
Abstract
Canine infectious respiratory disease is a common, worldwide disease syndrome of multifactorial etiology. This review presents a summary of 6 viruses (canine respiratory coronavirus, canine pneumovirus, canine influenza virus, pantropic canine coronavirus, canine bocavirus, and canine hepacivirus) and 2 bacteria (Streptococcus zooepidemicus and Mycoplasma cynos) that have been associated with respiratory disease in dogs. For some pathogens a causal role is clear, whereas for others, ongoing research aims to uncover their pathogenesis and contribution to this complex syndrome. Etiology, clinical disease, pathogenesis, and epidemiology are described for each pathogen, with an emphasis on recent discoveries or novel findings.
Keywords
For many years, canine infectious respiratory disease (CIRD), also known as “kennel cough,” was considered a common disease syndrome of limited clinical significance that was largely controlled through routine vaccination. Within the last decade there has been a resurgence of interest in canine respiratory pathogens, sparked either through perceived ineffectiveness of vaccines at controlling respiratory disease outbreaks, particularly within large dog populations such as rehoming kennels, or through the emergence or reemergence of pathogens that result in more severe clinical signs or even fatalities.
CIRD, as with similar syndromes in cattle (bovine respiratory disease complex) and pigs (porcine respiratory disease complex), is increasingly being considered as a complex infection with a multifactorial etiology. Clinical signs such as coughing, nasal discharge, and dyspnea are now rarely associated with a single pathogen and more often are attributed to multiple agents that act sequentially or synergistically to cause disease. Canine parainfluenza virus (CPIV), 6 canine adenovirus type 2 (CAV-2), 36 and canine herpesvirus 1 (CHV-1) 61 have traditionally been considered the main viral causes of CIRD, often in conjunction with or preceding Bordetella bronchiseptica infection. Several multivalent vaccines offer protection against these known pathogens; however, outbreaks of respiratory disease occur in dogs despite routine vaccine use, suggesting a role for other pathogens in CIRD.
It was during an investigation into one of these large outbreaks of respiratory disease at a rehoming kennel in London that canine respiratory coronavirus (CRCoV) was first isolated in 2003. 47 In the following years, a number of other novel or reemerging pathogens have been implicated in CIRD, including canine pneumovirus, canine influenza virus, pantropic canine coronavirus, Streptococcus zooepidemicus, and Mycoplasma cynos. Indeed, it is likely that many of these agents were present and causing disease in kenneled dogs much earlier; however, recent advances in pathogen detection, particularly the use of molecular methods, have resulted in their identification. To emphasize this point, a retrospective study of archived tissue revealed the presence of CRCoV in the canine respiratory tract as early as 1996. 41 The routine use of vaccines and antibiotics may also have exerted a selective pressure on viruses and bacteria, respectively, such that the relative prevalence of various canine respiratory pathogens has changed with time. For example, CPIV continues to be a prevalent pathogen in CIRD, whereas CAV-2 and CHV-1 are now considered less common pathogens, at least in the UK. 45
Much of the investigation and research into these new and emerging pathogens has been published in microbiology or related journals; therefore, it is now critical that we present this information to veterinary communities. This review summarizes the latest research on these pathogens and their role in canine respiratory disease.
Canine Respiratory Coronavirus (CRCoV)
Etiology
CRCoV belongs to the Betacoronavirus genus of the Coronaviridae family, most closely related to bovine coronavirus (BCoV) and human coronavirus OC43. 46 CRCoV is serologically and genetically distinct from canine coronavirus (CCoV), an Alphacoronavirus, which is typically an etiological agent of enteric disease, although pantropic CCoVs have recently been identified and are discussed later in this review.
CRCoV was first described in 2003 during an investigation into the pathogenesis of CIRD in a large UK rehoming center. 47 This study showed a strong association between exposure to CRCoV and the development of CIRD in dogs entering the kennel. Since the initial report, a number of serological surveys and surveillance studies have been undertaken and the global presence of CRCoV has been demonstrated. 2,3,29,40,57,63,75,86,89,97,110 CRCoV is now considered an important and widespread etiological agent of respiratory disease in kenneled dogs. In 2008, a review of the origins and epidemiology of CRCoV was published; 42 therefore, here we will focus on more recent developments.
Clinical Disease
Collective findings from outbreaks and clinical cases indicate that CRCoV is associated with mild respiratory disease during the early stages of CIRD, with typical clinical signs including a dry cough and nasal discharge, 43,45,47 which are not unique to this agent.
In the first experimental CRCoV infection study to be reported, the pathogenesis of the disease resulting from infection with 5 CRCoV isolates that originated from the UK and USA was investigated. 74 Following intranasal challenge, all 5 isolates gave rise to clinical signs of respiratory disease (nasal discharge, sneezing, and coughing), consistent with findings in kenneled dogs. 45,74 Viral shedding from the oropharynx was readily detected up to 10 days postinfection by reverse transcription polymerase chain reaction (RT-PCR) and 6 days postinfection by virus isolation. 74 CRCoV was also detected and reisolated from a wide range of respiratory tissues and respiratory-associated lymphoid tissues (trachea, palatine tonsil, nasal cavity, nasal tonsil, bronchial lymph node, apical and diaphragmatic lung lobes, and lung lavage fluids). Consistent with findings from kenneled dogs, the trachea and nasal tonsil were the most common sites of infection and were also reported to have the highest viral loads. 73,74 The reisolation of CRCoV from the tissues of experimentally infected dogs with clinical disease fulfilled the final criteria for Koch’s postulates and definitively proved a causal relationship between CRCoV and respiratory disease in dogs.
During an investigation of 10 kenneled dogs with respiratory disease, CRCoV was predominantly detected in respiratory tissues; however, in 2 dogs CRCoV was also detected in the spleen, mesenteric lymph node, and colon. 73 In separate case studies, CRCoV was detected in a rectal swab from a dog presenting with vomiting and diarrhea 110 and in the lung, spleen, mesenteric lymph node, and intestines of a dog that died from hemorrhagic gastroenteritis. 29 In both cases, enteric viral pathogens (CCoV and canine parvovirus [CPV], respectively) were detected and were the most likely causes of disease and death. By RT-PCR, CRCoV was detected in a limited number of rectal swabs from experimentally infected animals. 74 Interestingly, detection was limited to only 4 dogs, 3 of which had been challenged with a CRCoV strain that had not undergone in vitro cell culture passage. This raises the question as to whether there are strain-related differences in tropism or whether cell culture adaptations could account for the lack of CRCoV detected in the rectal swabs collected from dogs challenged with cell culture-grown strains of virus.
The detection of CRCoV in gastrointestinal samples suggests that like some bovine coronavirus strains, 78 CRCoV may have the potential to exhibit a dual tropism. Conversely, attempts to isolate CRCoV from enteric samples have to date been unsuccessful, 29,74,110 and therefore whether CRCoV is able to establish a productive infection in enteric tissues, or whether it simply passes through the gut as a bystander, is unclear. Further investigation of this and the potential clinical consequences is required.
It is not possible to diagnose CRCoV based on clinical signs alone due to the multifactorial etiology of CIRD and the indiscriminant nature of the signs observed compared with those of many other respiratory viruses involved in the disease complex.
Serological assays can be used to determine whether CRCoV is involved in a CIRD outbreak using paired serum samples collected at least 2–3 weeks apart. ELISAs and immunofluorescence assays based on both CRCoV and BCoV antigens have been published, 29,47 and such diagnostic tests are becoming more widely available.
Pathogenesis
Indisputably, CRCoV displays a clear preferential tropism for canine respiratory tissues, particularly those of the upper airways (Fig. 1). Immunohistochemical staining was used to detect CRCoV antigen in the cytoplasm of ciliated epithelial and goblet cells of the trachea (Fig. 2) and bronchioles of experimentally infected dogs. 74 Identical staining was observed in the ciliated epithelial cells of tracheal tissues acquired from naturally infected dogs 41,88 and in an in vitro CRCoV-infected canine tracheal organ culture system. 88
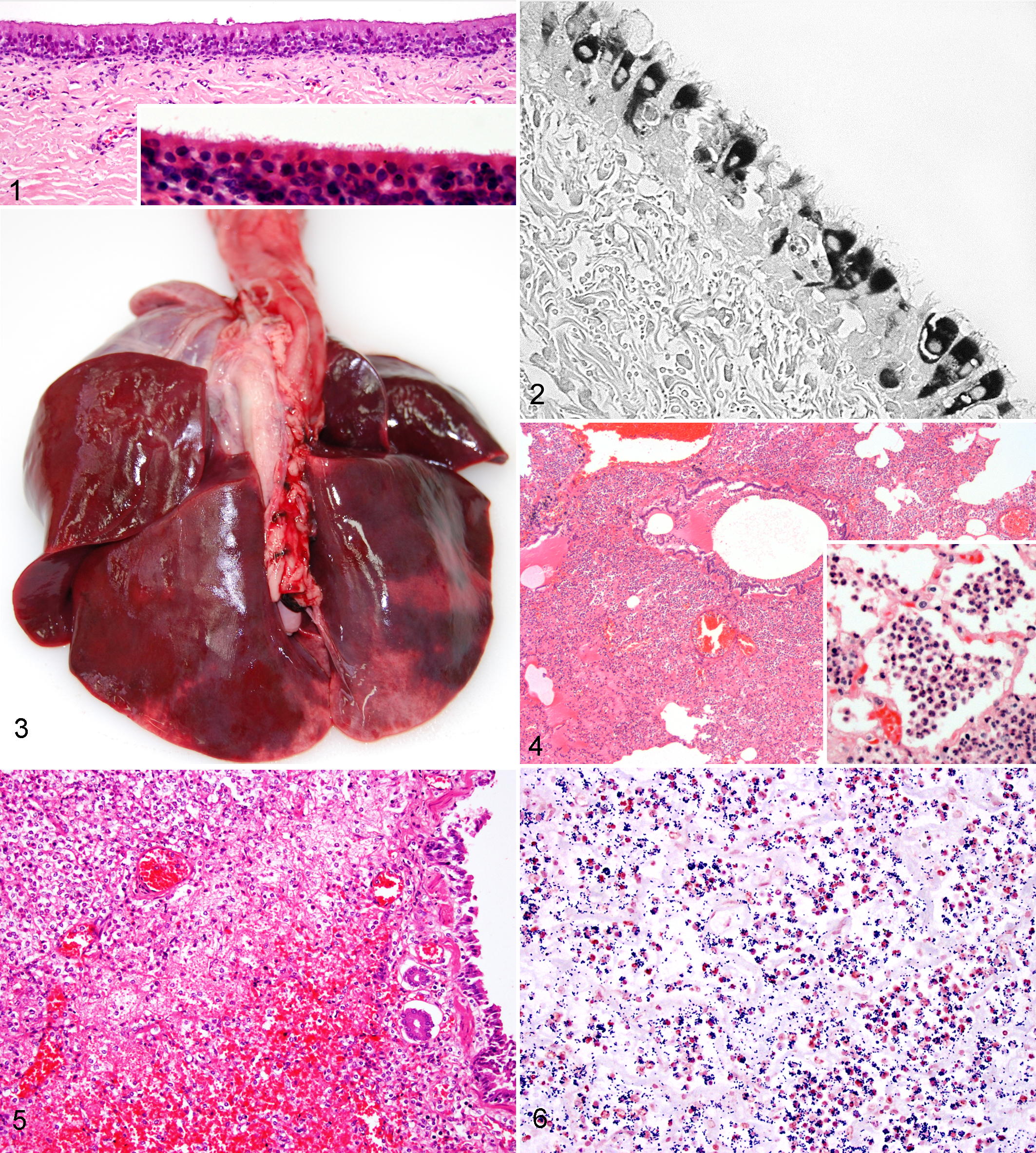
Histological examination of the respiratory tissues from experimentally challenged dogs showed a clear and consistent association between exposure to CRCoV and inflammation in the nares and trachea, with significant injury, such as shortening and clumping, or loss of tracheal cilia. 74 Perivascular and peribronchiolar lymphoid aggregates were observed in the lungs, but changes were less significant and more variable than those within the upper airways. The reported histopathological changes were apparent at the earliest euthanasia time point at 3 days post infection and were still clearly evident at 14 days post infection, although some recovery was noted. 74 As yet there are no published reports detailing the histopathological changes associated with natural CRCoV cases, and given the typically multifactorial nature of CIRD, it would be difficult to attribute any histopathological lesion to a single agent. However, the histopathological changes observed in the experimental study are consistent with those typically associated with the early stages of CIRD, that is, mild to moderate tracheobronchitis. These findings support the hypothesis that CRCoV causes damage to the mucociliary clearance mechanisms of the upper airways, which may predispose dogs to secondary infections. Further evidence in support of this was seen in an in vitro CRCoV-infected canine tracheal organ culture system, in which a moderate reduction in mucociliary clearance was observed. 88
In addition to the histopathological changes in the tissues, significant changes in the leukogram profile of dogs experimentally infected with CRCoV were also reported. 74 As would be expected following viral antigen stimulation, there was a statistically and clinically relevant lymphocytosis at 6–14 days post infection. However, many dogs also had decreased neutrophil concentrations, which in some individuals presented as neutropenia with mild left shift and toxicity, indicative of an acute inflammatory response with a high demand for neutrophils. However, unlike cases of murine and rat coronavirus infections, 9,54 where neutrophils commonly infiltrate infected tissues and are implicated in an immune-mediated pathogenesis, the presence of predominantly lymphoid aggregates in tissues suggests that this does not occur during CRCoV infection. 74
Coronaviruses use multifaceted approaches to counteract host antiviral responses, providing an opportunity for viral replication and dissemination within the host. Other β-CoVs have been shown to suppress the activation of antiviral effector genes, and hence the production of proinflammatory cytokines, by both passive 107 and active 49 inhibition of transcription factor IRF-3. The findings of an in vitro study suggest that CRCoV is also able to inhibit proinflammatory cytokine and chemokine induction during the early stages of infection. In that study, using a CRCoV-infected canine tracheal organ culture system, the messenger RNA (mRNA) levels of 3 key proinflammatory cytokines, interleukin-6, interleukin-8, and tumor necrosis factor-α, were suppressed at 24–48 hours post inoculation. 88 During this time, viral load within the tissue remained stable. At 96 hours, there was an increase in viral load and a corresponding increase in the mRNA levels of all 3 cytokines. The mechanisms used by CRCoV to achieve inhibition require further investigation; at present it is unknown whether there are strain-associated differences and whether these findings are consistent in vivo.
The discovery of CRCoV and the subsequent research into its pathogenesis have highlighted the importance of viruses in CIRD, not only through direct cellular damage but also through the less obvious effects on the host immune system. Combined, these effects can result in significant morbidity while predisposing the animal to other, more serious secondary infections.
Diagnosis
The most suitable and readily available test for CRCoV is RT-PCR. A number of published RT-PCR assays target different regions of the viral genome 47,73,110 and are offered by commercial diagnostic companies. As CRCoV is found most frequently in the trachea and nasal cavity, oropharyngeal and or nasal swabs are the most suitable diagnostic samples. Where postmortem samples are available, nasal cavity, nasal tonsil, trachea, and lung samples should be collected for testing by RT-PCR. Formalin-fixed tissues may also be tested for CRCoV antigen using immunohistochemical detection, 74,88 although this is currently carried out only for research purposes and is not available commercially.
Canine Influenza Virus (CIV)
Etiology
An influenza virus was first detected in racing Greyhounds in Florida in 2004. 25 These dogs had been suffering from periodic outbreaks of respiratory disease of unknown etiology for a number of years prior to the isolation of this virus. Genetically, the virus is most closely related to equine influenza virus H3N8, and indeed all 8 genome segments are of equine origin, suggesting a direct transmission event from horses to dogs possibly around 1999–2000, based on a retrospective serological survey of racing Greyhounds. 4,25,80 What is intriguing about CIV is its ability to transmit directly from dog to dog, and subsequent work has shown that antibodies to the virus are detected widely in the canine population across the USA. 38,80
Pathogenesis and Epidemiology
The virus is most prevalent in group-housed dogs, such as in kennels, and as such can spread rapidly among the immunologically naive animals, affecting all ages. 37 Seropositivity to CIV has been reported to be up to 49% in at-risk dog groups such as those in multidog households or rehoming kennels, and there appear to be geographic differences such as an increased incidence in the northeast USA during 2008. 5
Strictly speaking, CIV refers to infection with H3N8; however, given the worldwide interest and abundance of research into human influenza and the fear of future pandemics, it was only a matter of time before other influenza subtypes were detected in dogs. In 2004, an avian H5N1 virus was detected in a dog that died with severe respiratory signs after eating a duck carcass. 102 In 2009, the so-called H1N1 “swine influenza” hit the headlines, and soon afterward sporadic reports occurred of cases of canine infections with this virus in China 66 and the USA (http://vetmedicine.about.com/b/2009/12/22/first-confirmed-case-of-h1n1-in-a-dog.htm) and serological evidence of infection in Italy. 39 These latter infections are likely to have spread to the dogs from their infected owners and thus are likely dead-end infections. However, the coexistence of multiple types of influenza circulating in dogs, which are capable of cross-species transfer between dogs and humans, should underline the importance of continued vigilance by both the veterinary and medical communities regarding the increased risk of viral genetic reassortment, particularly as many of us have more intimate contact with dogs than with those species traditionally regarded as the “mixing vessels” for reassortment events, such as pigs or poultry. Indeed, to emphasize this point, in 2007 in South Korea a number of outbreaks of respiratory disease in kenneled dogs were attributed to avian H3N2 influenza, 98 and Song et al 100 reported on a novel reassortment between pandemic H1N1 and canine H3N2 in Korean dogs to create a novel H3N1 CIV. Currently, a great deal of active research is underway, particularly in China, examining the role of low pathogenic avian influenza viruses (specifically H3N2 and H5N2) in canine respiratory disease. The seroprevalence of H3N2 among pet dogs is reported to be up to 33% in parts of northeastern China, 114 and more important, in experimental infections the virus has been shown to cause marked necrotizing lesions within the respiratory tract, predominantly the lungs, but also inflammation and necrosis in other organs, including the heart, kidneys, liver, spleen, and brain. 112 Perhaps most worrying is the recent demonstration that H5N2 is capable of dog-to-dog transmission, resulting in generally mild clinical signs of respiratory disease (sneezing, nasal discharge, and mild coughing) but demonstrating that dogs can play a role in transmission and spread of influenza viruses. 101
Clinical Disease
Unlike most other viral respiratory pathogens of dogs, CIV infection usually causes clinical signs in the absence of vaccination and lack of previous exposure. However, the clinical signs are what might be expected from most other viral pathogens, such as mild anorexia, depression, coughing, and nasal and ocular discharge. 35,99 Again, as with the other viral pathogens, infections are rarely uncomplicated, and worsening clinical signs and the development of pneumonia generally suggest the involvement of secondary bacterial infections.
In experimental infections, CIV invariably results in upper respiratory tract lesions with damage to the ciliary epithelium increasing the susceptibility to other pathogens. 17,37 However, CIV infection also seems to result in a greater incidence of lower respiratory tract involvement than most of the other canine viruses. This latter feature distinguishes CIV from CRCoV, which seems to be more restricted in tropism with a clear preference for the tracheal epithelium. 46 Most dogs develop necrotizing and hyperplastic tracheitis and bronchitis with involvement of submucosal glands, in addition to bronchitis and pneumonia. 17 Initially, the pneumonia is characterized by neutrophils filling alveolar spaces and loss of alveolar type 1 epithelial cells. This later progresses to alveolar thickening due to type 2 epithelial cell hyperplasia, and neutrophils are replaced by alveolar macrophages. Interestingly, there does seem to be a greater susceptibility in racing Greyhounds, which to date has not been explained; however, deaths within these dogs from severe pneumonia with pulmonary and thoracic hemorrhage are widely reported. 17 However, within the pet dog population, fatal cases of uncomplicated CIV-pneumonia are rare and thus it is likely, as with the other viral respiratory pathogens, that environmental or host/breed-associated factors are important for the development of the most severe expression of disease. Indeed, in most cases of fatal pneumonia in Greyhounds, a secondary bacterial pathogen, principally Streptococcus zooepidemicus, is often implicated. 17
Diagnosis
RT-PCR is the preferred and most widely available diagnostic method for suspected clinical cases. Given the emergence of a number of different influenza genotypes, a test that covers multiple types, in addition to H3N8, is also recommended.
Canine Pneumovirus (CnPnV)
Etiology
CnPnV was first described in 2010 following a retrospective study of respiratory disease in dogs from 2 animal shelters in the USA. 92 It was reported that after 3 passages in A72 cells, a cytopathic effect uncharacteristic of viruses commonly isolated from dogs was observed. After investigators failed to identify other common agents, a pool of monoclonal antibodies against a human respiratory pathogen (human respiratory syncytial virus) was used, and fluorescent staining was observed in infected cell cultures. Degenerate primers were used to amplify 3 regions of the viral genome, and sequence analysis revealed that the unknown virus was closely related to murine pneumovirus (MPV), a common pathogen of research and commercial rodent colonies. 64
In 2011, the full genome sequence of CnPnV was completed and its place within the pneumovirus genus of the subfamily Pneumovirinae, family Paramyxoviridae, was confirmed. 91 CnPnV is closely related to a number of important human and veterinary respiratory pathogens, including human and bovine respiratory syncytial virus, human metapneumovirus, avian pneumovirus, and, most closely, MPV.
Epidemiology
Since its initial discovery, CnPnV has been detected in kenneled dogs in 8 US states. 91 Although CnPnV is readily detected in dogs with respiratory disease, 91,92 its role as a causative agent of disease has not been properly examined. It will be necessary for extensive serological surveys and surveillance studies to be undertaken to determine the presence of CnPnV globally and to establish an association with clinical disease.
Pathogenesis
The pathogenesis of CnPnV infection in dogs remains to be investigated. For other pneumovirus species, transmission occurs via contact with respiratory secretions, and the apical airway epithelial cells are the principal targets for infection. Indeed, following an experimental inoculation of BALB/c mice, CnPnV was shown to replicate effectively in the lungs, with viral antigen detected predominantly in the epithelial cells lining the bronchioles. 81
The host immune response, in particular neutrophil activity, has been associated with tissue damage during pneumovirus infection. 48,96,109 In the mouse model, CnPnV infection was associated with the induction of a local proinflammatory cytokine response and the presence of mild multifocal perivascular neutrophilic inflammation in the lung. 51,81 Compared with MPV infections, however, there were minimal histopathological changes, reduced neutrophil infiltration of the parenchyma, and no apparent hemorrhage or edema. 51 Similar reductions in neutrophil infiltration have been observed in MPV-challenged mice deficient in type 1 interferon receptors or in the absence of interferon-γ signaling. 11,50 Pneumoviruses have developed many immune modulation strategies, in particular mechanisms for circumventing host type 1 interferon responses. It will be interesting to establish whether these reported findings are consistent in the natural host of the dog and, if so, whether they are linked to interferon signaling and immune-modulatory mechanisms of the virus.
The glycoprotein (G protein) of related pneumovirus species mediates attachment to host cells and as such contains the major neutralizing epitopes. Considered an important virulence factor, a high degree of genetic and antigenic variation is shown to exist within the G protein of RSV species, which frequently gives rise to escape mutants. 103 In a recent study, the G protein sequences of several isolates were reported, which led to the designation of A and B subtypes, both of which were identified in multiple locations across the USA and included 2 strains isolated from cats. 51 The study compared group A and B viruses in the BALB/c mouse model, in which virus replication was detected as early as 3 days post intranasal inoculation (dpi), with peak levels reached at 6 dpi. Local cytokine production and inflammatory responses were examined, and some differential responses between A and B strains were observed. 51 Whether a clinical outcome can be linked to subtype warrants further investigation; however, for human RSV, the findings are conflicting. In addition to in vivo studies, clinical surveillance should be undertaken, and it will be important to establish the degree of variation across the CnPnV genome and in particular the G protein to determine whether variation is associated with pathogenicity in the dog.
Diagnosis
At present the only available test is a quantitative RT-PCR assay offered by the group that first identified this novel virus. 92 The recommended diagnostic samples include nasal and pharyngeal swabs, tracheal wash, and bronchoalveolar lavage fluid. Postmortem samples from the lung and upper airways are also suitable for testing with this assay.
Pantropic Canine Coronavirus
Etiology
Canine coronavirus (CCoV) has traditionally been considered an enteric pathogen of low virulence that is frequently isolated from dogs with diarrhea but is also detected in feces of dogs displaying no clinical signs. 26,104 In 2005, an outbreak of fatal systemic disease occurred in Italy; CCoV was detected in affected dogs in a range of tissues including the lungs, where pulmonary lesions were observed. Although the presenting clinical signs associated with the infection are predominantly gastrointestinal and neurological, its presence in the lungs of infected dogs may suggest a role of this tissue in the pathogenesis of CIRD. To distinguish from cases with a purely enteric tropism, the term pantropic canine coronavirus was applied.
In contrast to canine respiratory coronavirus (a Betacoronavirus), CCoV belongs to the genus Alphacoronavirus and is grouped into 1 viral species with the closely related feline coronaviruses (FCoV) and porcine transmissible gastroenteritis virus (TGEV). CCoV is divided into CCoV type I, which is closely related to FCoV type I, and CCoV type II, which is further divided into types IIa and IIb. 26 CCoV type IIb is closely related to TGEV. 31,44
CCoV type IIa is the most prevalent of the types within the canine population; however, coinfections of CCoV types I and II are also common. 32
Clinical Disease
A pantropic variant of CCoV was first isolated from an outbreak of systemic and fatal disease in dogs in a pet shop in Bari, Italy. The dogs exhibited lethargy, vomiting, hemorrhagic diarrhea, and neurological signs. 13 All 7 affected dogs were between 45 and 56 days of age and died within a few days of onset of clinical signs.
At necropsy, gross findings were hemorrhagic enteritis and serosanguinous abdominal effusion, multifocal consolidation of the lungs, superficial hemorrhages within liver and spleen, petechial hemorrhages on lymph node surfaces, and multifocal hemorrhagic renal infarcts.
Histopathological examination showed a fibrinopurulent exudate within bronchi, bronchioles, and alveoli; degeneration of bronchial and bronchiolar epithelia; fibrinoid vascular necrosis; alveolar hemorrhage; and edema. 111 Further extraintestinal lesions included lymphoid depletion within the spleen and acute renal infarcts. In the lungs, viral antigen was detected within bronchial and bronchiolar epithelial cells and alveolar septa as well as occasional arterial walls. The authors suggest that the vascular lesions observed may have led to the renal infarcts and abdominal effusion.
CCoV type IIa was isolated from the affected tissues and was detected by RT-PCR, whereas other canine viral or bacterial pathogens were not identified. The viral isolate CB/05 was shown to have a high sequence identity with FCoV type II strain 79-1683 within the spike gene. 13
Pathogenesis and Epidemiology
Subsequent experimental infections produced similar clinical signs to those observed during the original outbreak, with a more severe clinical course in 2.5-month-old dogs compared with 6-month-old dogs. 27 Infected dogs also exhibited lymphopenia with cell counts below 60% of the initial count. Further experimental studies showed depletion of CD4+ T-lymphocytes for up to 30 days post infection. 69
Since the original report, further outbreaks have occurred in Italy, 33 France and Belgium, 115 and Greece. 76 Clinical signs exhibited by pups during 5 outbreaks in Belgium and France included vomiting, diarrhea, and convulsions. CCoV was detected by RT-PCR in the lungs of affected dogs in all outbreaks. Pulmonary lesions were described as subacute interstitial pneumonia and alveolar edema. In 4 of the 5 outbreaks, canine parvovirus type 2c was detected in fecal samples of affected dogs. Spleen and liver were tested by PCR for CPV-2c but were found to be negative. The authors suggest that infection with CPV-2c may facilitate systemic spread of CCoV. While the original pantropic isolate CB/05 is a type IIa CCoV, one outbreak in France was associated with CCoV I. 115
A large European surveillance study of 354 cases of fatal disease associated with clinical signs including leucopenia, enteritis, respiratory distress, and/or neurological signs identified CCoV in extraintestinal tissues in 33 cases. 28 Of these, 24 were CCoV type IIa and 9 were CCoV IIb, indicating that systemic infection is not limited to CCoV type IIa and confirming the findings of Zicola et al. 115 In addition, similar to previous studies in France and Belgium, dogs with systemic CCoV infection were commonly coinfected with CPV-2.
Currently available inactivated vaccines against CCoV have been shown to be ineffective against challenge with pantropic CCoV IIa; 30 the clinical signs, however, were milder in vaccinated dogs. To date, no consistent genetic marker has been identified for pantropic CCoV strains.
Diagnosis
CCoV may be detected in fecal samples of clinically healthy dogs, and the seroprevalence is high. 104,106 Therefore, the only definitive test for an infection with pantropic CCoV is the detection of CCoV in extraintestinal tissues by virus isolation in canine or feline cell lines or detection by RT-PCR. Numerous RT-PCR protocols have been published allowing the detection of CCoV 84 as well as quantitation and subtyping; 34 however, other diagnostic tests are unproven and currently not commercially available.
Other Canine Viruses
Canine Bocavirus (CBoV)
Bocavirus is a genus of the Parvovirinae subfamily, of the Parvoviridae family. 55 The genus includes bovine parvovirus and minute virus of canines (MVC) 55 as well as porcine, 22 gorilla, 59 and human bocavirus 1 species, which commonly infect the respiratory and gastrointestinal tracts of young animals and humans. Most of the attention to date has focused on human bocavirus (HBoV) found ubiquitously worldwide, and in particular HBoV-1, which is linked to respiratory disease. Initially, the frequent detection of HBoV-1 in asymptomatic individuals and the high viral coinfection rates detected in symptomatic cases indicated that HBoV-1 may exist only as a bystander. However, the application of improved diagnostics has demonstrated a high seroprevalence and a positive correlation between disease and high copy numbers of HBoV, providing increasing evidence that HBoV-1 is an important respiratory pathogen (reviewed by Jartti et al 56 ).
In a recent metagenomic study, short sequences with a distant relationship to other animal bocaviruses were identified in respiratory samples from dogs. 58 Primer walking techniques extended the available sequence to reveal the presence of a novel bocavirus, CBoV. CBoV is genetically distinct from MVC. MVC typically causes only a subclinical infection in adult dogs but can also cause abortions in bitches and severe respiratory infections in newborn puppies. 10,16
Analysis of 35 CBoV variants indicated that these novel CBoVs represent a highly diverse group of viruses, which can be divided into 3 groups. Group A variants were significantly more prevalent in healthy dogs raised in controlled conditions than in diseased animals, and group B and C variants were substantially more prevalent in dogs with respiratory disease. 58 Parvovirus capsid proteins contain determinants of immunogenicity and host cell tropism, and minor genetic changes can alter the host range and pathogenic potential. 79 It is possible that the increased association of disease with C variants may be the result of a unique deletion in the C-variant VP2 capsid gene; 58 however, this remains to be examined.
A clear relationship between CBoV and respiratory disease in dogs is yet to be established, and furthermore it was not determined whether other viral pathogens were also present in the infected dogs as is frequently the case with HBoV-1. However, the higher prevalence of CBoV B and C variants in dogs with respiratory disease indicates that these viruses may infect diseased animals as opportunistic pathogens or may enhance the severity of lesions in coinfections. 58
Canine Hepacivirus
Recently, a novel virus, tentatively named canine hepacivirus, was identified in respiratory swabs collected from 9 of 33 dogs housed in rehoming shelters in the USA. 60 The dogs represented 5 different outbreaks of respiratory disease. Six canine hepacivirus-positive dogs were identified in one outbreak, and 3 were identified in a second outbreak in the same rehoming shelter 2 weeks later. The virus was also detected in the hepatocytes of liver samples from 5 unrelated dogs that died from gastrointestinal illness. Subsequent attempts to detect canine hepacivirus and antibodies to this virus in canine serum samples have been unsuccessful; 14,67 however, other samples were not examined, and the absence of the virus from the small groups of dogs surveyed is unlikely to represent its true ecology.
Canine hepacivirus is the first nonprimate hepacivirus (NPHV) to be described, and subsequently a closely related virus has also been detected in the serum samples of horses in the UK and USA. 14,67 Comparative phylogenetic analysis has confirmed that canine hepacivirus is the closest genetic relative of hepatitis C virus (HCV) to date. 60 HCV is associated with chronic hepatitis in humans, and the discovery of NPHVs may provide new insights in to its origins. The discovery of canine hepacivirus in dogs with respiratory disease is intriguing; however, a role for this novel virus in the pathogenesis of disease remains to be determined.
Streptococcus zooepidemicus
Etiology
Streptococcus equi subsp zooepidemicus (S. zooepidemicus) is considered to be a commensal organism in the upper respiratory and lower genital tracts of horses but may be associated with opportunistic infections such as abscesses, endometritis, and reproductive failure. 105 S. zooepidemicus has been known to cause sporadic disease in dogs since the late 1970s; however, in recent years several outbreaks of acute and frequently fatal hemorrhagic pneumonia have been described in dogs from a number of countries. 15,19,62,82
S. zooepidemicus is a beta-hemolytic, Lancefield group C bacterium that is distinct from the group G bacteria such as Streptococcus canis, which are more commonly isolated as commensal organisms from dogs. 52 Although there are occasional reports of the bacterium causing upper respiratory tract signs such as chronic nasal discharge and rhinitis, 83 it is more frequently associated with acute hemorrhagic and fibrinosuppurative bronchopneumonia in kenneled and racing dogs. 52
Clinical Disease
Clinically, infected dogs may present initially with signs similar to “kennel cough,” including a moist cough and serous or mucoid nasal discharge. 85 However, pyrexia is also frequently observed and there is rapid progression to depression, anorexia, and dyspnea. Hemorrhagic nasal discharge or hematemesis has frequently been reported. Death can occur within 24–48 hours of the first clinical signs, and occasionally this sudden deterioration is observed without any preceding signs.
Lesions are remarkably consistent in almost all dogs and comprise severe acute (or even peracute) fibrinosuppurative, necrotizing, and hemorrhagic bronchopneumonia with pleuritis (Figs. 3–5). 87 In most cases, numerous bacterial cocci can be observed within infiltrating neutrophils or free within alveolar spaces (Fig. 6). In some cases, possibly representing those presenting earlier, there is fibrinous and/or hemorrhagic pneumonia with very few neutrophils. 87 These latter cases may suggest a significant role for exotoxins, acting as superantigens, which damage the pulmonary vasculature leading to fibrin leakage, edema, and widespread hemorrhage. Hemorrhages are also frequently observed on the pleural surfaces and mediastinum, 87 again suggesting a systemic process mediated by toxins.
Pathogenesis
The pathogenesis of S. zooepidemicus infection in dogs has yet to be fully elucidated; however, a significant role for bacterial exotoxins acting as superantigens seems likely. Certainly canine isolates of S. zooepidemicus have been shown to contain known exotoxin genes similar to those occurring in Streptococcus pyogenes in humans and Streptococcus equi in horses. 77,87 Indeed, the rapid clinical course that characterizes most cases of S. zooepidemicus infection mirrors streptococcal toxic-shock syndrome in humans. Levels of proinflammatory cytokines and chemokine genes, including interleukin-6, tumor necrosis factor-α, and interleukin-8, were shown to be significantly elevated in the lungs of dogs with acute pneumonia, versus those from which S. zooepidemicus was isolated but lacking clinical signs of respiratory disease. This implicates a host-specific overexuberant or uncontrolled host immune response in the pathogenesis of the pneumonia. 87
The role of clinically silent carrier animals is an area of current research because S. zooepidemicus has been isolated from clinically healthy dogs. 19 Indeed, although S. zooepidemicus is capable of causing disease as a primary canine pathogen, Larson et al 65 failed to reproduce clinical disease in healthy dogs experimentally challenged with the bacterium alone. However, when dogs were dual challenged with CIV (H3N8) and S. zooepidemicus, they developed severe necrotizing neutrophilic pneumonia, suggesting that in some situations S. zooepidemicus acts as an opportunistic or secondary pathogen. The histopathological lesions were also much less severe in those dogs vaccinated against CIV prior to dual challenge. This study underlines the complex role of S. zooepidemicus in CIRD and demonstrates how studying a single pathogen in isolation may not fully represent the in vivo situation.
Diagnosis
S. zooepidemicus can be routinely cultured from nasopharyngeal swabs or from the lung in postmortem cases and frequently yields a pure growth in those cases where the characteristic lung changes (described above) are observed. PCR for speciation of streptococci is also possible and, although not widely available as a diagnostic method, is recommended since recent genetic work has shown that canine isolates of S. zooepidemicus can carry doxycycline resistance genes and that empirical use of antibiotics during outbreaks of disease might be driving the selection pressure for antibiotic-resistant strains. 21
Mycoplasma cynos
Etiology
Mycoplasma cynos is a member of the class Mollicutes. The Mollicutes are unique among bacteria owing to the lack of a cell wall. They are the smallest living organisms capable of independent existence and are thought to have evolved from the gram-positive Firmicutes (eg, Bacilli, Clostridia) taxon by considerable genome reduction. 90
M. cynos was first isolated from the lungs of a dog with clinical pneumonia in 1972. 94 Since this initial discovery there has been intermittent research interest in the organism; as such, there are limited data available regarding the prevalence, pathogenesis, and nature of the immune response associated with M. cynos infection. A comprehensive review of canine mycoplasmas was recently published by Chalker; 18 here we focus on M. cynos and highlight some of the more recent literature regarding its contribution to CIRD.
In 2004, a study was undertaken to investigate the microorganisms involved in CIRD at a large rehoming kennel. Mycoplasmas were isolated from both diseased and healthy dogs; however, M. cynos was the only mycoplasma significantly associated with respiratory disease. 20 More recently, M. cynos has been isolated from several cases of CIRD including lethal bronchopneumonia in a litter of Golden Retriever puppies; 113 pneumonia in a Miniature Pinscher caused by concurrent infection with M. cynos, canine distemper virus, and CAV-2; 23 and respiratory disease in a colony of laboratory Beagle dogs. 53
Clinical Disease
Determining the clinical signs associated with M. cynos infection is far from straightforward because this bacterium is often isolated from dogs that are concurrently infected with other pathogens, particularly viruses, which also contribute to clinical disease. 93 In agreement, Chalker et al 20 reported that in the main study population in their investigation, Bordetella bronchiseptica, CRCoV, S. equi subsp zooepidemicus, and M. cynos were all associated with CIRD and that further work was required to understand the interaction between these organisms and the contribution of M. cynos to respiratory disease.
Reports suggest that the pathological changes associated with isolation of M. cynos include grossly palpable lung consolidation, loss of cilia on bronchial and bronchiolar epithelial cells, low-grade serofibrinous pleuritis, severe acute generalized catarrhal-suppurative and hemorrhagic or fibrinonecrotizing bronchopneumonia with infiltration of lymphocytes and other mononuclear cells, pulmonary edema, exudation of neutrophils and macrophages into alveolar spaces, and hyperplasia and exfoliation of type II pneumocytes. 23,53,113
It is difficult to be sure whether M. cynos is able to cause these pathological changes alone or whether they are a result of concurrent infections; however, a recent case report of puppies with lethal pneumonia has provided evidence to suggest the former. 113 During this investigation, immunohistochemical examination of formalin-fixed, paraffin-embedded lung tissue using a polyclonal antibody to M. cynos revealed punctate antigenic signals surrounding foci of severe neutrophilic inflammation; no specific staining was shown in 6 age-matched negative control dogs. Furthermore, mycoplasmas were cultured solely and in abundance from samples of lung tissue and these were later confirmed to be M. cynos. This evidence suggests that M. cynos played a central role in the development of bronchopneumonia and pleuritis in these puppies. 113
Pathogenesis
Transmission of M. cynos usually occurs via droplets from nasal and ocular secretions and saliva. In 2004, Chalker et al 20 reported that during their investigation, M. cynos was the only species of mycoplasma to be isolated from the air of the kennel. Dogs were most likely to be infected with M. cynos during the first 2 to 3 weeks of their stay in the kennel, and a decline in the isolation of M. cynos was observed in dogs housed for periods greater than 21 days, possibly reflecting an active immune response to the bacterium. 20 Later, Rycroft et al 95 confirmed this association and detected an increase in antibody titer to M. cynos over a 21-day interval within the kennel; the authors reported that the response was positively related to clinical respiratory disease.
Very little research has been performed on molecular characterization of M. cynos. A recent report describes the strain typing of 14 M. cynos isolates from 6 dogs with respiratory disease: 68 pulsed-field gel electrophoresis and random-amplified polymorphic DNA were used to investigate the genetic similarity of the strains from 2 separate kennels in the UK. Isolates from each kennel were found to be genetically similar; furthermore, even after a break of several months, genetically identical isolates were recovered from within the same kennel, suggesting persistence of strains within the local environment. 68
The recent identification of sialidase activity in M. cynos 7,70 has provided some insight into how the bacteria may cause disease and spread within the canine host. Sialidase is an enzyme that assists in microbial colonization and dissemination within the host. 24 It is a known virulence factor in other bacteria, including some mycoplasma (M. alligatoris, M. synoviae, and M. gallisepticum). 8,12,70,72 The enzyme can exert a direct toxic effect on the host cells as well as interfere with host defense mechanisms. 24 The sialidase enzyme catalyzes hydrolysis of α-(2-3)-, α-(2-6)-, and α-(2-8)-glycosidic linkages of terminal sialic residues in oligosaccharides, glycoproteins, glycolipids, colominic acid, and synthetic substrates. It is thought that sialidase activity in some mycoplasmas is due to the utilization of sialic acid residues for attachment to host epithelial cell surfaces. 71
Due to the lack of an available genome sequence, genetic characterization of M. cynos has been limited in comparison to other more frequently studied mycoplasma species. The recent publication of the M. cynos genome sequence 108 will help to improve understanding of M. cynos pathogenesis. Comparative genomic analysis between M. cynos and other well-studied mycoplasmas will aid identification of putative virulence determinants that could inform future therapeutics to prevent or control M. cynos infection in dogs. Indeed thus far, there is no conclusive evidence to confirm whether M. cynos is a primary or secondary pathogen, and further work is necessary to determine the pathogenesis of mycoplasma infection in dogs.
Diagnosis
M. cynos is a fastidious organism requiring complex growth media when cultured in vitro. As a consequence, mycoplasma infection in dogs is often overlooked since few laboratories routinely test for its presence unless requested by the veterinarian. 18 Underdiagnosis of mycoplasma infections of dogs with respiratory disease may have contributed to the lack of research into M. cynos over the years. When mycoplasma culture is requested, diagnosis is typically achieved by culture of the organisms from nasal or oropharyngeal swabs followed by genus-specific PCR and M. cynos–specific PCR to confirm identity. 20
Summary
Canine infectious respiratory disease is arguably one the most frequent clinical syndromes for which group-housed dogs are submitted for both veterinary examination and treatment. Prevention is key to its control, and, certainly for viral pathogens, successful vaccination is essential. Current vaccines, although having their place in preventive strategies for CIRD, are not without their limitations as was clearly highlighted with CRCoV. Vaccines can only be as good as the antigens they contain, and if essential ones are absent, their protection will be limited. However, taking a more holistic approach, prevention of viral-induced disease could result in lower antibiotic use, the indiscriminate use of which may have contributed to the emergence of resistant strains of S. zooepidemicus.
Of the viral agents discussed, all have the potential to be a primary agent of disease; however, within the disease complex it is likely that they work together to predispose dogs to secondary bacterial infections, leading to a prolonged or more severe clinical disease, similar to that observed with other complex respiratory infections in cattle and swine.
CIRD is undoubtedly a complex infection, and future vaccines and control strategies will need to reflect this multifactorial etiology. This review presents a contemporary evaluation of 8 pathogens that may, in various dogs, contribute to the overall pathogenesis of CIRD. In time, research and development will need to unravel their importance and their interactions with each other and also to define future control strategies.
Preventing and controlling respiratory infections in dogs are of concern to veterinarians and pet owners, but increasingly the close interrelationship between dogs and humans should reinforce the need to investigate canine respiratory outbreaks comprehensively. Indeed the current “One Health” agenda, based on the idea that animal health and human health are inextricably linked, has no greater relevance than when considering CIRD since many of these shelter/rescue dogs go on to become household pets potentially harboring these organisms.
Canine infectious respiratory disease is an ever-changing syndrome—viruses and bacteria continue to evolve and adapt with research always lagging behind, at times taking on a retrospective approach. As techniques for identifying novel pathogens continue to advance, new agents are being discovered at an increasingly rapid rate. The future challenge for veterinarians and in particular veterinary pathologists is to never become complacent and to always be alert to the potential for new or reemerging infections.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.