycles of 94°C for 30 seconds, 53°C for 30 seconds, and 72°C for 30 seconds. A post-PCR melt curve analysis was performed to determine the amplicon melting temperature. This step consisted of raising the incubation temperature from 55°C to 95°C in 0.5°C increments every 10 seconds for 80 cycles. A positive amplicon was expected to melt at 74.5°C to 75.5°C, as observed with the CPV-2–positive control DNA.
Double Labeling for CPV-2 Antigen and CaCV-1 Nucleic Acid
Serial sections of formalin-fixed, paraffin-embedded tissues were first used for immunohistochemistry (IHC) for CPV-2 and subsequently for in situ hybridization (ISH) for CaCV-1. IHC labeling was performed using a mouse monoclonal anti-canine/feline parvovirus antibody (CPV1-2A1; Custom Monoclonals International, Sacramento, California), applied for 30 minutes at a 1:400 concentration at room temperature, as previously described. 22 Sections were processed with the Autostainer Link 48 (Dako North America, Carpinteria, California) using the EnVision Flex+ detection system (Dako North America), and the immunoreaction was visualized with 3,3′-diaminobenzidine substrate (Dako North America) as previously described. 16 Deparaffinization and antigen retrieval of formalin-fixed, paraffin embedded tissues were performed on the PT Link (Dako North America), using the Dako Target Retrieval Solution, proteinase K for 15 minutes (Dako North America). Sections of small intestine with CPV-2 lesions that had tested positive by PCR served as positive controls. For the negative controls, the primary antibody was replaced with Tris-HCl buffer. Slides were rinsed and transferred directly to the automated ISH stainer.
In situ hybridization for CaCV-1 for formalin-fixed, paraffin-embedded tissue was developed on an automated slide-processing system (Discovery XT; Ventana Medical Systems, Tucson, Arizona) following a previously published approach. 18 A digoxigenin-labeled oligonucleotide probe (DIG-5′-CCA ACC AAT GAT CCT CAG ACA GAG ACA CCG TTG CTA TGG AAC TTT GAC C-3′) was designed on the replicase gene sequence of CaCV-1 (GenBank accession no. JQ821392.1). Hybridized viral nucleic acid was detected using an anti-digoxigenin antibody (Invitrogen Corporation, Frederick, MD) and a streptavidin-alkaline phosphatase detection system (UMap BLUE anti-Rb AP 760-4314) with a BlueMap NBT/BCIP substrate kit (ChromoMap Blue 760-161; Ventana Medical Systems). For negative controls, the oligoprobe was replaced with a canine herpesvirus probe. Sections of small intestine that had tested positive by real-time PCR with very low Ct values served as positive controls. Slides were counterstained with hematoxylin and coverslipped.
Results
All 3 puppies had similar gross and microscopic lesions. Grossly, the serosal surface of the small intestine was diffusely dull and granular, and there were segmental areas of serosal reddening with few yellowish-white, friable, fibrin strands loosely attached to the serosa. The lumen of the small intestine contained variable amounts of watery, red-tinged, foul-smelling content. The Peyer’s patches were red and slightly depressed below the mucosal surface. The large intestine contained pasty, reddish brown digesta, and the mucosal surface had multiple petechiae.
Histologically, the small intestine exhibited severe segmental mucosal collapse and villar shortening due to extensive loss of overlying enterocytes and also necrosis of the intestinal crypts (Fig. 1a). There was also marked lymphoid necrosis of the Peyer’s patches in the ileum, the mesenteric lymph node, and the spleen, characterized by marked lymphocytolysis of lymphoid follicles (Fig. 3a). In addition, there was multifocal granulomatous inflammation composed of aggregates of epithelioid macrophages within the Peyer’s patches of the ileum, within the mesenteric lymph nodes (Fig. 4a), and in the spleen. The medullary sinus of the mesenteric lymph nodes contained moderate to large numbers of histiocytes, representing medullary histiocytosis. All other organs were histologically unremarkable.
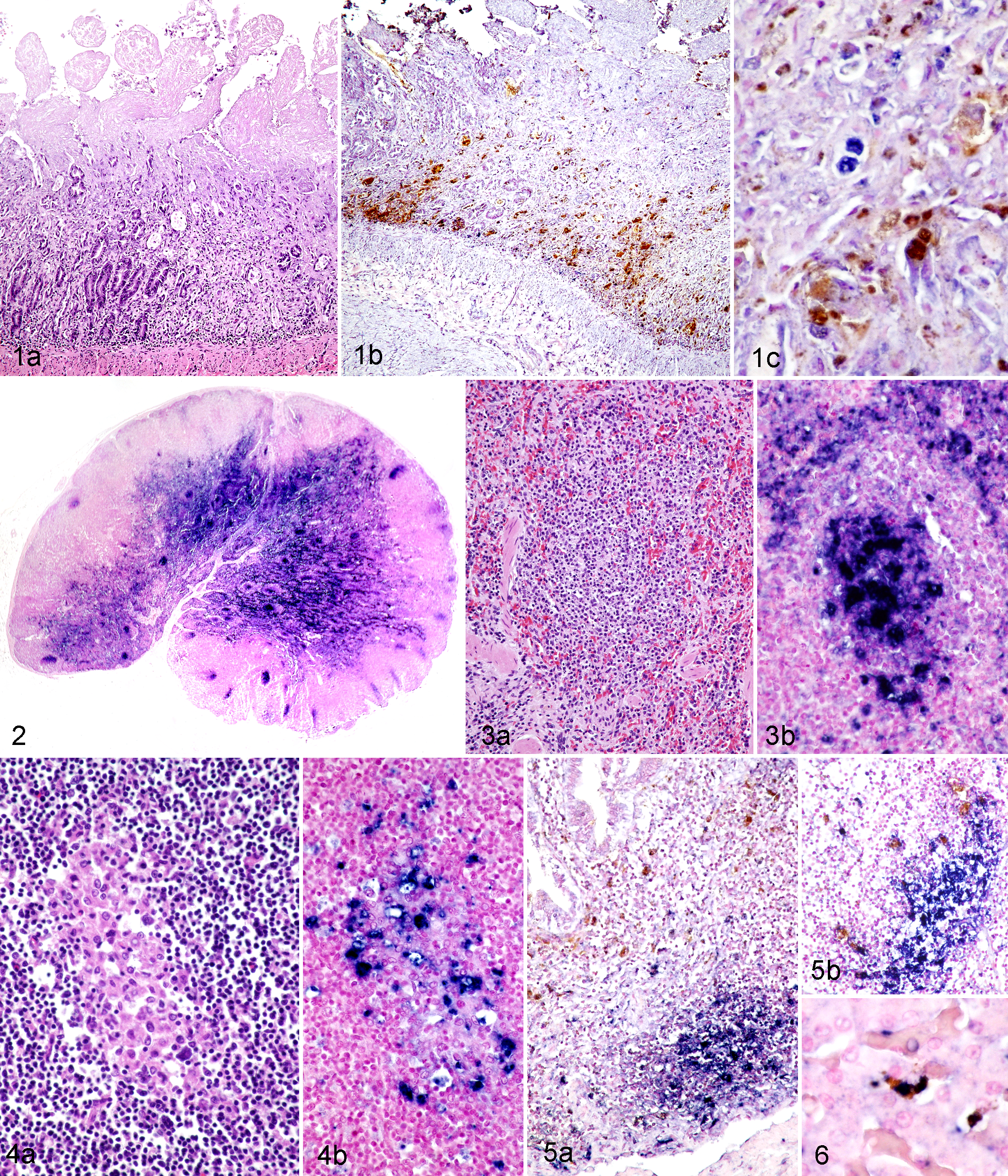
Dual infection of dogs with canine parvovirus 2 (CPV-2) and canine circovirus 1 (CaCV-1). Detection of CPV-2 antigen by immunohistochemistry (IHC, brown chromogen) and CaCV-1 nucleic acid by in situ hybridization (ISH, blue chromogen).
PCR analysis of intestinal samples detected large amounts of CPV-2 (Ct<15) and CaCV-1 DNA (Ct<13) in all 3 dogs. Immunohistochemistry identified large amounts of CPV-2 antigen within the necrotic crypt epithelium of all dogs (Fig. 1b), but viral antigen was rarely detected elsewhere (Fig. 2). Few dendritic cells in mesenteric lymph nodes, Peyer’s patches, and spleen contained intracytoplasmic CPV-2 antigen (Figs. 3b, 4b, 5a, 5b). Phagocytosed CPV-2 antigen was also detected in the cytoplasm of a few Kupffer cells in the liver (Fig. 6). Large amounts of CaCV-1 nucleic acid were detected within lymphoid tissues. Mesenteric lymph nodes were strongly positive for CaCV-1 nucleic acid in follicular centers, marginal sinuses, and medullary sinuses (Fig. 2). A similar pattern of follicular center and marginal sinus labeling was detected in the spleen (Fig. 3b) and Peyer’s patches. Epithelioid macrophages within the previously described multifocal areas of granulomatous inflammation were also strongly positive for CaCV-1 by ISH (Fig. 4b). In contrast, in the small intestinal mucosa, only few nuclei of regenerating crypt epithelial cells labeled positive by ISH for CaCV-1 (Fig. 1c). In the liver, CaCV-1 nucleic acid was detected within the nuclei of the Kupffer cells (Fig. 6). All other organs examined were negative for CaCV-1 and CPV-2 by ISH or IHC, respectively. No pathogenic bacteria were isolated from sections of small intestine.
Feces collected from the 3 dogs that had survived the different clinical disease episodes were negative for CaCV-1 by PCR but positive for CPV-2 (Ct values ranging from 20–25). There were also low amounts of CPV-2 (Ct values ranging from 32–37) detected in the whole blood and serum of all 3 dogs, but surprisingly, very low Ct values for CaCV-1 (Ct values ranging from 13–30) indicated a large viral load in whole blood and serum, regardless of the prior disease status.
Discussion
Here we report a recurrent outbreak of a dual infection of CaCV-1 and CPV-2 in a Papillon breeding colony. Concurrent infection was confirmed by PCR, and the presence of viral antigens/nucleic acids within the lesions was verified by IHC or ISH, respectively. CPV-2 antigen was consistently identified within necrotic crypts of the small intestine. The large amounts of CPV-2 antigen in the necrotic crypts are characteristic for severe canine parvoviral disease and most likely represent the underlying cause for the reported clinical presentation of vomiting and bloody diarrhea in these outbreaks. While CPV-2 also infects lymphocytes and causes lymphocytolysis, only small amounts of CPV-2 antigen were detected in lymphoid tissues and mainly in dendritic cells rather than in areas of necrosis and not in foci of granulomatous inflammation. The absence of CPV-2 antigen could also reflect a time point in the course of the disease, and viral antigen may have been cleared after causing severe lymphoid necrosis. The large amounts of CaCV-1 nucleic acid in areas of lymphoid necrosis and within epithelioid macrophages within foci of granulomatous inflammation support a hypothesis of CaCV-1 causing those lesions in this outbreak rather than CPV-2. Granulomatous inflammation has not been reported as a characteristic microscopic lesion of dogs infected with CPV-2 but is a hallmark of circoviral disease in other species like pigs with postweaning multisystemic wasting syndrome (PMWS).
Little is known about the clinical significance or pathogenesis of CaCV-1 infections in dogs. Virus has been detected in both symptomatic and asymptomatic dogs, and only vasculitis and histiocytic inflammation have been associated with CaCV-1 DNA in some dogs. 15 We consistently identified CaCV-1 in foci of granulomatous inflammation and lymphoid necrosis similar to what has been reported in pigs with PCVAD. 6,12 However, the clinical significance of this finding is unknown. CaCV-1 could have simply replicated in lymphoid tissues of dogs that had previous pathology caused by parvoviral infection without any clinical affect. A more likely hypothesis is a synergistic effect of the CaCV-1 infection that allowed development of clinical disease in dogs despite up-to-date vaccination against CPV-2. A synergistic relationship between circovirus and parvovirus was first reported in pigs, where dual infections with PCV-2 and porcine parvovirus (PPV) were shown to reproduce clinical disease consistent with PMWS. 2,8,11,14 While dual infection with circovirus and parvovirus may represent a common coinfection that causes clinical disease in different species, nonspecific immunostimulation is sufficient to cause severe circoviral disease in pigs. 17 It has been speculated that induction of host cell proliferation is the main requirement for S-phase–dependent circovirus replication, 7,27 and the trigger mechanism for such proliferation can be highly variable. Furthermore, coinfections with other viruses may interfere with the ability of infected animals to clear circovirus through altered cytokine production and impaired interferon-mediated innate immune response. 19 We speculate that infection of CPV-2, which first resulted in crypt epithelial cell and lymphocyte necrosis and subsequently led to proliferation of regenerating epithelial cells and lymphoblasts, provided the necessary target cells for CaCV-1 replication in these outbreaks, ultimately resulting in more severe or prolonged clinical disease. Alternatively, primary infection with CaCV-1 may cause immunosuppression and thus allow concurrent or secondary CPV-2 infection, despite previous vaccination. The tissue distribution of CaCV-1 and CPV-2 in these 3 cases better supports the first hypothesis. Nevertheless, experimental reproduction of such dual infection is required to further elucidate the true significance of CaCV-1 infections in dogs.
To investigate CaCV-1 shedding and possible mechanisms of transmission, we screened feces and whole blood as well as serum for CaCV-1 DNA by real-time PCR. CaCV-1 DNA could be detected in a single dog for more than 1 year postrecovery from clinical disease. This finding is consistent with the viral shedding pattern of PCV-2 in pigs where virus may be present in high concentration in serum as well as nasal and fecal excretion for extended periods of time following natural infection of a population. 23 Transmission of PCV-2 occurs most commonly through direct contact with infected animals or indirect contact with contaminated secretions or excretions, since circoviruses are highly resistant within the environment. 21 The recurrence of clinical disease in this breeding colony could stem from persistent infection either within the infected animals or reinfection from the environment. Shedding of virus by animals that did not develop clinical disease may be due to individual levels of protective immunity and could present a significant epidemiological risk.
Dual infection with CaCV-1 and CPV-2 caused recurrent outbreaks of severe diarrhea and death that clinically resembled canine parvoviral disease in a dog breeding colony, despite up-to-date vaccination against canine parvovirus infection. CaCV-1 was consistently associated with granulomatous inflammation and lymphoid necrosis in different lymphoid organs of 3 examined dogs, while CPV-2 was primarily present in necrotic crypt epithelium. We speculate that infection with CPV-2, which caused crypt epithelial cell and lymphocyte necrosis and subsequent proliferation of regenerating epithelial cells and lymphoblasts, provided the necessary target cells for CaCV-1 replication in these outbreaks and ultimately resulted in more severe and prolonged clinical disease. Alternatively, primary infection with CaCV-1 may cause immunosuppression and thus allow concurrent or secondary CPV-2 infection, despite previous vaccination. In-depth studies of the pathogenic potential of CaCV-1 are needed to fully elucidate the clinical significance of an infection in dogs and to provide proper guidance regarding the necessity of future diagnostic test development, CaCV-1 surveillance, quarantine requirements in shelter and breeding colony environments as well as veterinary clinics, and ultimately targeted efforts for vaccine development.
Footnotes
Acknowledgements
We thank the papillon breeder and veterinarian for sample collection and providing complete history, the histology section for the IHC and ISH, and the virology section for the PCR-based testing.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.