
Abstract
Ependymal cells are epithelial support cells that line the central canal and ventricular cavities of the central nervous system, providing the interface between the cerebrospinal fluid and the parenchyma of the brain and spinal cord. The spinal ependymal layer (SEL) is composed of 3 main cell types: tanycytes, ependymocytes, and cerebrospinal fluid–contacting neurons. A fourth cell type, termed the supraependymal cell, is also occasionally described. Cells of the SEL show restricted proliferative capacity in health but display neural stem cell properties both in vitro and in vivo in various disease states. A growing body of literature is devoted to the regenerative roles of the SEL, particularly in the context of spinal cord injury, where mechanical damage to the spinal cord leads to a significant increase in SEL proliferation. SEL-derived cell progeny migrate to sites of injury within the injured spinal cord parenchyma and contribute primarily to glial scar formation. In additional to their role as endogenous neural stem cells, cells of the SEL may be an important source of cytokines and other cell signaling molecules, such as tumor necrosis factor, heat shock proteins, and various growth factors. The SEL has become of recent interest to neuroscience researchers because of its potential to participate in and respond to diseases affecting the spinal cord (eg, traumatic spinal cord injury) and neurodegenerative disease. The intimate association of the SEL with the cerebrospinal fluid makes intrathecal therapies a viable option, and recent studies highlight the potential promise of treatments that augment SEL responses to disease.
Keywords
Ependymal cells are epithelial support cells that line the central canal and ventricular cavities of the central nervous system (CNS). They form the interface between the cerebrospinal fluid (CSF) and the brain and spinal cord parenchyma. Specialized groups of ependymal cells make up the choroid plexus and circumventricular organs of the brain. Much of what is known about the embryogenesis, structure, and function of ependymal cells focuses on the population located within the ventricular system of the brain, while a smaller body of literature exists related to the spinal ependymal layer (SEL) and its role in health and disease. 27,50,69 The SEL has, however, become of recent interest to neuroscience researchers because of its potential to participate in and respond to various diseases affecting the spinal cord. The goal of this review is to summarize the basic science and veterinary-specific literature related to the SEL and its role in disease. This review does not address neoplastic processes of the SEL, as the reader can be referred elsewhere in the veterinary literature for recent descriptions of spinal ependymal neoplasms in the dog and cat. 51,74
Cell Types and Morphology
The SEL is the ependymal lining of the spinal cord central canal. Ependymal cells are embryonically derived from neuroepithelial cells of the neural plate and arise from neural precursor cells at various time points in gestation, depending on the species. 11,46
The histologic characteristics of the normal SEL in lower vertebrates, rodents, dogs, cattle, nonhuman primates, and humans have been described. 1,12,38,43,56,58,60,61 It is a common misconception that the SEL is a homogeneous monolayer of cuboidal epithelial cells. Actually, the adult mammalian SEL is composed of at least 3 distinct cell types: ependymocytes, tanycytes, and CSF-contacting neurons (CSF-CNs; Fig. 1). While the functional and immunohistochemical (IHC) properties of these cell types remain to be defined, differences in morphology are readily apparent.
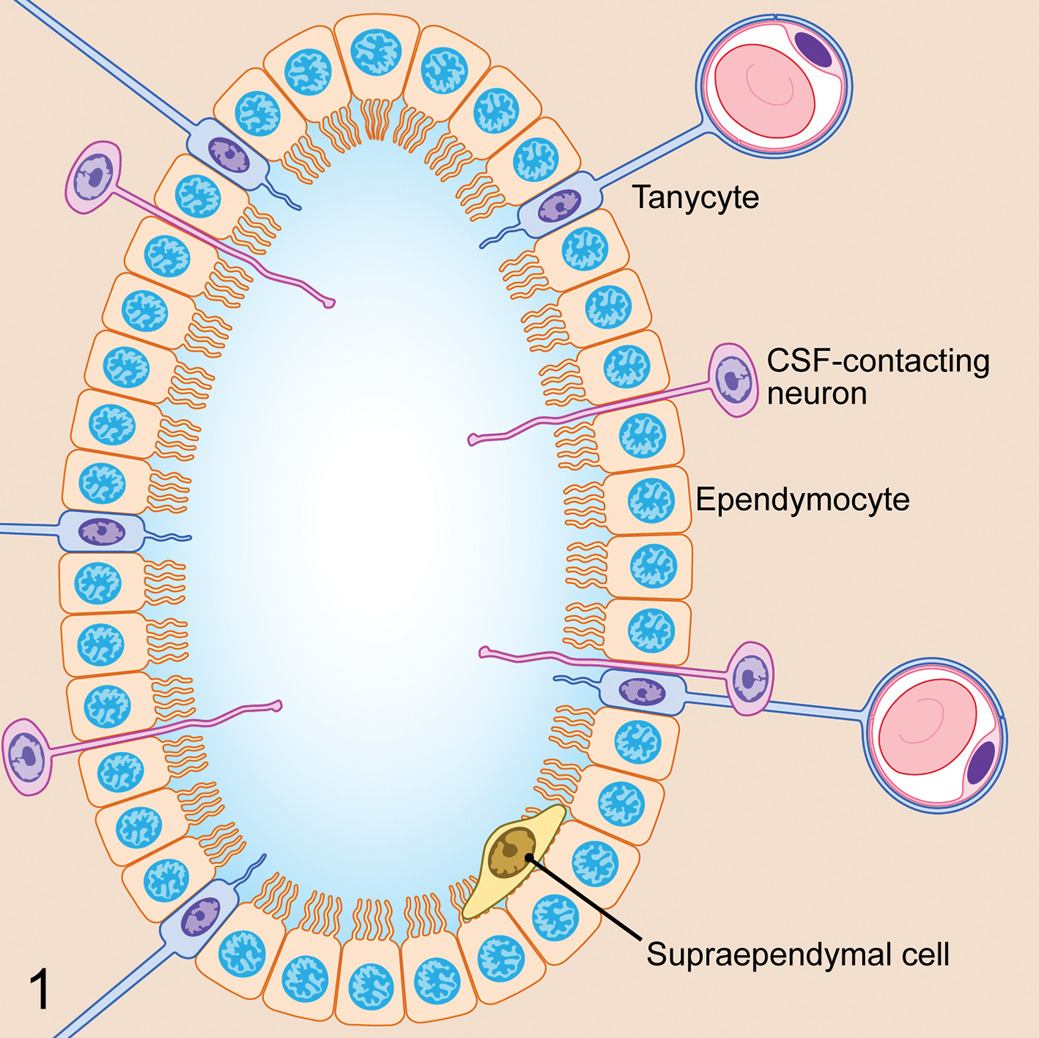
The spinal ependymal layer consists of 3 basic cells types: ependymocytes, tanycytes, and cerebrospinal fluid (CSF)–contacting neurons. A fourth cell type, the supraependymal cell, is also occasionally described in the literature.
Ependymocytes are the most plentiful cell type in the SEL, with a cuboidal epithelial morphology and 1 to 4 luminal cilia (Fig. 2). 26,40 These cells lend structural support to the spinal cord central canal, but additional roles of ependymocytes are poorly defined in contrast to what is known about other cell types in the SEL.
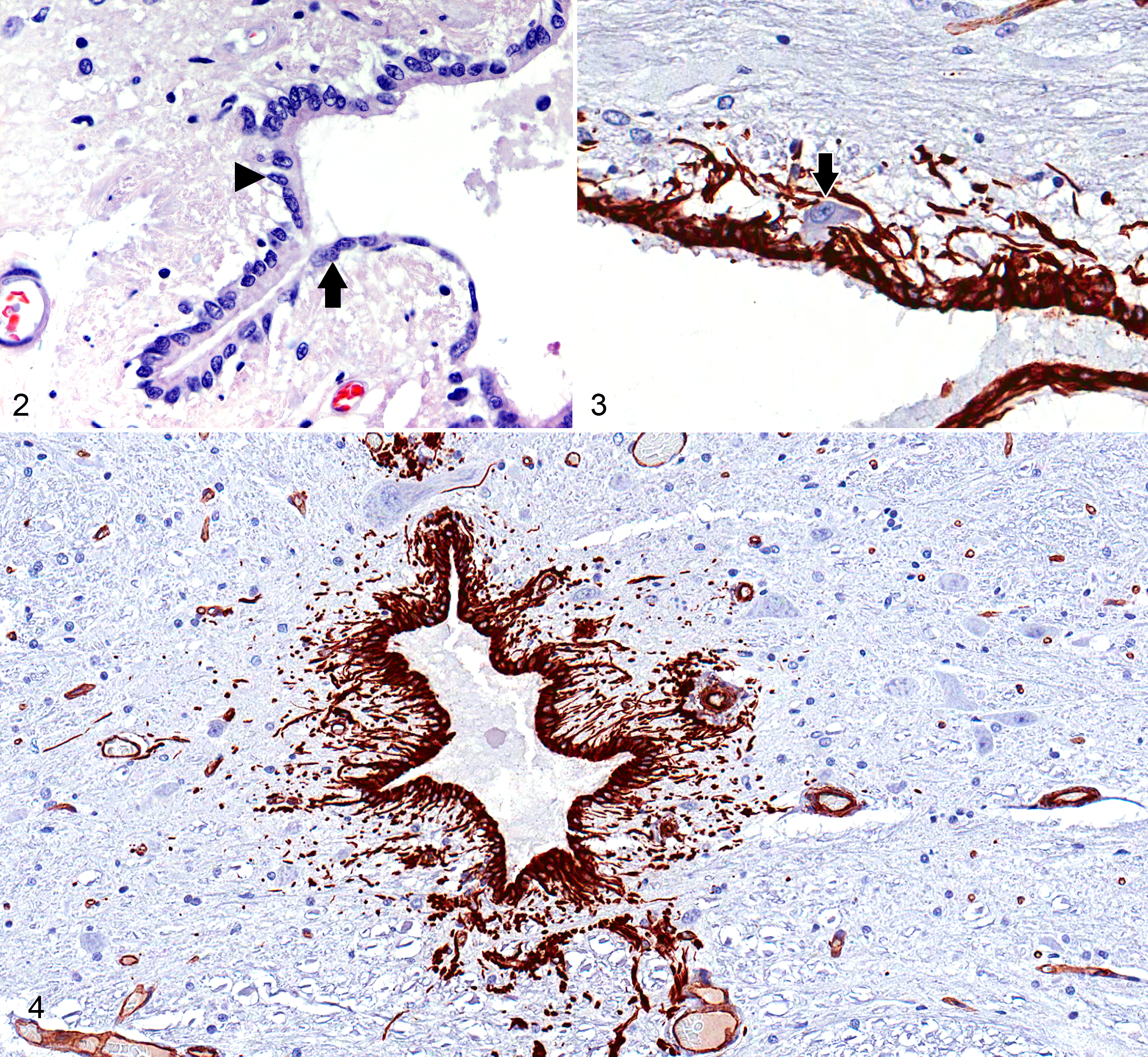
Midthoracic spinal cord (T7), normal dog.
Tanycytes are radial glial cells with a single luminal cilium and long basal cell processes stretching from the SEL to terminate on blood vessels within the spinal cord parenchyma (Fig. 2). 8 These cells facilitate transport of bioactive substances between the CSF and blood vessels by maintaining physical contact between the 2 compartments. Many tanycytes express vasoactive intestinal peptide, indicating an important role in the control of spinal cord vascular tone. 26 At least 4 subtypes of ventricular ependymal tanycytes have been described, each having a functional relationship with a subpopulation of neurons; however, less information is available regarding subtypes of spinal ependymal tanycytes. 57
CSF-CNs are a neuronal cell population found within the SEL of most species. These cells bear strong morphologic resemblance to bipolar neurons in the retina (Fig. 3). They have an irregular distribution around the central canal, with cell bodies that are located either within or adjacent to the layer of cuboidal ependymal cells. CSF-CN are plentiful in lower vertebrates, such as fish and amphibians, but are also reported in higher mammals, such as cats and dogs. 26,32,43,55,70 In most species, the highest concentration of CSF-CN can be found in the filum terminale of the spinal cord. 71,72 The luminal pole of these cells is a short dendrite with a ciliated surface that makes direct contact with CSF. The basilar pole behaves as an axon that radiates from the SEL toward the superficial regions of the spinal cord, in some cases extending as far as the meninges. 71 CSF-CNs function via nonsynaptic signal transmission and are believed to be a type of sensory neuron, possibly sensing changes in CSF composition, pressure, and flow. IHC studies have demonstrated numerous bioactive substances within these neurons, including acetylcholine esterase, GABA, vasoactive intestinal peptide, and somatostatin. 71,72 CSF-CNs also express ATP receptors such as P2X2 and respond to ATP currents. Changes in CSF pH excite CSF-CN, suggesting that these cells may contribute to homeostasis of CSF pH. Recently, the presence of calcium-permeable polycystic kidney disease 2-like 1 (PKD2L1) channels has been shown to be a specific and highly conserved marker for CSF-CNs in the embryonic and adult spinal cord, indicating a proprioceptive sensory function for these neurons. 18
A fourth pleomorphic group of cells, termed supraependymal cells, has also been reported in the SEL in species ranging from amphibians to humans. 34 Supraependymal cells are located within the central canal but are intimately associated with the luminal surface of the SEL. These cells are also associated with the choroid plexus of the third and fourth ventricles where they are referred to as epiplexus or Kolmer cells. 3 While the roles of this cell population in the SEL are undefined, their counterpart in the choroid plexus and ventricular ependyma displays a macrophage phenotype and is thought to play an important role in immunosurveillance. 3,34 It is currently unknown whether supraependymal cells exist in the canine SEL.
Immunohistochemical Properties
Study of the SEL has been hindered by difficulties in cell identification. Most studies have identified these cells morphologically and based on proximity to the spinal cord central canal. 43,64 This becomes problematic in scenarios such as traumatic spinal cord injury (SCI), where the SEL may become disrupted or cells of the SEL may migrate away from the central canal.
An assortment of IHC markers has been evaluated in the SEL of vertebrate animals and is summarized in Table 1. The intermediate neurofilament vimentin is expressed strongly in the SEL of most species (Fig. 4). 1,24,31,43 Glial fibrillary acidic protein (GFAP) is a marker of tanycyte lineage in the ventricular ependyma, and GFAP-positive cells have been observed with varying frequency in the mammalian SEL, depending on the species. 1,19,24,54 For example, a distinct population of GFAP-expressing cells has been observed in the SEL of adult mice, while GFAP expression is extremely limited in the SEL of healthy adult dogs but increases significantly after SCI (Figs. 5, 6). 20,43 Because most cells of the SEL are of neural crest origin, this cell layer is almost universally positive for S100 in all species (Fig. 7).
In Vivo Expression of Immunohistochemical Cell Markers in the Quiescent Adult SEL, With Reference to Species Evaluated.

Abbreviations: D, dog; H, human; M, mouse; Ma, macaque; NA, not applicable; R, rat; SCI, spinal cord injury; SEL, spinal ependymal layer.
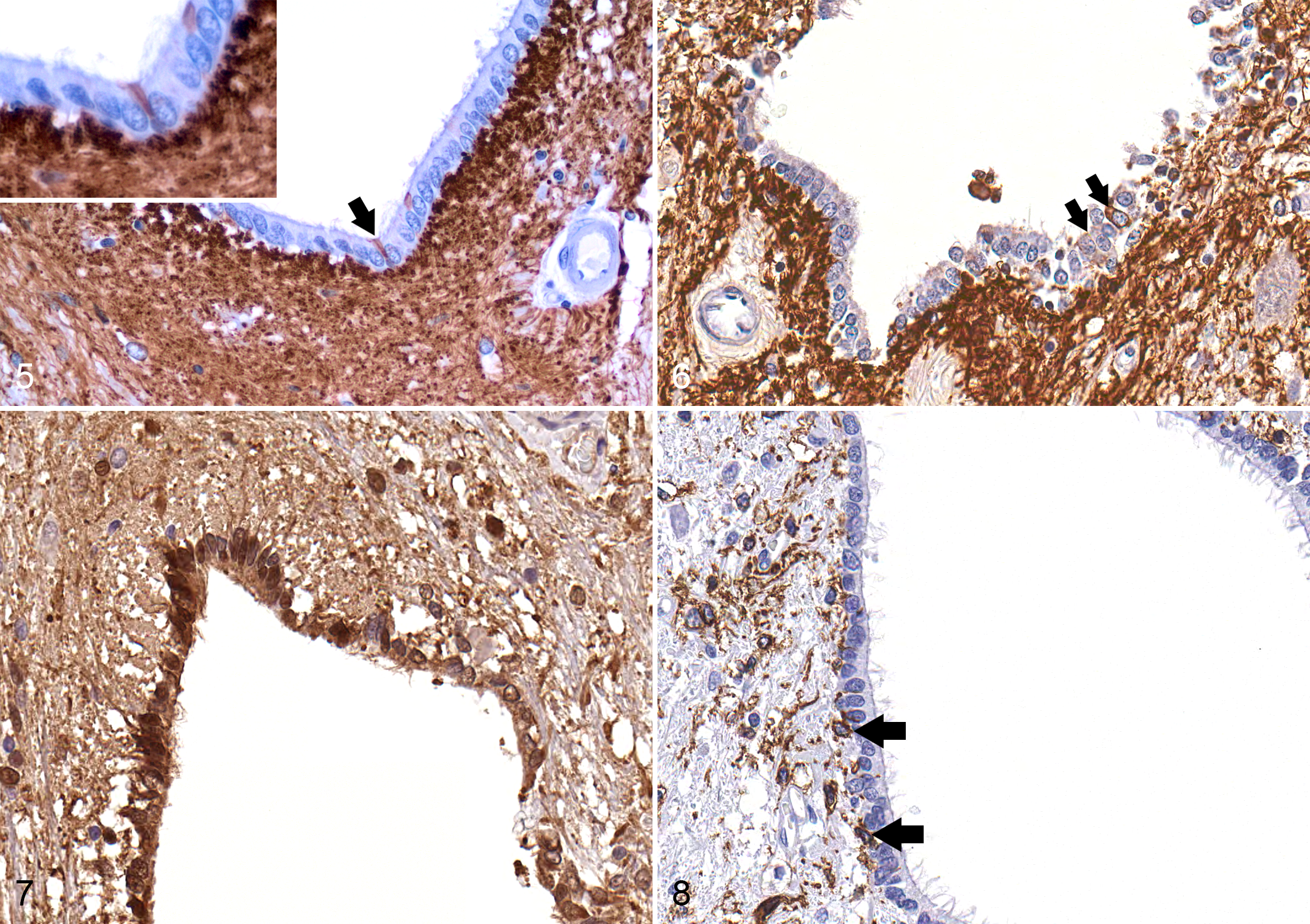
Several markers of neural precursor lineage are also expressed in the SEL, including nestin and SOX2. 1,19,24,54 Recently, activating transcription factor 3 (ATF3) has been reported to be a novel nuclear marker for migrating ependymal stem cells in the rodent spinal cord. 41 ATF3 is a member of the ATF/cAMP responsive element-binding (CREB) protein family of transcription factors, which participates in control of the cell cycle, plays a role in apoptosis, and facilitates invasiveness and mobility of cancer cells. 42,68,73 In the normal rodent spinal cord, ATF3 staining was confined to the cytoplasm of quiescent ependymal cells, but a nuclear staining pattern was observed when cells became “activated’ in the context of SCI. 42 This allowed the investigators to track cells of the SEL that had mobilized and migrated into the injured spinal cord parenchyma. Nestin, SOX2, and ATF3 expression has not been reported in the canine SEL, but the expression patterns of these markers in both health and disease warrant investigation.
Proliferative and Regenerative Capacity
Former dogma says that the CNS of adult mammals is a static tissue with limited proliferative capacity. However, we now know that there is much more regenerative capacity and plasticity within the CNS than originally thought. In fact, the production of new neurons occurs continually over a lifetime from stem cell populations that reside in the areas adjacent to the lateral ventricles, in the subventricular zone (SVZ). 79 The SVZ is composed of a mixture of astrocytes, quiescent ependymal cells, blood vessels, neuroblasts, and microglia. 24 In vitro it is these SVZ astrocytes that exhibit neural stem cell activity, and these cells have been shown to produce migratory neuroblasts that replenish interneurons of the olfactory bulb in the healthy state. 24 While neurogenesis does not take place in the normal spinal cord, recent work has indicated that specific cell populations within the adult mammalian spinal cord are capable of proliferation in health and in response to certain disease states and that a neural stem cell population resides within and around the SEL. 11,26,31,64
The proliferative capacity of the SEL is well described in lower vertebrates such as salamanders, where complete regeneration of the spinal cord is possible after a crush or transection injury. 2,16,33,49,78 This regeneration occurs through differentiation of neural precursor cells located in the SELs that become activated after injury and proliferate to replace lost tissue. 16,30,65 After spinal cord transection, cells of the salamander SEL show increased expression of the highly conserved stem cell marker nestin and migrate to the site of injury to participate in reparative neurogenesis. 78
Given the proliferative capacity of the SEL in lower vertebrates, the capacity for proliferation of the mammalian SEL is of great interest. Cells of the mammalian SEL show restricted proliferative capacity in health but display neural stem cell properties both in vitro and in vivo in various disease states. 6,29,40,53 Cells of the SEL are generally in a quiescent state in the healthy spinal cord but can become activated by various physiologic signals, including Ngn2, Notch1, BMP4, and Numb. 13,76 Proliferation of cells in the SEL under normal circumstances is minimal, confined predominantly to the dorsal aspect of the SEL and producing only other cells of ependymal lineage that remain within the SEL. 6 The role of proliferation in this scenario appears to be simply to maintain a healthy cell population within the SEL. However, under pathologic conditions such as that after SCI, cells of the SEL show marked proliferative capacity and produce progeny with multilineage potential that migrate into the parenchyma of the injured cord. 6,9,60,64 Fate-mapping studies have shown that almost all cells isolated from the spinal cord after SCI with stem cell properties are derived from the SEL. 6 This activation of cells in the SEL and the subsequent proliferative response are a widespread phenomenon within the injured cord, occurring both at the lesion epicenter and at distant sites within the CNS. 12,43,64 For these reasons, cells of the SEL present an attractive endogenous alternative to stem cell transplantation. 7,39
A growing body of literature is devoted to the regenerative roles of the SEL in various disease processes affecting the spinal cord, with perhaps the most focused effort involving SCI. Even in the normal spinal cord, cells of the SEL express SOX2, a transcription factor associated with neural progenitor identity. 21 The proliferative capacity of cells of the SEL increases dramatically within the injured cord, where cells of the SEL show enhanced neural stem cell properties. 6,40,60 SCI causes mechanical damage to the central canal, which leads to a significant increase in SEL proliferation, enhanced expression of nestin and SOX2 within the SEL, and ultimately, migration of SEL-derived progeny to sites of injury within the spinal cord parenchyma. 21,22,47,48 Lacroix et al 31 recently evaluated proliferative patterns of the SEL in a rodent model of contusion SCI. Cells of the SEL showed a rapid and bimodal time course for proliferation at the lesion epicenter, peaking first at 3 days after injury and then again at 14 days after injury. Rostral to the lesion epicenter, SEL proliferation occurred more gradually and peaked at 14 days after injury. SEL proliferation did not occur at any time point in sections of spinal cord caudal to the lesion epicenter. This group also evaluated chemically induced and autoimmune demyelination and reported no evidence of SEL proliferation in either of these models, despite reports from other groups that ependymal cell proliferation may occur under these circumstances. 14,15
In the rat, millions of ependymal cell progeny are produced in the weeks following contusion SCI. 5,31,45 This wave of proliferation produces primarily cells of astrocytic lineage, which form the glial scar, while a much smaller portion of ependymal progeny displays an oligodendrocyte phenotype. 40 This is in contrast to in vitro behavior of cells of the SEL, where ependymal cells activated by SCI produce significant numbers of astrocytes, oligodendrocytes, and even motor neurons. 6,45,50 Clearly the microenvironment within the injured spinal cord influences differentiation of ependymal cell progeny, and efforts are currently focusing on in vivo manipulation of endogenous ependymal cells to enhance oligodendrocyte or neuronal differentiation.
Rodent studies have also previously shown that transplantation of ependymal-derived neurospheres cultured from spinal cord–injured donor animals results in improved neurologic recovery after SCI. 45 Histopathologically, ependymal cell transplantation resulted in reduced cavitation within the injured cord. Transplanted ependymal cells were observed to migrate long distances rostral and caudal to the site of implantation, crossing the lesion epicenter. Interestingly, these transplanted cells rarely showed evidence of differentiation within the injured cord, suggesting that paracrine and immunomodulatory mechanisms are the primary reason for improvement in tissue repair and outcome. 45,75
Proliferation of cells of the SEL has been studied to a lesser extent in models of neurodegenerative disease, such as amyotrophic lateral sclerosis (ALS); however, ependymal cells may play an important role in this context as well. Marcuzzo et al 37 recently evaluated the proliferative capacity of the SEL in G93A-SOD1 mice, a rodent model of ALS. This group found that cells of the SEL from affected mice maintained their multipotent potential in cell culture but that SEL progeny contained a higher proportion of neurons when compared with control animals. This finding correlated with changes in micro-RNA expression that would be expected to drive cell fate toward neuronal differentiation. 37
Secretory Roles
Other recent work highlights important questions regarding potential secretory roles of cells in the SEL. 45 In addition to their role as endogenous neural stem cells, cells of the SEL may be an important source of cytokines and other cell signaling molecules. Ventricular ependymal cells serve as an important source of tumor necrosis factor in systemic inflammation and as a source of interleukin 8 (IL-8), inducible nitric oxide synthase, and heat shock protein 70 in both health and disease. 25,44,63,66 Enhanced expression of IL-1β, tumor necrosis factor, and heat shock protein 70 in the SEL has been observed in laboratory models of injury and in a spontaneous canine model of SCI caused by intervertebral disc extrusion. 4,43,56 Changes in protein expression in the SEL are reflected in CSF composition. Liberation of proinflammatory cytokines and damage-associated molecular patterns from ependymal cells in the injured spinal cord has the potential to modulate the inflammatory response both locally and globally within the CNS. An increased number of CD18+ cells has been observed in the SEL of dogs with the neurodegenerative disease canine degenerative myelopathy, which is considered a spontaneous large animal model of familial ALS (Fig. 8). 35 The significance of this finding is not completely understood, but these cells may affect the neural stem cell niche known to reside within and adjacent to the SEL. 4,43
Beyond immunomodulatory roles, cells of the SEL may serve as important paracrine mediators and sources of growth factors within the CNS. Cells of the SEL have been reported to express growth factors, such as brain-derived neurotrophic factor, glial cell line–derived neurotrophic factor, and nerve growth factor, all of which foster a favorable environment for cell survival and axonal regrowth after injury. 10,21 After SCI, expression of brain-derived neurotrophic factor in the SEL increases significantly, supporting the potential role of the SEL in tissue recovery beyond its inherent regenerative capacity. 21,67 Conversely, enhanced expression of purinergic receptors such as P2X7 by ependymal cells after SCI may inhibit neurogenesis and induce cell necrosis and apoptosis, indicating that injured ependymal cells may negatively influence cell survival after CNS injury. 23,41 In rodent models of SCI, administration of drugs that inhibit P2X7 receptor function has shown promise in enhancing recovery and reducing detrimental gliosis and glial scar formation. 36,52
Targeting Ependymal Cells for Therapy
Ependymal cells have recently become therapeutic targets in the treatment of CNS diseases, such as SCI, multiple sclerosis, and ALS. Interventional mechanisms range from viral vector gene and cytokine therapies in ALS and multiple sclerosis to the administration of growth factors to stimulate endogenous neural stem cell proliferation in SCI. 39,75 Viral vector studies show preferential infection of ependymal cells by replication-deficient herpes simplex virus type I and adeno-associated viral vectors, with subsequent long-term therapeutic gene expression in various inflammatory and neurodegenerative disease models. 17,28,39,77
In many ways, the SEL is an ideal therapeutic target in spinal cord disease. Because of its intimate contact with the CSF, it is highly amenable to intrathecal drug delivery. This can be accomplished via a simple lumbar or cerebellomedullary cistern puncture in the clinical setting, depending on the spinal cord region of interest. Drug delivery by this route avoids the limitations imposed by the blood-brain barrier, which is often difficult to penetrate, even when significant CNS inflammation is present. 39
Intrathecal delivery of growth factors or other cytokines to stimulate or influence endogenous stem cell proliferation within the SEL is relatively noninvasive when compared with direct implantation of exogenous stem cells, which often requires a surgical approach to the vertebral column. Additionally, recruiting and modulating endogenous stem cells for tissue regeneration circumvents feasibility issues related to culture of autologous and exogenous stem cells and avoids the ethical issues surrounding embryonic stem cell therapies. 59 Several recent editorials have highlighted the potential utility of targeting ependymal cells as a noninvasive therapy for neurologic disease, and interest in this area will likely grow in the coming years. 39,62
Conclusions
The growing body of scientific literature dedicated to the SEL highlights this cell population as an important player in cell-based therapies to treat spinal cord disease. The SEL is uniquely positioned to provide and support proliferative responses after SCI and to exert both local and global influences within the CNS. The intimate association of the SEL with the CSF makes targeted intrathecal therapies a viable option. Several recent studies in rodent models have highlighted the potential promise of treatments aimed at augmented SEL responses to disease. Future studies will continue to focus on treatments to drive oligodendrocyte and neuronal differentiation after SCI and should work to further define the secretory roles of this cell population in health and disease.
Footnotes
Acknowledgements
We gratefully acknowledge Ms Florinda Jaynes for her assistance with immunohistochemical staining and Mr Tim Vojt for his assistance with figure preparation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received the financial support for the research, authorship, and/or publication of this article: Funding for this work was provided in part by a grant from NCI P30CA016058.