
Abstract
The spinal cord ependymal layer (SEL) is a recent focus in spinal cord injury (SCI) research because of its potential to serve as a source of endogenous neural stem cells. Dogs are an important spontaneous model of SCI; however, there is a paucity of information available in the literature regarding the canine SEL. Here we describe the histologic appearance and immunohistochemical staining patterns of the SEL in normal dogs (n = 4) and dogs with acute SCI caused by intervertebral disk extrusion (n = 7). Immunohistochemical staining for PCNA, Ki-67, caspase 3, E-cadherin, GFAP, and vimentin was employed in both groups. Staining for Ki-67 was absent in the SEL of normal and SCI-affected dogs, indicating possible restricted proliferative capacity of the canine SEL acutely after SCI. GFAP-positive cells were increased after SCI at both at the lesion epicenter and at proximal spinal cord sites (P = .001 and P = .006, respectively), supporting the possibility of astrocytic differentiation within the SEL after SCI. Total E-cadherin staining did not differ between normal and SCI-affected dogs (P = .42 for lesion epicenter, P = .09 at proximal sites) and was restricted to the apical cell surface in normal dogs. After SCI, E-cadherin staining was membrane-circumferential and cytosolic in nature, indicating possible loss of cellular polarity after injury that could drive cell migration from the SEL to injury sites. Enhanced GFAP expression and changes in E-cadherin expression patterns support additional studies to evaluate the canine SEL as a source of endogenous neural precursors that may be modulated for future clinical interventions after SCI.
The spinal cord ependymal layer (SEL) and periependymal region have become a recent focus in spinal cord injury (SCI) research because of the potential for this region to serve as an endogenous source of neural stem cells to aid in tissue regeneration and repair. Cells of the SEL line the spinal cord central canal and are derived from neuroepithelial stem cells of the ventral neural tube. 9
It is a common misconception that the SEL is composed of a homogeneous monolayer of cuboidal epithelium. In vertebrate species, the SEL actually consists of at least 3 distinct cell types: tanycytes, cuboidal ependymal cells, and cerebrospinal fluid (CSF)–contacting neurons. 16 Tanycytes are elongated, nonbranching, uni- or bipolar cells that radiate from the central canal into the gray matter and frequently terminate on blood vessels. Tanycytes are thought to facilitate communication between CSF and capillaries. 16,39 Cuboidal ependymal cells or ependymocytes are ciliated cells connected together via gap junctions whose function in the normal and diseased spinal cord is poorly defined. 6 CSF-contacting neurons are a type of neuron observed frequently in the SEL of lower vertebrates such as fish and amphibians; however, their occasional presence has also been reported in rodents, cats, and primates. Structural and functional characteristics of these neurons are well conserved across species. 23,29,34,50 They display electrophysiologic properties similar to those of immature neurons and express markers of neuronal lineage such as NeuN and doublecortin. They also express P2X receptors, are thought to be involved homeostatic mechanisms that maintain pH in the CSF, and send collateral axons to other local neurons and to higher spinal segments. 29,45,50
In lower vertebrates, cells of the SEL play a prominent role in the regeneration observed after spinal cord transection. 8,27 In mammals, cells of the SEL actively proliferate during development but take on a quiescent state in the adult. 4 Occasional proliferation of cells in the SEL is noted in normal mice and appears to be concentrated at the dorsal aspect of the central canal, providing cells that generally remain within the ependymal layer. 14 Recent studies have also demonstrated that cells of the mammalian SEL have enhanced proliferative capacity under pathologic conditions such as SCI. 3,21 In rodents, cells of the SEL re-express neural stem cell markers and undergo a robust proliferative response after SCI. 33,35 In this scenario, SEL progeny migrate to the site of injury within the spinal cord parenchyma and exhibit differentiation into astrocytes and oligodendrocytes. These SEL-derived glial cells play vital roles in the formation of functional neuronal circuits and the glial scar. 3,28,30,38 In vitro, cells from the SEL of both mice and humans can also assume stem cell properties, generating multipotent neurospheres in culture. 1,7,28 Given these findings, experimental treatments that exploit the SEL as a source of endogenous neural stem cells are currently being explored in rodent models of SCI, with some success. 32 However, as with other promising treatments for SCI, translation from rodent models to human patients will present a challenging next step.
Dogs have a high incidence of spontaneous SCI, caused primarily by intervertebral disk extrusion (IVDE). 25,37 Primary and secondary mechanisms of SCI in dogs with IVDE bear strong similarity to those observed in humans with SCI. 15,44 Accordingly, dogs have gained favor as a “bridge” between rodent models of SCI and human patients because they address many translational issues. Specifically, dogs offer heterogeneity in lesion and patient-related variables, providing a situation that more closely approximates clinical trials of human SCI. 17,25 Spontaneous SCI in dogs offers a valuable clinical model to test therapeutic interventions targeting the cells of the SEL; however, to the authors’ knowledge, there are no published works that examine the normal histologic and immunohistochemical features of the canine SEL or its responses to naturally occurring SCI. Further study is needed to define the phenotypic characteristics of the canine SEL for future clinical and translational work related to SCI.
The goals of this study were to describe the normal histologic appearance and immunohistochemical staining patterns of the canine SEL and to examine proliferative responses to acute, spontaneously occurring SCI. Here we examine immunohistochemical staining for several markers of proliferation, apoptosis, and cell lineage in the SEL of spinal cord sections from neurologically normal dogs and compare these results with those seen in dogs with acute SCI caused by IVDE.
We chose to examine 2 markers of cellular proliferation: proliferating cell antigen (PCNA) and Ki-67. PCNA, also referred to as cyclin or DNA polymerase delta auxiliary protein, 36,48 is commonly used as a marker of cellular proliferation. PCNA was originally characterized as a DNA sliding clamp for DNA polymerases, and it is an essential player in DNA replication and cell cycle progression in eukaryotic cells. 46 However, other functions of PCNA have been documented in nonproliferating cells. In particular, PCNA is also expressed during DNA repair and possibly during RNA transcription, indicating that PCNA staining is not entirely specific as a marker for cell proliferation. 5,13,20,22,48 High numbers of PCNA-positive cells are reliably present in some quiescent cell populations, including normal epithelial cells located in the vicinity of malignant neoplasms, 51 Schwann cells of normal peripheral nerve, 26 and the SEL of other species including, the rat and human. 10,47 The PCNA staining pattern of the normal canine SEL has not been previously described. In contrast, Ki-67 is strongly expressed in all phases of the cell cycle except for G0 and is considered a more specific marker of proliferation than PCNA. 11,19,41 Assessment of PCNA expression in combination with Ki-67 expression can provide insight into both the proliferative and metabolic activity of a cell population, particularly when a cell population is positive for one marker but not the other.
We hypothesized that the canine SEL would be similar to that reported in other mammalian species with respect to cell morphologies and would exhibit proliferation after SCI, as evidenced by increased total SEL cell count and enhanced staining for markers of cellular proliferation. In addition, given available literature in other species, we hypothesized that many cells in the canine SEL would be PCNA positive in the resting state.
Materials and Methods
Tissue Samples and Histology
The normal morphology and immunohistochemical properties of the canine SEL were investigated in spinal cord tissue from healthy adult mixed-breed dogs ranging in weight from 19.6 to 30.2 kg (n = 4). This tissue was derived from dogs used in a separate and published study supervised and approved by The Ohio State University Institutional Animal Care and Use Committee. 2 Mid-thoracic spinal cord tissue samples from these dogs were immersion fixed in formalin and paraffin-embedded for subsequent histologic examination. For comparison, tissue from dogs euthanized because of acute SCI (n = 7) was examined. SCI-affected dogs represented routine necropsy submissions to The Ohio State University College of Veterinary Medicine diagnostic pathology service. The severity of clinical neurologic signs at the time of euthanasia was retrospectively quantified based on information in the patient medical record using a previously published scale ranging from 1 to 5, with a score of 5 indicating a dog with paraplegia and absence of nociception. 42,49 Additional clinical information retrospectively gathered from the medical records of SCI-affected dogs included breed, age at the time of euthanasia, duration of neurological signs prior to euthanasia, and cause and location of SCI. This information is summarized in Table 1. Spinal cord tissue was collected at the time of necropsy, immersion fixed in formalin, paraffin-embedded, and then processed for routine histologic evaluation with hematoxylin and eosin (HE) staining. Representative sections of spinal cord included lesion epicenter (variable between dogs) and a section of mid-thoracic spinal cord proximal to the lesion epicenter (T7–T9). The lesion epicenter in SCI-affected dogs was defined as the spinal cord segment that was subjectively the most severely compressed on gross evaluation at necropsy. This information was noted in the pathology report for each patient, and the spinal cord segment associated with this vertebral level was considered the lesion epicenter for the purposes of this study. Histopathologic abnormalities within the spinal cord of SCI-affected dogs included variable degrees of neuronal necrosis, axonal swelling and degeneration, suppurative inflammation, and gray matter hemorrhage (Table 1; Figs. 1, 2).
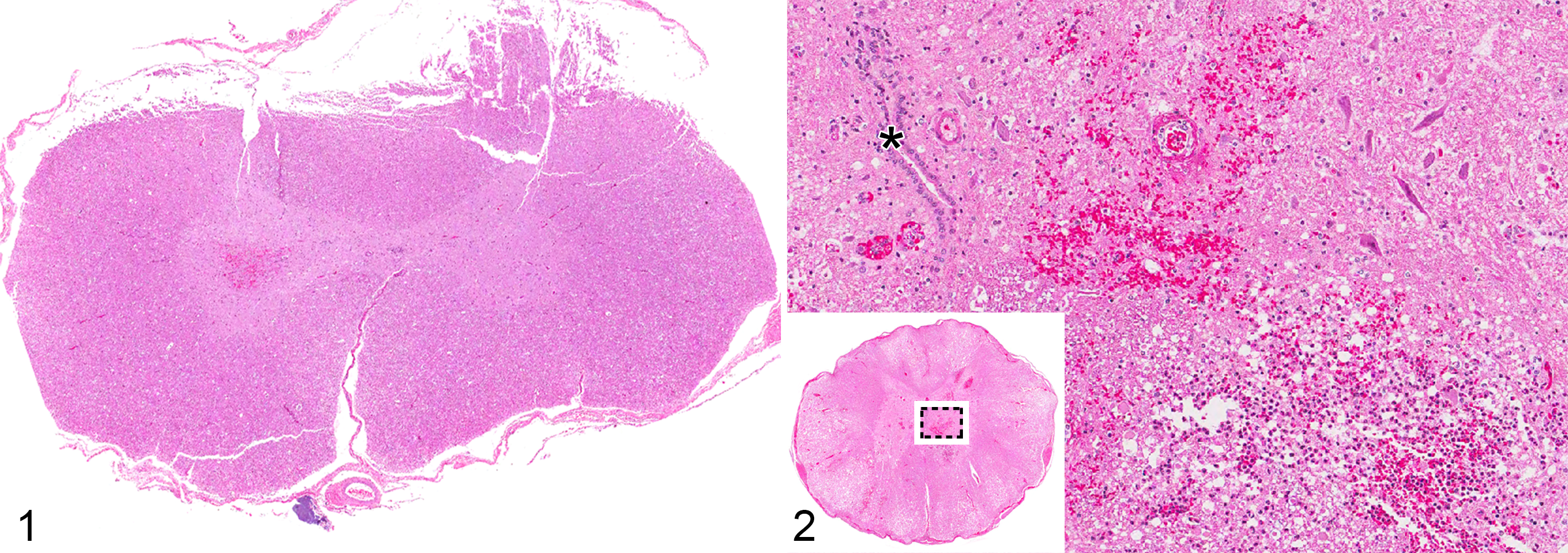
Acute spinal cord injury (SCI), thoracolumbar spinal cord (lesion epicenter, case Nos. 2 and 5, respectively), dog. Hematoxylin and eosin.
Spinal Cord Ependymal Layer (SEL) Responses to Traumatic Spinal Cord Injury Evaluated in Dogs With Naturally Occurring Intervertebral Disk Extrusion (IVDE).
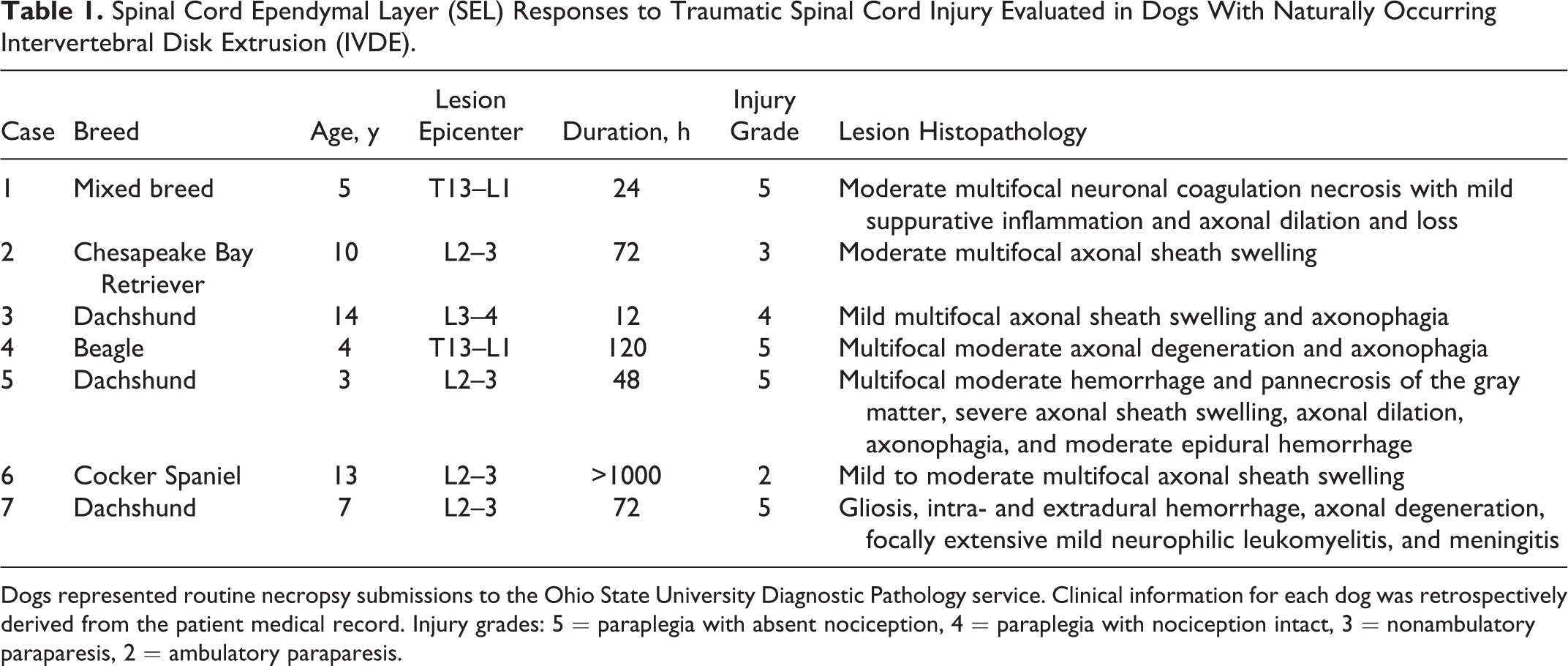
Dogs represented routine necropsy submissions to the Ohio State University Diagnostic Pathology service. Clinical information for each dog was retrospectively derived from the patient medical record. Injury grades: 5 = paraplegia with absent nociception, 4 = paraplegia with nociception intact, 3 = nonambulatory paraparesis, 2 = ambulatory paraparesis.
Quantitative Analysis of Spinal Ependymal Layer Cell Numbers
For both normal and SCI-affected dogs, the total number of cells in the SEL was quantified manually by a single blinded investigator (S.A.M.) based on HE-stained transverse sections. As previously reported by Takahashi et al 47 in 2003, cells making contact with the central canal and cells contacting the ependymal lining of the central canal with more than 50% of their circumference were considered part of the SEL. Normal values were established for SEL numbers in control dogs by evaluating 3 consecutive transverse sections of mid-thoracic spinal cord per dog. For SCI-affected dogs, the number of cells in the SEL was quantified in available transverse sections at the lesion epicenter, the location of which varied from dog to dog, and in mid-thoracic spinal cord sections proximal to the lesion epicenter.
Immunohistochemistry
Transverse sections of spinal cord from both SCI-affected dogs (lesion epicenter and proximal mid-thoracic site) and normal controls (mid-thoracic site) underwent immunohistochemistry (IHC) staining for markers of cell proliferation, cell lineage, and apoptosis. PCNA (Dako, Carpinteria, CA; monoclonal mouse anti-rat; 1:100) and Ki-67 (Thermo Fisher Scientific, Waltham, MA; monoclonal rabbit anti-human; 1:180) were used to assess for proliferation and metabolic activity. Caspase 3 (Cell Signaling Technology, Beverly, MA; polyclonal rabbit anti-human; 1:180) was used as a marker of apoptosis. Cell lineage and differentiation were assessed using E-cadherin as an epithelial cell marker (BD Biosciences, San Jose, CA; monoclonal mouse anti-human; 1:200), GFAP as a marker of glial lineage (Dako; polyclonal rabbit anti-cow; 1:100), and vimentin as a marker of mesenchymal cell lineage (Dako; monoclonal mouse anti-pig; 1:100). All IHC staining was performed concurrently in the Comparative Pathology and Mouse Phenotyping Shared Resource at The Ohio State University College of Veterinary Medicine. After deparaffinization, sections were treated with 3% hydrogen peroxide for 10 minutes. Species-specific biotin-conjugated secondary antibodies were used at working dilutions indicated above. Peroxidase labeling was visualized with 3,3-diaminobenzidine. Sections were counterstained with hematoxylin. Negative controls for IHC included tissue probed with non-hyperimmune total mouse or rabbit IgG.
Evaluation of IHC Images
Individual slides were digitized using the Aperio ScanScope (Aperio Technologies, Vista, CA), and digital images were analyzed using Aperio ImageScope software. Normal IHC staining patterns were established for the SEL in control dogs by evaluating 3 transverse sections of mid-thoracic spinal cord per dog. For SCI-affected dogs, the SEL was examined in transverse sections at the lesion epicenter, the location of which varied by dog, and at a mid-thoracic spinal cord segment proximal to the lesion epicenter.
For IHC stains that provided primarily nuclear staining (PCNA, Ki-67, and caspase 3), manual cell counts were performed by a single blinded investigator (S.A.M.) recording the number of positively stained cells in the SEL of each spinal cord section examined, then calculating a mean number of IHC-positive cells per dog. For IHC stains that provided primarily cytoplasmic staining (GFAP, vimentin, and E-cadherin), immunoperoxidase signal was quantified via positive pixel counting using the following procedure: for each slide examined, a region of interest (ROI) containing only the SEL was manually defined using 40× magnification on the ImageScope software. A previously published algorithm for digital quantitative analysis of IHC staining was used. 31 Positive pixel counts were performed for 3 representative areas within each ROI, and values were reported as “positivity” (calculated as the number of positive pixels identified by the algorithm in a defined field, divided by the number of total pixels in the field). Possible values for positivity ranged from 0 to 1.0. The values obtained for the SEL in each tissue section were averaged to produce a single “mean positivity” of the SEL for each dog.
Statistical Analysis
Descriptive statistics, including median and range, were calculated for continuous variables such as neurologic injury grade. Mean positivity, or mean number of positive cells from the group of SCI-affected dogs, was compared by t test to that of controls. All values are presented as mean ± standard error of the mean (SEM) where data were normally distributed, and values are reported as a median and range where it was not. For IHC findings that were statistically different between groups, additional analysis was performed via Pearson correlation to determine whether patient factors such as injury severity or duration of neurologic signs correlated with IHC findings. P < .05 was considered statistically significant for all tests.
Results
SEL Cell Types and Number
Consistent with what has been reported in other species, multiple morphologically distinct cell types could be readily identified in dogs and appeared similar to those previously reported in rodents. 30 These included tanycytes with oval to fusiform nuclei and single long slender cell processes that radiate from the central canal into the gray matter (Fig. 3); cuboidal ependymal cells with round to slightly oval nuclei, faintly staining cytoplasm and variable numbers of luminal cilia (Fig. 3); and CSF-contacting neurons (Fig. 4). The mean total number of ependymal cells present in transverse sections of the SEL in normal dogs was 136 ± 25.
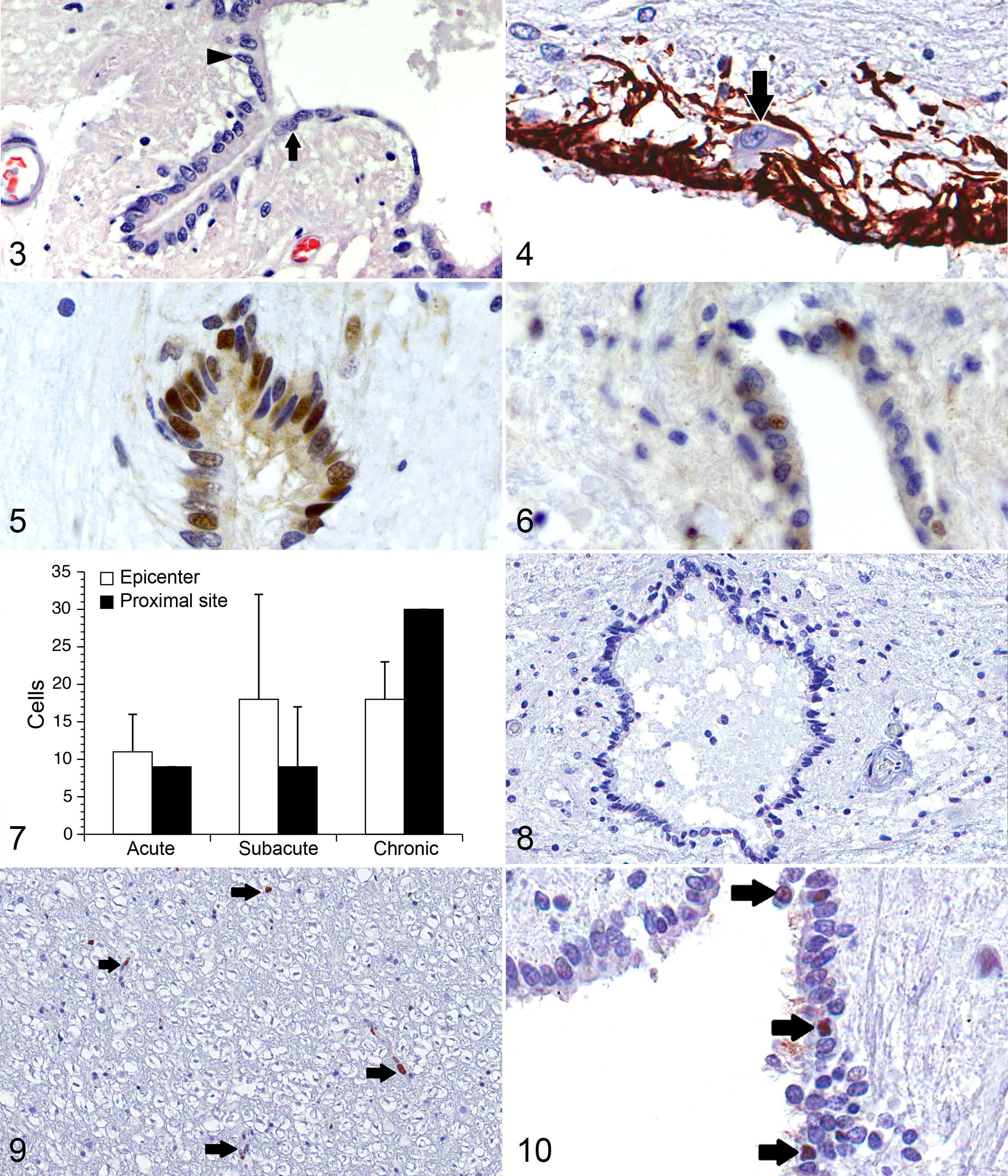
Normal mid-thoracic spinal cord, dog.
The mean total number of cells located in the SEL of SCI-affected dogs was higher both at the lesion epicenter and at proximal thoracic sites, 162 ± 25 and 160 ± 48, respectively; however, this finding was not statistically significant (P = .1 for both sites). There was a trend toward correlation between SEL total cell counts at the lesion epicenter and injury severity in SCI-affected dogs (P = .08), such that dogs with more severe injuries tended to have higher total SEL cell counts. A correlation was not identified between total SEL cell count and duration of injury, patient body weight, or age.
PCNA and Ki-67 Expression
Immunohistochemistry for PCNA and Ki-67 was employed to assess proliferative activity. Many cells in the SEL of normal dogs contained PCNA-positive nuclei (Fig. 5). Mean total number of PCNA-positive cells per transverse spinal cord section in normal dogs was 60 ± 7. Quantification of the dorso-ventral distribution of PCNA-positive cells in the normal SEL was performed by drawing a line through the middle of the central canal on transverse spinal cord sections and counting the number of PCNA-positive cells within the dorsal and ventral aspects of the SEL. There was no significant difference in the percentage of cells that were PCNA positive in the dorsal vs ventral aspects of the SEL (49% vs 45%, P = .23), although both mean total cell count (85 ± 8 dorsal vs 45 ± 9 ventral) and mean total number of PCNA-positive cells (42 ± 8 dorsal and 31 ± 8 ventral) were greater in the dorsal aspect of the SEL (P = .03 and .005, respectively). Mean total number of PCNA-positive cells identified in the SEL of SCI-affected dogs was 16 ± 7 at the lesion epicenter and 14 ± 6 at proximal sites. This number was significantly decreased at both the lesion epicenter and proximal sites compared with controls (P = .003 and .002, respectively) (Fig. 6). There was no correlation between the number of PCNA-positive cells at the lesion epicenter and injury severity or duration; however, the number of PCNA-positive cells at proximal thoracic sites was positively correlated with duration of injury (r = 0.931; P = .03) and was inversely correlated with injury grade (r = –0.993; P = .004), where a higher injury grade indicates a more severe neurologic injury. Figure 7 provides descriptive statistics regarding PCNA expression in acute (n = 2), subacute (n = 3), and chronically (n = 2) affected dogs. In contrast, the SEL was completely devoid of Ki-67–positive cells in both control and SCI-affected dogs (Fig. 8). Numerous Ki-67–positive cells were noted throughout the gray and white matter of SCI-affected dogs, primarily associated with vascular endothelium (Fig. 9).
Caspase 3 Expression
There were no caspase 3–positive nuclei in the SEL of any of the normal dogs examined; however, an occasional strongly caspase 3–positive nucleus was observed within the SEL of SCI-affected dogs (Fig. 10). This ranged from 0 to 5 at the lesion epicenter (median 1 cell per section) and 0 to 1 at proximal thoracic sites (median 0 cells per section). There was not a significant correlation between number of caspase 3–positive cells and injury severity or duration in SCI-affected dogs.
E-cadherin, GFAP, and Vimentin Expression
Staining for E-cadherin was localized exclusively to the apical surface of the SEL in normal dogs with a mean positivity of 0.214 ± 0.03 as measured by positive pixel count (Fig. 11). Mean positivity for E-cadherin in SCI-affected dogs was 0.324 ± 0.05 at the lesion epicenter and 0.432 ± 0.05 at proximal thoracic sites. There was not a statistical difference in E-cadherin positivity between control and SCI-affected dogs at either the lesion epicenter or proximal sites (P = .42 and .09, respectively); however, the distribution of E-cadherin staining differed between groups. Where control dogs displayed a strictly apical pattern of staining, SCI-affected dogs tended to display circumferential membrane and cytosolic staining of SEL (Fig. 12).
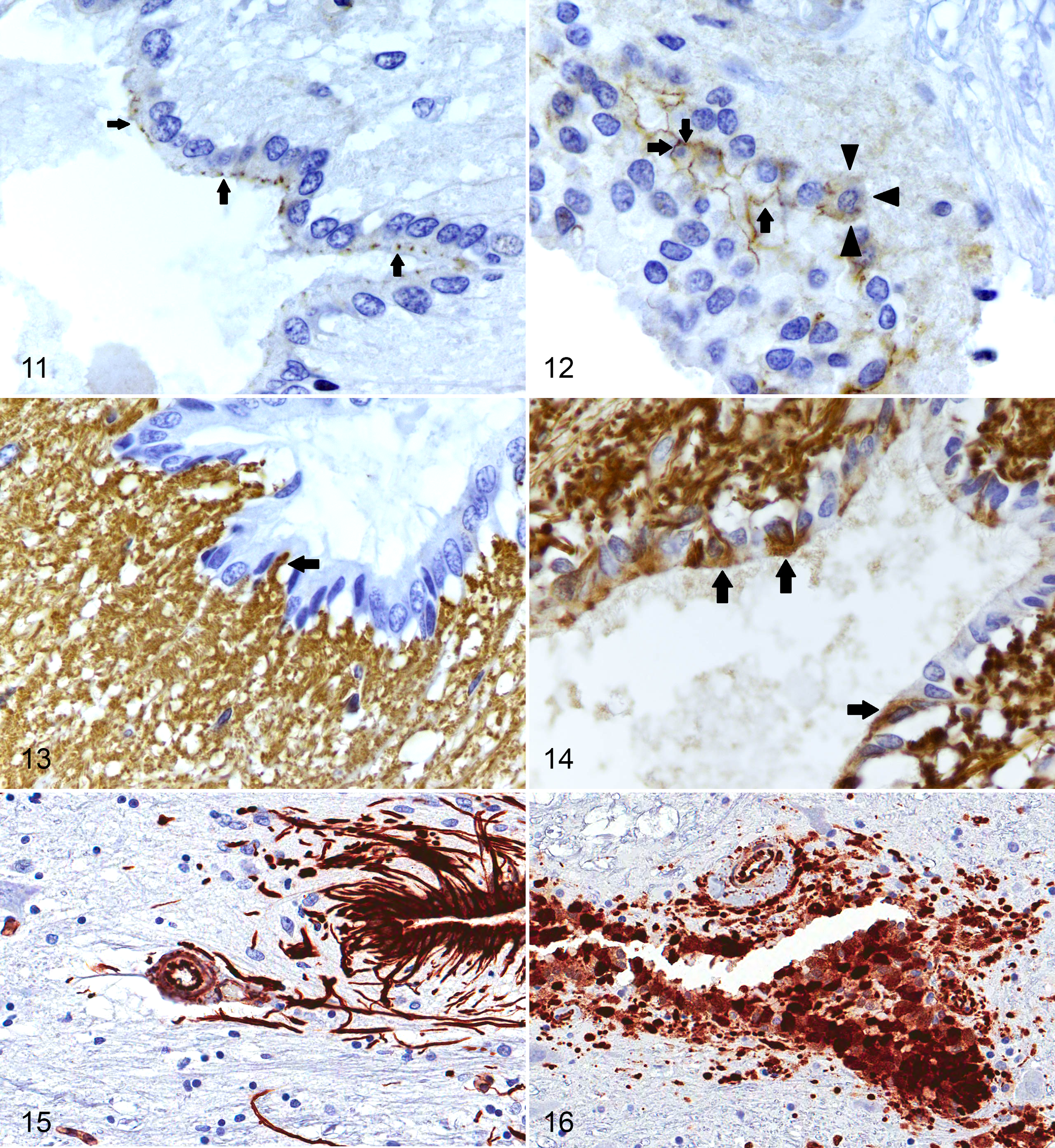
Normal, mid-thoracic spinal cord, dog. E-cadherin staining (arrows) in normal dogs is restricted to the apical surface of cells in the spinal ependymal layer. Immunohistochemistry for E-cadherin.
Mean positivity for GFAP in the SEL as measured by positive pixel count in normal dogs was 0.193 ± 0.05. There were no GFAP-positive cell bodies located in the SEL of normal dogs; however, GFAP-positive cell processes were occasionally observed within the SEL (Fig. 13). Mean GFAP positivity in the SEL of SCI-affected dogs was 0.540 ± 0.06 at the lesion epicenter and 0.470 ± 0.11 at proximal thoracic sites, and cytoplasmic GFAP staining was clearly noted in cell bodies located within the SEL (7 ± 3 positive cell bodies per transverse section at the lesion epicenter and 9 ± 2 positive cell bodies per transverse section at proximal sites) (Fig. 14). The finding of increased GFAP positivity in SCI-affected dogs was statistically significant at both the lesion epicenter and proximal sites (P = .001 and .006, respectively). GFAP positivity was inversely correlated with injury severity such that it was higher at the lesion epicenter in dogs with more mild injuries (r = –0.688; P = .04). There was also a significant positive correlation between GFAP staining and duration of injury, such that staining was higher in dogs with longer durations of injury (r = 0.841; P = .01).
Mean positivity of the SEL for vimentin, as measured by positive pixel count, was 0.907 ± 0.02 in normal dogs. Many cells of the normal SEL and their associated processes showed strong, uniform cytoplasmic staining for vimentin. Based on morphology, vimentin positivity was restricted to tanycytes and cuboidal ependymal cells, with CSF-contacting neurons appearing vimentin negative (Fig. 2). Vimentin-positive cell processes of SEL tanycytes extended well in to the gray matter and ventrally along the median fissure of the normal spinal cord. Occasionally, vimentin-positive cell processes were observed to envelope small vascular structures within the gray matter (Fig. 15). Mean positivity for vimentin in SCI-affected dogs was 0.889 ± 0.03 at the lesion epicenter and 0.941 ± 0.01 at proximal sites. There was not a significant difference in total vimentin positivity of the SEL in SCI-affected dogs compared with controls (P = .61 for epicenter and P = .20 for proximal sites), although vimentin-positive cell processes appeared subjectively blunted and disorganized in SCI-affected dogs compared with normal dogs (Fig. 16).
Discussion
To our knowledge, this is the first study to evaluate histologic and immunohistochemical properties of the normal canine SEL or to examine proliferative responses of the canine SEL in spontaneous SCI. Morphologically, the SEL in normal dogs was similar to other mammalian species such as the rat and human. 14,16,30 We observed several morphologically distinct populations of cells in the SEL, including tanycytes, CSF-contacting neurons, and cuboidal ependymal cells. We also documented several structural and functional changes in the SEL of SCI-affected dogs, not only at the lesion epicenter but also at distant sites within the spinal cord. These include both local and proximal alterations in expression patterns of markers of cellular proliferation, apoptosis, and cell lineage.
One of the most dramatic changes noted in the SEL of SCI-affected dogs was an approximately 3-fold increase in GFAP staining. In normal dogs, only occasional GFAP-positive cell processes could be noted in the SEL. This finding is expected since most tanycytes and cuboidal ependymal cells, the most prominent cell types in the SEL, are expected to be GFAP negative. 14 In normal dogs, we attribute the occasional GFAP-positive processes to astrocytes in the periependymal region, although future studies assessing additional markers of neural lineage will be required to confirm this. However, after SCI, GFAP-positive cell bodies were frequently noted in the SEL at both the lesion epicenter and proximal sites. This result mirrors what has been previously reported in rats, where this enhanced GFAP expression after injury is attributed to astrocytic differentiation of neural precursors located in the SEL. 47 Ependyma-derived astrocytes in rodents have been shown to migrate to sites of injury within the spinal cord to contribute to glial scar formation. 28,38 In addition, they exert regulatory influence within the ependymal cell niche and may initiate or maintain inflammatory responses in the cellular microenvironment after injury. 14,47 Our results would indicate that this occurs not just locally at the site of injury but more diffusely throughout the injured spinal cord and supports the concept of astrocytic differentiation of endogenous neural precursors in the canine SEL. As such, the dog model may lend itself well to evaluation of treatments that modulate this process after SCI. An alternative explanation for enhanced GFAP expression in the SEL after SCI may be astrocyte migration into the injured SEL.
We did not observe a statistically significant increase in total SEL cell count in SCI-affected dogs compared with controls. This finding was further supported by lack of expression of Ki-67 in the SEL, a highly specific marker of cellular proliferation. The unexpected paucity of Ki-67 expression observed in SCI-affected dogs in this study may reflect a lack of proliferative activity in the SEL at the time points examined and possible overall restricted proliferative capacity of the canine SEL; however, it is possible that proliferation within the SEL may occur either more acutely or more chronically after SCI than was examined here. In rodent models of SCI, ependymal cell proliferation as determined by bromodeoxyuridine labeling peaks between 3 and 7 days after injury. 33 The median duration of clinical signs for the dogs in this study was 72 hours; however, only 3 dogs fell into the 3- to 7-day window. In addition, a temporal profile of ependymal cell reactions after SCI has not been evaluated in the dog and may differ from that of rodents. While not statistically significant, there was a modest increase in SEL cellularity after SCI. This should be evaluated in a larger cohort of dogs and should include more systematic analysis of multiple spinal cord regions in future studies directed at examining the timing and proliferative capacity of the canine SEL after injury. An additional limitation to our assessment of SEL cellularity is that there are no SEL-specific IHC markers validated for use in dogs by which we could identify cells of interest. As such, our analysis of SEL numbers was restricted to identifying cells based on their morphology and position as part of the central canal, which can be broken or discontinuous after SCI. It is possible that additional cells of SEL origin were present within the parenchyma of the injured cord but were not identified during our analysis, leading to an underestimation of SEL cellularity after SCI.
We observed a paradox between PCNA and Ki-67 expression in the SEL. Specifically, PCNA expression was high despite an absence of Ki-67 staining in both normal and SCI-affected dogs. Baseline expression of PCNA in resting cells in the canine SEL is not surprising and in fact is consistent with what is reported in other species, including humans and rodents, where 30% to 80% of cells in the SEL may be strongly PCNA positive in the resting state. 10,47 This finding has been previously attributed to the low specificity of PCNA staining for cell proliferation. PCNA is generally regarded as a marker of cell proliferation expressed during G1- and S-phases of the cell cycle, where it functions as a sliding clamp for DNA polymerases. 26 However, PCNA also plays an important role in DNA repair and possibly in RNA transcription, such that expression may occur in the absence of cell proliferation. 40 More interesting was the observation that there was a significant decrease in PCNA expression in the SEL of SCI-affected dogs both at the lesion epicenter and at proximal spinal cord segments. PCNA expression in SCI-affected dogs positively correlated with the duration of clinical signs such that more acute injuries showed lower PCNA expression in the SEL, and this expression increased with duration of injury. In addition, dogs with more severe injuries had lower levels of PCNA expression than did dogs with mild injuries. A similar phenomenon has been reported in a rodent experimental model of SCI, where PCNA expression in the SEL was related to injury severity and fell to a minimum immediately after injury and then rebounded over time. 47 This finding may indicate ineffective or inhibited cellular repair processes in the SEL of more severely injured dogs acutely after injury and is of interest because it appears to be a global phenomenon in the SEL within the injured spinal cord. Because the median duration of injury was short in this study, we were unable to assess the effect of chronicity on PCNA expression; however, the importance of diffusely altered cellular repair mechanisms in the SEL after injury warrants further investigation.
There was not a significant difference in level of E-cadherin expression between normal and SCI-affected dogs; however, cells of the SEL in SCI-affected dogs displayed a unique cytosolic and circumferential orientation of E-cadherin expression. E-cadherin is an epithelial cellular adhesion protein that participates in proliferation, survival, and differentiation of neural stem cells through a niche-cell mechanism in the subependymal zones of the adult central nervous system. 18 It also plays an important role in epithelial differentiation during embryonic development and is a major component of epithelial cell-cell junctions. E-cadherin also regulates directional expansion of tissues such that orientation of E-cadherin–dependent adhesions between cells dictates the plane of cell division that occurs. 12,43 The switch from apical to cytosolic and circumferential membrane expression of E-cadherin after SCI may indicate a loss of cellular polarity in the SEL after injury and could serve to guide migration of cells from the SEL to injury sites within the spinal cord parenchyma.
Caspase 3–positive cells were observed in the SEL at the lesion epicenter after injury but not at proximal sites or in normal dogs, indicative of enhanced activation of apoptotic pathways in cells of the SEL at the site of injury. This finding is consistent with a previous report of apoptotic cells in the SEL at the injury site in a rodent model of SCI but not distant locations within the spinal cord. 47 Previous studies in rodents have correlated the number of apoptotic cells in the SEL with severity of injury; however, no such relationship was identified in the present study. Unfortunately, due to the small number of spontaneous clinical cases with tissue available for study, it was impossible to statistically compare results from groups of acute, subacute, and chronically affected dogs, making it difficult to draw reliable conclusions about this relationship.
In summary, the SEL of normal dogs has similar morphologic and immunohistochemical properties to that of humans and rodents. Unexpectedly, the proliferative capacity of the canine SEL appears restricted, at least acutely after SCI. Functional changes in the SEL after injury are consistent with what is reported in other species, including the potential for astrocytic differentiation of neural precursors within the SEL. These changes are noted both locally at the lesion epicenter after SCI and at distant sites throughout the spinal cord, indicating global changes to SEL function that may be harnessed for regenerative therapy or to modulate cellular responses to injury. Enhanced GFAP positivity and changes in E-cadherin expression patterns would support future studies evaluating the canine SEL as a source of endogenous neural precursors, and future directions should include the validation of more specific markers of neural stem cell lineage such as SOX2 for use in IHC studies in the dog. 24 Given our findings, dogs with spontaneous SCI present a valuable opportunity for continued investigation of the endogenous regenerative capacity of the SEL to improve recovery after SCI.
Footnotes
Acknowledgements
We thank Florinda Jaynes for her assistance with immunohistochemical staining, Shelly Haramia and Jenny Bolon for their assistance with slide digitization, and Tim Vojt for his assistance with figure preparation.
Author Contribution
Conception or design: SAM, MO. Data acquisition, analysis, or interpretation: SAM, MO. Drafting the manuscript: SAM. All authors participated in critically revising the manuscript, gave final approval, and agree to be accountable for all aspects of work to ensure integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this work was provided in part by a grant from NCI P30CA016058.