
Abstract
Aging is the biological process of declining physiologic function associated with increasing mortality rate during advancing age. Humans and higher nonhuman primates exhibit unusually longer average life spans as compared with mammals of similar body mass. Furthermore, the population of humans worldwide is growing older as a result of improvements in public health, social services, and health care systems. Comparative studies among a wide range of organisms that include nonhuman primates contribute greatly to our understanding about the basic mechanisms of aging. Based on their genetic and physiologic relatedness to humans, nonhuman primates are especially important for better understanding processes of aging unique to primates, as well as for testing intervention strategies to improve healthy aging and to treat diseases and disabilities in older people. Rhesus and cynomolgus macaques are the predominant monkeys used in studies on aging, but research with lower nonhuman primate species is increasing. One of the priority topics of research about aging in nonhuman primates involves neurologic changes associated with cognitive decline and neurodegenerative diseases. Additional areas of research include osteoporosis, reproductive decline, caloric restriction, and their mimetics, as well as immune senescence and chronic inflammation that affect vaccine efficacy and resistance to infections and cancer. The purpose of this review is to highlight the findings from nonhuman primate research that contribute to our understanding about aging and health span in humans.
Keywords
Nonhuman primates share similar physiology and a close phylogenetic relationship to humans. The use of nonhuman primates in comparative experimental studies thus contributes to our knowledge about aging processes and translation of applications for improving health span in humans and other animals. Key evolutionary and adaptive features in nonhuman primates include pentadactylism for grasping and manipulating objects, binocular vision enabling 3-dimensional depth perception that is related to development of the cerebral cortex, and progression toward bipedalism and upright posture. 124,224 Correlates of average natural life span of an organism are highly complex, but body size in conjunction with metabolism, reproduction, immunity, and environmental stress, among other factors, is associated with average longevity such that larger animal species tend to live longer. 14,63 Interestingly, human and nonhuman primates exhibit unusually longer average life spans that are nearly 4-fold higher than those of most other mammals relative to their body sizes. 14 In addition, nonhuman primates exhibit similar key life span metrics as humans, such as higher infant mortality rate, followed by lower mortality during the juvenile stage and then an extended period of increasing age-related morbidity and mortality. 32 Since nonhuman primates can be studied under well-controlled experimental conditions, intervention strategies for evaluating vaccines or drugs are often tested in nonhuman primates for safety and efficacy before application in humans. This review highlights the contributions of nonhuman primate research to our understanding about aging and health span in humans.
By far, the predominant nonhuman primate species utilized in biomedical research facilities as well as for studies on aging are rhesus macaques (Macaca mulatta) and cynomolgus macaques (Macaca fasicularis). 124,128 Specifically, among the facilities with nonhuman primates in North America that were recently surveyed, 80% housed rhesus macaques of Indian or Chinese origin, followed closely by cynomolgus macaques housed in 73% of the facilities. 128 Aspects of aging research studies that utilize macaques include neurobiology, anatomy, physiology, cognition, and behavior, 18,163,184,212,216 as well as reproductive senescence, 11 caloric restriction (CR), 46,143 and immune senescence. 10,148,213 The use of macaques in research appears to represent the best compromise between phylogenetic and physiologic relatedness to humans, cost efficiency, life span, resources, expertise in animal husbandry practices, and adaptability for translation of results to humans. To improve efficiency, accessibility, and applicability, however, increasing emphasis is being placed on purpose-bred animals and further advancing animal husbandry practices so that lower primates also may be included for relevant model development of research on aging.*
Prosimians, or “premonkeys,” are the most phylogenetically distant nonhuman primates from humans. Among the prosimians, grey mouse lemurs (Microcebus murinus) have been the most extensively studied for relating processes of aging in relation to humans. 16,126 For example, the mouse lemur was the first nonhuman primate species to demonstrate a relationship between cerebral atrophy and cognitive decline with aging that simulated what was seen in aging humans. 167 Neuroscience studies about memory, behavior, and psychomotor function have utilized both captive and wild mouse lemurs. † Results from studies using mouse lemurs also demonstrated correlations between aging and renal function, 5,142 sleep deprivation, 174 circadian rhythms, 84 reproduction, 33,56 thermoregulation, 202 –205 metabolism and dietary restriction, 57,58,125,140,141,168 oxidative stress, 53 and replicative senescence. 197 The use of prosimians in research is more cost-efficient, but limitations include their smaller size that restricts specimen sampling; differences in metabolic, biochemical, and endocrine responses compared with humans; and a need for continued development in animal husbandry techniques to reduce stress-related behaviors of captive prosimians. 16
Among the New World monkeys (NWMs), common marmosets (Callithrix jacchus) often are used to examine aging related to neurogenesis, vision, neuroanatomy, cognition, and pathogenesis of neurodegenerative diseases such as Parkinson and Alzheimer. ‡ Research on reproductive senescence 132,179 and metabolic changes associated with aging (eg, insulin-resistance, diabetes, and obesity) also utilize common marmosets. 159,178,200 Cotton-top tamarins (Saquinus oedipus) and squirrel monkeys (Saimiri sciureus) are used to examine reproductive senescence and in comparative studies of aging. §
Chimpanzees (Pan troglodytes) have been the most common great apes used in biomedical research, as they closely resemble humans structurally and genetically, with >98% DNA sequence homology. 49 Chimpanzee use in research, however, has declined due to ethical concerns, high costs, and complexities in maintenance. 75 –77
Psychosocial Influences
Quality of life during aging is dependent on complex interrelationships between psychosocial and physical health parameters. Studies of humans indicate that psychosocial dimensions comprise networks of social relationships, feelings of connectedness versus loneliness, and overall satisfaction with life that together compose predictors of longevity and quality of life. 2 Measurements of such parameters often are subjective and utilize self-evaluations among humans of varying ages and socioeconomic strata. Analogous research about psychosocial influences and aging in nonhuman primates examines social rankings and stress in captive and free-ranging colonies. Rhesus macaques in free-ranging colonies exhibit a median life span of approximately 15 years of age due to predation, a lower ability of older animals to find food, reproductive stress with aging, and increased accumulation of “wear and tear” effects, also referred to as allostatic stress. 101 Captive rhesus macaques, however, have an average median life span of 25 years. Comparison between the 2 housing conditions thus enables evaluation of environmental influences on restorative homeostatic mechanisms during aging. 43,45,101
Social rank among aging females constitutes an important measure for relating psychosocial stress to aging that may be comparable to socioeconomic pressures and effects on quality of life in aging humans. 2,101 Female social dominance ranking is inherited, and female rhesus macaques born to low-ranking females tend to remain at lower social status. Characteristics of the higher psychosocial stress associated with lower social status and aging have linked the hypothalamic-pituitary-adrenal axis, immune senescence, and reproductive stress with decreased longevity, allostatic (“wear and tear”) stress, and reduced quality of life. 85,101 Measureable indicators of psychosocial stress and aging featured higher levels of fecal cortisol, increased proinflammatory serum cytokine levels, declining ex vivo natural killer cytotoxic activity (as a function of immune senescence), and declining body mass index and body weight. In addition, older lower-ranking female rhesus macaques exhibited reproductive stress and produced smaller infants at wider time intervals to reduce energy output than did higher-ranking older females. 101 Reduced psychosocial stress during aging, however, was reported in rhesus macaques and Japanese macaques living in colonies where strong intergenerational relationships developed among females to presumably provide mutual support in conflicts with nonfamilial members. 2,45,147
Caloric Restriction
Over 75 years ago, McCay et al reported that calorie intake reduction by approximately 30% of an ad libitum diet that avoided malnutrition increased longevity in rats. 145 CR has since been reported to also extend average life span in many organisms, including yeast, worms, fruit flies, and mice. 46,117 Beneficial effects in rhesus macaques suggest that CR also may contribute to improving human health span. 46,117 Many of the basic mechanisms of longevity associated with CR that affect cell growth, metabolism, and reproduction are conserved across species, but physiologic outcomes and biomarkers of improving health span appear to be more complex in higher mammals and primates. 148,229 In addition, CR may not be feasible in humans, because compliance and milestones would be difficult to monitor and achieve. For example, long-term CR in humans is associated with reduced libido and lower body temperature. 193 Therefore, nonhuman primates are being used, first, to define the beneficial effects of CR that promote healthier aging and, second, to define and test pharmacologic CR mimetics for safety and efficacy that may then be applied to humans for improving health span and life span. 46,103,105,117,131,141
Old World monkeys (OWMs), such as rhesus macaques, cynomolgus macaques, and baboons, play prominent roles in CR studies because they exhibit higher risks for obesity, insulin resistance, type 2 diabetes, immunologic decline, and metabolic syndrome related to cardiovascular disease (CVD) and impaired cognition, which also occur in aging humans. 27,46,52,117,206,226 CR initiated in nonhuman primates after reaching maturity improved health span and was associated with increased insulin sensitivity, decreased hyperglycemia and incidence of type 2 diabetes, preservation of skeletal muscle function, reduced visceral white adipose fat, attenuated development of CVD, and reduced total energy expenditure. If initiated prior to adulthood, however, CR was deleterious to immune responses in rhesus macaques 150 and, when administered to pregnant baboons, induced increasing insulin resistance and higher risk for developing type 2 diabetes in the offspring. 40 The magnitude of increased life span as a consequence of CR, however, was reported to decline with increasing complexity of the organism. 148,229 This contributes to a debate about whether rhesus macaques undergoing CR exhibit increased life span because of the varied or polymorphic genetic backgrounds in the test populations, different levels of physical activity, and/or medical interventions applied to monkeys that may not have been used in studies with lower animals. 46,104,117,143,148
Antiaging compounds are being tested for efficacy in nonhuman primates, especially in rhesus macaques fed a high-fat/high-sucrose diet that induces many of the morbidities associated with aging, such as obesity, type 2 diabetes, and CVD. Sirtuins are NAD+-dependent deacetylases that participate in DNA repair, mitochondrial biogenesis, and regulation of inflammation. 131,154 Treatment of animals subjected to high-fat/high-sucrose diet with the sirtuin activator resveratrol prevented central arterial stiffening, increased insulin sensitivity, reduced blood glucose levels, preserved β cells in islets of Langerhans, reduced adipose cell size, and reduced macrophage infiltration with declining NFκB expression (ie, lowered inflammation). 78,109,144 Mouse lemurs, photoperiod-dependent prosimian primates, also have been examined for effects of CR or resveratrol treatment as a CR mimetic. These animals acclimated well to their treatments after 1 year, and while oxidative stress–producing DNA and RNA damage increased initially, animals in the CR treatment group and in the resveratrol-supplemented diet subsequently exhibited reduced DNA and RNA damage during aging. 58,141 The use of nonhuman primates is thus expected to continue for preclinical safety and efficacy testing of antiaging compounds.
Organ Systems
Immune System and Inflammation
Immune senescence is a decline in regulated immune responses and increasing susceptibility to infections as well as noncommunicable diseases. Chronic inflammation is one of the hallmarks of immune senescence associated with most diseases of aging and is commonly referred to as “inflamm-aging.” 79,80 Advantages to using nonhuman primates to study immune senescence include similarities to humans in immune responses, cross-reactivity in immunologic reagents (eg, antibodies), and comparable range of infectious and noncommunicable diseases of aging, such as adenocarcinoma of the colon and chronic inflammation observed in rhesus macaques (Suppl. Figs. 1, 2). Also, monkeys can be naturally or experimentally infected with many of the same pathogens that infect humans to test efficacy of vaccines and treatments under controlled experimental conditions. Furthermore, nonhuman primates are amenable for use in longitudinal studies that require repeated sampling, and such studies can be better controlled for time of infection, compliance, and risk behaviors (eg, illicit drug use, cigarette smoking) that may confound interpretation of results in human studies.
Several reviews describing immune senescence in rhesus macaques have been published recently. 91,149,152 Adaptive immune responses appear to be more affected with age than innate immune responses. 91 A recent report comparing young, adult, and aged rhesus macaques, however, identified subtle increases in CD14+CD16+ monocytes (toward a nonclassical monocyte/macrophage phenotype) and a shift toward the CD11c+ myeloid dendritic cell population. 10 Antigen-presenting cell populations and Toll-like receptor expression are broadly similar between adult and aged populations. A decline in ex vivo cytotoxic function in natural killer cells strongly correlates with aging and is predictive for impending mortality in aged rhesus macaques. 44 Changing levels of circulating proinflammatory cytokines, such as increases in TNF-α, IL-6, and IFN-γ, also correlate with age in humans and nonhuman primates, especially rhesus macaques. It is still difficult, however, to specifically define the effects of shifting levels of cytokines, chemokines, and other circulating factors on biological versus chronological aging outcomes among individuals (ie, as measures of healthy vs less healthy aging). 10,66,152
The most discernible changes during aging and in the adaptive immune system are observed in the T-lymphocyte compartment. Absolute numbers of circulating T cells do not change, but aging is associated with declining naïve T cells, increasing numbers of terminally differentiated effector memory T cells, and declining CD4+-to-CD8+ T cell ratios. 66,91,106 The shifts in these cell populations are associated with decreasing progenitor stem cells in bone marrow and thymic atrophy, as also observed in nonhuman primates (Suppl. Figs. 3, 4). In addition, continued exposure to pathogens and persistent infections promote a shift toward increased effector memory T cells and a shrinking naïve T-cell antigen recognition repertoire. 41
The repertoire and number of circulating B cells decline with aging in humans and rhesus macaques. 66,91 Homeostatic responses or shifts in subpopulations of B cells, however, have not been fully examined in aging nonhuman primates. Mucosal antibody responses become compromised with increasing age. IgA responses to cholera toxin/cholera toxoid were lower in intestinal lavage specimens of older rhesus macaques, while IgM levels were higher. 201 This was explained by the observation that emigration of IgA immunoblasts from Peyer patches to the small intestinal lamina propria declined in aging rhesus macaques due to reduced expression of homing molecules. 183
Vaccine studies in older rhesus macaques further demonstrate declining immune competence with aging. Antibody responses in influenza-vaccinated rhesus macaques >19 years of age were significantly lower than in young adults aged 4 and 7 years, but boost immunizations in the older animals produced responses comparable to those of younger monkeys. 42 Preexisting antibodies from earlier seasonal exposures are believed to explain why older humans were more resistant to the 2009 H1N1 influenza virus than younger individuals. Since humans could not be used to test this, naïve older adult rhesus macaques were infected with the CA04 H1N1 influenza virus. These animals developed higher viral loads than infected younger adults and also produced delayed T-cell responses, thereby supporting the hypothesis that preexisting antibodies in the older humans may be protective for some seasonal influenza viruses. 112
Intervention strategies to improve immune responses during aging or to delay immune senescence are being tested in rhesus macaques as well. Older rhesus macaques vaccinated for influenza and treated with IL-7, a cytokine that promotes T-cell homeostasis, exhibited increased production of naïve T cells and higher antibody responses than older monkeys not given IL-7. 9 CR similarly improves immune functions and delayed T-cell senescence in rhesus macaques when initiated during young adulthood (ie, approximately 5–7 years of age). 143,151 Conversely, HIV-infected individuals undergoing antiretroviral therapy develop chronic diseases associated with aging earlier than non-HIV-infected persons and are considered to undergo accelerated aging. 1,50,100 The rhesus macaque model of simian immunodeficiency virus (SIV) infection and antiretroviral therapy is emerging as a model to study immune response mechanisms of accelerated aging, as there are no reported genetic mutations in nonhuman primates analogous to Hutchinson-Gilford Progeria and Werner progeria syndromes that occur in humans. 160
In summary, immune senescence and diseases of aging in humans and nonhuman primates are characterized by chronic inflammation and greater susceptibility to infectious diseases and malignancies. Responses to vaccines in the elderly are also reduced and less effective. These changes are generally associated with increasing levels of circulating proinflammatory cytokines and chemokines, a reduced naïve T-cell antigen recognition repertoire, and a growing population of effector memory T cells against persistent infections. It is anticipated that with aging of the human population worldwide and the growing pressures on public and medical health systems, the use of nonhuman primates will continue to expand for developing prevention and treatment strategies.
Central Nervous System
The effect of aging on the central nervous system is an area of intense interest in humans and thus contributes to the growth of studies examining similarities with nonhuman primates. As in humans, there is evidence of increased oxidative stress in aged rhesus macaques, 210 which is linked to higher circulating levels of homocysteine and cognitive decline. 223 CR, conversely, appears to contribute to successful cognitive and neurologic aging in macaques through maintaining white matter volume, 21 lower levels of neuroinflammation, 221 cortisol, 222 and iron, 116 as well as decreased mortality. 28
Published studies describing lesions and changes in the central nervous system during aging most commonly use species of nonhuman primates phylogenetically closer to humans (Table 1). As nonhuman primates age, some neuronal populations are more affected than others, such as the shifts in density of calcium binding protein expression on neurons. 88,89 For example, the percentage of calbindin-positive inhibitory GABAergic (gamma-aminobutyric acid) neurons were reported to increase over age in rhesus macaques, while there were fewer parvalbumin-positive glutamic acid decarboxylase 67 neurons. The investigators speculated that this shift in binding receptor density on neurons is important for buffering calcium and regulating excitotoxic effects. Reduced cognitive capacity in aged macaques was also associated with breakdown of myelin sheaths 164 and axon loss. 181 Furthermore, a reduced organization in neuronal arrangement was observed in aged animals 97,221 that was related to loss of white matter volume in key brain areas, including frontal lobe and cerebellum, 139 possibly contributing to an anterior-to-posterior gradient decline in white matter integrity 195 and loss of synapses and synaptophysin expression. 92 Overall brain volume declines in all primate species during aging but occurs proportionally later in chimpanzees than in humans. 39 Aged humans, however, have a higher prevalence of neurodegenerative diseases causing dementia, including Alzheimer and Parkinson diseases. One explanation is that the longer life span in humans compared with nonhuman primates provides time for greater deterioration of white matter prior to death. 39
Frequency of Studies of Age-Related Brain Disease Characteristics in Nonhuman Primates and Humans.a

aSearch terms indicated on the table were entered into PubMed (http://www.ncbi.nlm.nih.gov/pubmed), and the number of citations between 1964 through 2014 are indicated as follows: +/–, <10 citations; +, 10–99 citations; ++, 100–999 citations; +++, 1000–4999 citations; ++++, 5000 – 9999 citations; +++++, ≥10 000 citations.
Alzheimer Disease
While all primates exhibit a degree of progressive age-related neurodegeneration, the drastic neuron loss and clinically observed cognitive decline of Alzheimer disease are considered unique to humans. 77 Indeed, the great apes demonstrate fewer changes associated with brain aging than humans. 77 Several nonhuman primate models, however, are being used to explore aspects of neurodegeneration that occur in Alzheimer disease. 76,77,82,99,216
As noted above, diet is considered a major risk factor for metabolic syndrome and diabetes, which are associated with Alzheimer disease. 121 Diet and obesity are linked to neuroinflammation (astrogliosis) in rhesus macaques but not to the presence of amyloid in the brain. 196 However, in cynomolgus macaques, reactive astrocytes are reported to engulf β-amyloid, 119 an important step in the initiation of amyloid plaque formation. This occurs in conjunction with lower Aβ42 in the cerebral spinal fluid and accumulation in the brain. 227 Amyloid β deposits also are observed in brain of aged marmosets. 82 Curiously, there are neither degenerative nor histologic changes, even in areas with plaques that were apparent by magnetic resonance imaging. 211
Phosphorylation of tau protein is evident on glutamatergic synapses in aged rhesus macaques, 34 and tau accumulation in cerebral spinal fluid is linked to poor memory in older cynomolgus macaques. 60 Environmental influences or exposures during development may contribute to the increased phosphorylation and accumulation of tau protein that are associated with neurodegenerative disease later in life. For example, 23 years after being exposed to lead as infants, a group of rhesus macaques had greater levels of tau and β-amyloid in their brains than aged macaques that were not exposed to lead as infants. 24 Neuropathology similar to that observed in human Alzheimer disease, such as tau hyperphosphorylation and extensive neurofibrillary degeneration, is seen in the mouse lemur as well. 184 However, β-amyloid is rarely observed in nonhuman primate brains during aging, 77,209 thus limiting the use of nonhuman primates in Alzheimer disease research. In this respect, the nonhuman primate brain may be a better model for normal brain aging as opposed to age-associated brain disease.
Parkinson Disease
Parkinson and Parkinson-like diseases are related to a loss of dopamine within synaptic vesicles 169 or loss of these synapses. 134 In rhesus and cynomolgus macaques, administration of 1-methyl-4-phenyl-1,2,3,6 tetrahydropyridine (MPTP) is used to induce signs of Parkinson-like disease. MPTP administration activates microglia in rhesus and cynomolgus macaques 19,114 and decreases glutamate levels in rhesus macaques. 70 Rhesus and cynomolgus macaques also respond differently during the acute phase of MPTP administration. Cynomolgus macaques are generally more severely affected than rhesus macaques by measures of dyskinesia, parkinsonian motor disability scores, and functional recovery following withdrawal of MPTP. 171 In addition, both species require multiple injections of MPTP to induce stable Parkinson-like disease, but the doses required vary among individual monkeys of both species.
Similar to humans, treatment with L-DOPA significantly improves the clinical neurologic function of cynomolgus macaques. 87,156 In rhesus macaques, however, L-DOPA treatment produced limited recovery since hypokinesia and tremor were responsive to L-DOPA, but gait disorders did not improve. 87 The comparatively better outcome in cynomolgus macaques may result from using antagonists to glutamatergic receptors in addition to L-DOPA to prevent dysregulation of associated signaling proteins, such as pERK, D2 receptors, and protein kinase B. 156
The macaque models of MPTP-induced Parkinson-like disease also explored gene therapy approaches to correct for the loss of dopaminergic neurons. 230 These studies used adenoviral vectors to transduce rhesus adipose mesenchymal stem cells to stably express neurturin and tyrosine hydroxylase. Once these stem cells differentiated into dopaminergic neurons and implanted into striatum and substantia nigra, there were significant improvements in the behavioral signs (eg, balance disorder scores) of MPTP toxicity in rhesus macaques. In addition to the MPTP toxicity model, a second model of Parkinson disease in marmosets used a recombinant adeno-associated virus vector to overexpress α-synuclein in dopaminergic neurons within the substantia nigra pars compacta. This approach was successful in inducing the loss of dopaminergic neurons and expression of Parkinson-like signs. 74
Respiratory System
The lung undergoes changes in structure and function during aging that contribute to increased chronic lower respiratory tract diseases (eg, emphysema, bronchitis, chronic obstructive pulmonary disease [COPD]), which compose the third-leading cause of death in humans. 137,186 Alterations in the ribs and spine, as well as declines in muscle strength and efficiency, contribute to decreased thoracic cavity volume as well as reduced maximum lung inhalation and expiration volumes. Furthermore, the oxidative stress from inflammatory responses to infectious agents and pollutants produces damage to lung tissue architecture and adversely affects efficiency of gas exchange. Changes observed in lung tissue of aged rhesus macaques, for example, demonstrated pigment accumulation in macrophages as well as atheromatous arterial thickening (Suppl. Figs. 5, 6).
COPD dramatically increases in prevalence in elderly humans and is associated with greater susceptibility to respiratory infections. 186 Nonhuman primate models are used to better understand risk factors and mechanisms of COPD. For example, cigarette smoking is considered a principal risk factor in genetically susceptible individuals and exposure of cynomolgus macaques to cigarette smoke produced pulmonary disease similar to that observed in humans (eg, airway inflammation, peribronchial fibrosis, mucus metaplasia, bronchial lymphoid aggregates). 170 HIV-infected individuals with Pneumocystis jiroveci (syn carinii) pneumonia develop COPD, and this is being modeled in rhesus macaques with SIV and Pneumocystis coinfection. 120,190
Cardiovascular System
CVDs comprising coronary heart disease, heart failure, peripheral vascular disease, and stroke are the most common causes of death in older persons living in the developed world. Normal changes associated with aging include increased vascular rigidity or stiffening and reduced compliance or elasticity. 129,158 These changes are exacerbated by comorbidities and risk factors, such as diabetes, hypertension, dyslipidemia, sex, smoking, and obesity.
Arterial remodeling was observed with advancing age in rhesus macaques and was characterized by an increased intimal thickness, higher matrix metalloproteinase 2 mRNA and protein expression, and enhanced angiotensin II signaling. 220 Males tend to be at higher risk for CVD. In cynomolgus macaques, for example, aortic stiffness was more pronounced in males than in females, even though aortic pulse pressure similarly increased with aging in both males and females. 173 Microarray analyses of the aorta indicated that, compared with females, aged males exhibited greater expression of genes associated with smooth muscle switching from a contractile to a secretory phenotype. 173 Furthermore, while collagen density was maintained in both sexes of older cynomolgus monkeys, males displayed significant decreases in elastin density along with decreasing collagen type III and increasing collagen type VIII. 172 In contrast, no sex differences in collagen isoform shifts were observed in aging rats, thereby promoting the use of nonhuman primates to examine sex-specific differences in aortic stiffness with aging.
Ex vivo studies further demonstrated changes associated with aging in endothelial cells derived from femoral arteries of nonhuman primates. Endothelial cells from older baboons exhibited a more rapid decline in endothelium-derived nitric oxide synthase levels ex vivo as a biomarker for vascular aging. 188 Endothelial cells harvested from baboons fed a high-fat/high-sucrose diet demonstrated replicative senescence and increased expression of senescence-associated β-galactosidase independent of telomere length, and this model is being used further to study mechanisms of premature vascular senescence. 189 Fewer endothelial colony-forming cells could be harvested from blood of aged versus younger rhesus macaques, and these cells were less able to form vessels after transplantation into immunodeficient mice. 187 Ex vivo studies using vascular smooth muscle cells from rhesus macaques demonstrated that with increasing age, the nuclear factor erythroid–derived 2 pathway that regulates antioxidant responses was blunted or became dysfunctional, thereby inducing an increase in oxidative stress leading to NFκB activation and vascular inflammation. 210 Studies in baboons indicated that clinical biomarkers of CVD were affected by genes that regulate lipid and lipoprotein metabolism. 51 Heritable traits affecting risk for CVD unique to primates were examined in baboons and reported to be associated with obesity and red blood cell sodium-lithium countertransport activity, an indicator for salt-sensitive hypertension. 51
Intervention strategies to inhibit the rate of vascular senescence are being tested in nonhuman primates. Human endothelial cells incubated ex vivo with serum from rhesus macaques undergoing CR exhibited increased angiogenesis and greater expression of vascular endothelial growth factors than cells incubated with serum from macaques fed ad libitum. 54 Primary vascular smooth muscle cells from older rhesus macaques secreted higher levels of proinflammatory cytokines ex vivo (eg, IL-1β, MCP-1, TNF-α, IL-6) than did cells from younger animals, and this secretory profile was reversed by exposure to resveratrol, an anti-inflammatory and antiaging polyphenol. 55 Resveratrol also inhibited the rate of arterial wall inflammation and stiffening in rhesus macaques fed a high-fat/high-sucrose diet. 144
Nonhuman primates are being used to test hormonal intervention strategies related to risk for CVD. Estrogen deficiency, for example, is known to accelerate atherosclerosis during and after menopause. Estrogen replacement helps reduce development of coronary artery disease, but concerns exist about endometrial hyperplasia in the absence of combination hormone therapy with progesterone. As a result, studies in humans are being hampered by decreasing compliance due to a fear of breast cancer. 153 In rhesus and cynomolgus macaques, however, an orally active progestin, nomegestrol acetate, was reported to counteract the endometrial stimulation of estrogen, and this could prove beneficial for continued development of hormone replacement therapy to ameliorate development of atherosclerosis in postmenopausal women. 161 In addition, female cynomolgus monkeys fed a diet rich in soy protein and isoflavones exhibited a lower risk for CVD than monkeys fed a casein-lactalbumin diet. 6
Musculoskeletal System
The overall degeneration of bones, joints, and muscles during aging contributes to frailty increasingly observed with advancing age. Bones undergo thinning through a loss of minerals (eg, calcium) and a shift from bone production by osteoblasts toward resorption by osteoclasts leading to osteoporosis. The accumulated wear and tear on cartilage and joints leads to calcification and mineral deposition producing less flexibility. Osteoporosis commonly progresses during aging via systemic loss of bone mass and deterioration of trabecular architecture that increases the risk for bone fractures. 180 Skeletal muscle undergoes atrophy during aging from reduced production and loss of muscle fibers, reduced size of fibers, accumulation of mitochondrial DNA mutations, and deficiencies in cytochrome oxidase production that either alone or in combination affect strength and energy metabolism of the muscle. 72 Chronic inflammation associated with aging contributes to the overall degeneration of the musculoskeletal system and further promotes osteoarthritis that is considered the most common cause of chronic disability in the elderly. 135
Nonhuman primates, especially OWMs (eg, macaques, baboons), are particularly useful for studying changes in the musculoskeletal system because they develop bone and muscle loss similar to humans during aging. 37,64,65,175 OWMs and nonhuman hominids (eg, great apes) display haversian osteonal remodeling of cortical bone that occurs in humans but not in rodents, and they have a similar reproductive endocrine system that affects bone metabolism. 26,51,95,108
Increasing parity of up to 7 births by free-ranging rhesus macaque females is beneficial to retain bone mineral density and partially offsets the effects of bone loss from aging, while lower numbers of births are associated with earlier osteoporosis. 35 While less frequently studied, baboons also display similar skeletal biology as humans with respect to bone composition and causes of fracture. 51,96 Approximately 25% of older female baboons exhibit osteopenia and a steady decline in bone mineral density beginning at about 17 years of age. Unlike rhesus macaques, however, parity or interbirth intervals do not distinguish baboons with low versus higher bone marrow densities. 96
Estrogen decline after menopause accelerates bone loss and decreasing bone mineral density in OWMs. 20 In addition, ovariectomy in macaques reportedly mimics early and premenopausal changes that also occur in humans, such as remodeling and bone loss. 31,108 The ovariectomy model has been especially important for evaluating effects of estrogen replacement therapy, bisphosphonates, odanacatib (a cathepsin K inhibitor), intraosseous recombinant human bone morphogenetic protein 2 / calcium phosphate matrix, and parathyroid hormone on progression of osteoporosis, osteopenia, and osteoarthritis. Results from several of these studies were consistent with those from human clinical trials. ||
Castration (orchidectomy) has been applied as a model for osteopenia/osteoporosis in male macaques and produced accelerated thinning of the skull, higher risk for vertebral fractures, kyphosis of the spine, and remodeling of vertebrae and femurs. 118 In male rhesus macaques, vitamin K deficiency—considered a risk for bone loss in conjunction with the long-term use of anticoagulants—did not adversely affect skeletal changes if the animals were also administered sufficient vitamin D and calcium. 25 Among NWMs, older marmoset females treated with alendronate, a bisphosphonate used to reduce bone loss, demonstrated improved trabecular volume and number, supporting their use to study human bone physiology during aging. 17
Nonhuman primates are being used to address treatments for degenerative joint disease characterized by osteoarthritis and synovial inflammation (Suppl. Fig. 7). Acupuncture 138 or treatment with glucosamine, chrondroitin, and polysulfated glycosaminoglycan along with corticosteroids and analgesics 215 was reported to improve mobility in captive chimpanzees. Surgical removal of the femoral head and neck in a rhesus macaque with naturally occurring osteoarthritis also improved clinical outcome. 67
Advancing age furthermore produces sarcopenia and reduces overall function in humans and nonhuman primates. 48 Multiple mitochondrial DNA mutations increase in skeletal muscle cells of aging rhesus macaques similar to that in humans. 130 In aging African green monkeys, there are changes in the myosin heavy chain isoforms, decreasing cross-sectional area of type I and type II fibers, and atrophy of type IIA fibers in the vastus lateralis that are also observed in aging humans. 73
Reproductive System
Studies in captive and free-ranging nonhuman primates have examined reproductive senescence and postreproductive life span in relation to the time of interbirth intervals, declining rates of sexual activity, lack of observed breeding behaviors, and changes in social relationships and hormone levels in various species of nonhuman primates. 11,13 The results suggest that there exists a regression in age-related reproductive decline whereby prosimians (eg, mouse lemurs) and some NWMs (tamarins) exhibit an abrupt reproductive decline near the end of life, 199,225 while OWMs (eg, macaques, colobines, mangabeys) and some of the apes (eg, orangutans) display gradual but heterogeneous rates of reproductive senescence analogous to perimenopause, 30,102,110,192 and chimps and gorillas undergo operational if not actual menopause. 12,214 Histologically, ovaries of an aged captive sooty manabey, for example, exhibited a loss in distinct primordial and secondary follicles (Suppl. Fig. 8). Differences in reproductive decline may exist between wild and captive colonies of nonhuman primates. For example, orangutans in the wild continue to reproduce throughout life but in captivity display a postreproductive life span, despite wild and captive orangutans sharing similar maximum life spans. 192 These findings suggest that animal management practices may affect reproduction patterns. Female NWMs such as marmosets and tamarins in wild and captive colonies do not exhibit gradual declines in fertility with aging, but infant mortality increases with the age of the mother, suggestive of declining maternal fitness with increasing age. 179,199,225
Comparative analyses imply an evolutionary continuum. Extended postreproductive life span was not evident in the prosimians and callitrichid primates but developed in the cercopithecine or OWMs and some apes, followed by the occurrence of menopause in gorillas and humans that exhibit a long postreproductive life span. 11 Thus, menopause may have evolved to reduce reproduction as a means to conserve energy consumption and thus sustain female survival for care of offspring. Interestingly, human postreproductive life span evolved from increased longevity and reduced death rates without an increased reproductive life span. This distinction has been further studied in wild colonies of a prosimian species, 2 NWMs species, 2 OWM species and 2 great apes, in relation to a hunter-forager Dobe ! Kung population that experiences natural fertility and mortality resembling our preagricultural ancestors. 3 Comparison of results in the 90th percentile of age at last live birth versus age at death showed that humans were unique and behaved as statistically significant outliers by exhibiting longer postreproductive life spans than nonhuman primates, which did fall along the correlative continuum. These findings thus argue in favor of an evolutionary role of human mothers and survival of their offspring via rationing of energy expense to support child-rearing over reproduction.
Nonhuman primate models of reproductive aging relate rates of pre-, peri-, and postmenopausal events to risks for developing chronic diseases of aging, such as coronary heart disease, osteoporosis, and cognitive decline. 7,191 Disruptions in ovarian function are attributed to stress, anxiety, and depression in women and nonhuman primates, providing models to test effects of environment on health benefits associated with reproductive aging and risk for chronic diseases. 115 Far less is known about reproductive senescence in males using nonhuman primate models, although declines in reproductive capacity were reported in male spider monkeys and rhesus macaques. 98,182 Results from one study on male rhesus macaques suggested that reproductive decline was highly variable and related to social and demographic factors rather than aging body condition. 22 In another study, supplemental administration of the androgens dehydroepiandrosterone and 5α-dihydrotestosterone to aged rhesus macaque males resulted in hormonal levels and circadian patterns similar to those in younger males, thereby supporting development of hormone therapies for treating elderly men. 194 While aspects of reproductive senescence are found to be uniquely human, nonhuman primates continue to be relevant for testing intervention and treatment strategies that affect the postreproductive quality of life of humans. 20
Genetics and Epigenetics
Aging appears to be somewhat delayed in humans, and comparative genetics studies that focused on humans and chimpanzees in relation to other organisms were performed to explore an evolutionary basis. 62 The DNA genome sequences of humans and chimpanzees differ by 4%, and most of these differences reflect insertions and deletions; yet, aging begins later or is extended further in humans than in chimpanzees. 49 From comparative genetics studies, de Magalhães and colleagues projected that aging did not necessarily result from programmatic sequential changes in gene expression or epigenetics, but they also found that the genetic changes observed between species in relation to aging were not entirely the result of random chance. 61 They further suggested that multiple genetic regulatory processes established early in life and important during development are less subjected to selective pressure after reproduction and may become detrimental later in life. Thus, they hypothesized that natural selection pressures that occurred after humans and chimpanzees diverged mainly affected pathways for development and growth that subsequently also affected the longer life span in humans. 8,62,75
Epigenetics is the study of changes in gene expression arising from changes in the genome that do not involve alterations in the DNA sequence, may be heritable, and are affected by aging. These include classic epigenetic mechanisms such as DNA methylation, histone modifications, and the more recently discovered noncoding RNAs (summarized in Table 2). 59,228 Distinct, as well as global, changes in the epigenomic landscape occur during aging and cause aberrant gene expression contributing to a wide range of pathologic conditions, such as cancer, diabetes, obesity, and cardiovascular and neurodegenerative diseases. 59,228
Epigenetic Changes and Aging.
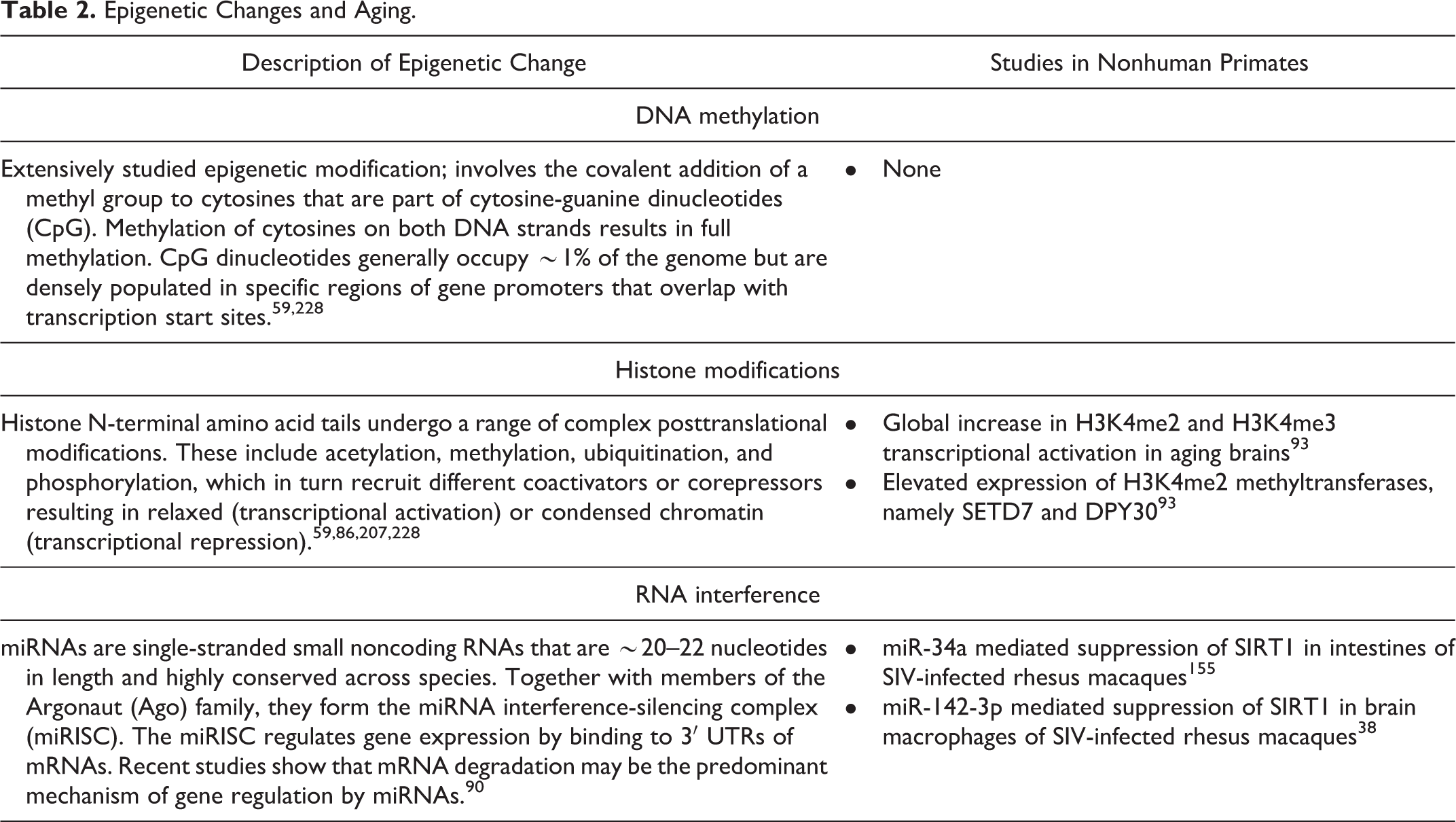
Aging is associated with genome-wide alterations in the distribution of 5-methylcytosine in such a way that DNA methylation levels are decreased globally. 113 While DNA methylation analysis has not been performed in nonhuman primates, increased gene expression of glucocorticoid receptor coincides with aging in rhesus macaques. 27 The use of peripheral blood leukocytes is less invasive and experimentally convenient, but this approach has limitations, as it does not accurately reflect epigenetic changes occurring in the target tissues in vivo. The availability of the complete rhesus macaque genomic sequence; the physiologic, anatomic, and immunologic similarities between rhesus macaques and humans; and the feasibility for repetitive and longitudinal sampling from multiple tissues offer unparalleled advantages for using rhesus macaques in characterizing aging-related epigenetic changes affecting glucocorticoid receptor gene expression and for future testing of novel glucocorticoid receptor mimetics.
Histone modifications represent another well-studied epigenetic change that involves remodeling at the N-terminal group of lysine residues by various modifications such as acetylation, deacetylation, methylation, ADP-ribosylation, ubiquitination, and the like. 59,86,207,228 Among these, histone acetylation, deacetylation, and methylation are considered the most predominant and best characterized posttranslational modifications. In rhesus macaque brains, aging is associated with a global increase in H3K4me2 and H3K4me3 transcriptional activation, suggesting the existence of open and active chromatin at transcription start sites and enhancer regions in the genome. 93 In addition, H3K4me2 methyltransferases, namely SETD7 and DPY30, show elevated expression. 93 The finding that H3K4me3 levels and the expression of its methyltransferases increase in yeast in response to double-stranded DNA damage response suggests a direct role for DNA damage response in triggering these specific modifications in brains of aged rhesus macaques. 71 Performing brain and other vital/specialized organ-focused epigenetic studies in humans is very challenging, and the findings from this study highlight the feasibility and profound contributions that nonhuman primate models can make toward understanding aging-associated epigenetic changes in specific organs/tissues of interest over time. 71
More recently, noncoding RNAs and microRNAs (miRNAs) in particular have been demonstrated to regulate the expression of genes linked to the aging process and include miR-34abc, miR-155, miR-21, miR-146a, miR-106ab, and miR-29ac. 94 Although studies linking specific miRNAs to the aging process in rhesus macaques have yet to be performed, we and others have identified specific miRNAs that negatively regulate the anti-inflammatory and antiaging SIRT1 expression in the intestine 155 and brain 38 of chronic SIV-infected rhesus macaques. Increased miR-142-3p expression in brain macrophages resulted in decreased SIRT1 expression leading to macrophage activation and encephalitis. 38 Similarly, we identified the miR-34a-SIRT1-acetylp65 axis to be a potential mediator of immune activation and persistent inflammation, also known as “inflamm-aging,” in the intestine. 155
Conclusion
Nonhuman primates continue to provide valuable information for elucidating aging processes that also occur in humans, and their use in biomedical and psychosocial studies is expected to further contribute to improvements in healthy aging. While the majority of studies related to gerontology utilize rhesus and cynomolgus macaques, improvements in animal husbandry practices will likely expand the use of smaller nonhuman primates. With the growing development of antiaging strategies, it is expected that nonhuman primates will additionally be highly relevant for preclinical studies testing antiaging strategies.
Footnotes
Notes
Declaration of Conflicting Interests
The authors declare that they have no conflicts of interest with respect to their authorship or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding support was from the National Institutes of Health (OD01104 to all authors; R01DK083929 to M.M.; AI097059, AI110163, HL125054, and AI116198 to M.J.K.), and the Tulane Research Enhancement Fund (M.J.K. and E.S.D.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.