
Abstract
Inbred mice are a unique model system for studying aging because of the genetic homogeneity within inbred strains, the short life span of mice relative to humans, and the rich array of analytic tools that are available. A large-scale aging study was conducted on 28 inbred strains representing great genetic diversity to determine, via histopathology, the type and diversity of spontaneous diseases that aging mice develop. A total of 20 885 different diagnoses were made, with an average of 12 diagnoses per mouse in the study. Eighteen inbred strains have had their genomes sequenced, and many others have been partially sequenced to provide large repositories of data on genetic variation among the strains. This vast amount of genomic information can be utilized in genome-wide association studies to find candidate genes that are involved in the pathogenesis of spontaneous diseases. As an illustration, this article presents a genome-wide association study of the genetic associations of age-related intestinal amyloidosis, which implicated 3 candidate genes: translocating chain-associated membrane protein 1 (Tram1); splicing factor 3b, subunit 5 (Sf3b5); and syntaxin 11 (Stx11). Representative photomicrographs are available on the Mouse Tumor Biology Database and Pathbase to serve as a reference when evaluating inbred mice used in other genetic or experimental studies to rule out strain background lesions. Many of the age-related mouse diseases are similar, if not identical, to human diseases; therefore, the genetic discoveries have direct translational benefit.
Laboratory mice are important tools for studying complex genetics and disease mechanisms for almost all human diseases. 52 High-resolution genetic variation data for the commonly used inbred strains (http://www.sanger.ac.uk/sanger/Mouse_SnpViewer/rel-1410) as well as the ability to manipulate the genome, particularly with the new CRISPR/Cas9 technology, 15,21,22,88 make the mouse the tool of choice for a wide range of biomedical studies. Common causes of human morbidity and mortality are often associated with age-related degenerative or neoplastic diseases, and with an increasingly long-lived human (as well as domestic animal) population, the economic and social impacts of these diseases are ever increasing. 80 The relatively short life span of laboratory mice has enabled many studies of inbred strains and hybrid stocks to investigate aging. 34,38,91 However, most of these studies focused on the genetics of life span rather than the underlying diseases commonly associated with aging, which usually have a direct impact on life span. Such diseases of aging can have a cumulative effect, where the complications are associated either with many minor degenerative diseases that eventually overwhelm the body’s homeostatic mechanisms resulting in death or with catastrophic diseases, such as metastatic cancer, which ultimately kill the patient. Paradigms are changing from an aging research focused strictly on expanding life span to improving the quality of life independent of longevity that is the health span. The relationship of these events and the genetics underlying their onset can be dissected with these large inbred mouse population data sets.
Historically, research focused on lesions in aging mice in strains that were commonly used in biomedical research at the time. This provided background data intended as an aid for interpreting experimental results but also provided insight into the aging process itself as well as how inbred strains differed from one another. A limited number of inbred strains (BALB/cStCrlf, C3H/Nctr, DBA/2NNia), hybrid stocks (B6C3F1, C57BL/6N X C3H/HeN; B6D2F1, C57BL/6NNia X DBA/2NNia; or B6C3F1, C57BL/6NNia X C3H/NNia, in which females from the first strain are bred with males from the second strain) and a variety of other strains and stocks were used for various types of aging studies. 11,14,34,43 More recently, studies focused on general types of lesions in aging mice used in genetic engineering experiments, both inbred and those on segregating backgrounds. 90 Over time, some of the diseases change due to improvements in husbandry, pathogen status, and genetic quality control. Helicobacter hepaticus was considered part of the normal flora prior to 1994, but it is now recognized an opportunistic pathogen and has been eliminated from many research and vendor colonies. 84,85 Klebsiella oxytoca, another organism long ignored, is now considered an opportunistic pathogen, especially in strains that either have mutations in their toll-like receptor 4 (Tlr4) gene or are immunodeficient. 10 Mammary cancer was once common in many mouse strains. The Jackson Laboratory was founded to use mice affected with mammary cancer to discover the underlying cause, which proved to be the Bittner agent, later called the mouse mammary tumor virus. 18,71 Mouse mammary tumor virus has been eradicated from most vendor colonies, resulting in a decrease in mammary cancer, especially from C3H substrains. 51
Understanding the complex genetics of chronic diseases of aging in humans as a result of studies with mice will ultimately provide biomarkers that can be used to guide lifestyle changes or pharmacologic intervention. If done early, such modifications may prolong health span, improve end-of-life care, and ultimately reduce health care costs in humans.
We report here an overview of detailed histopathology-based investigations of 28 inbred strains of mice at various ages in cross-sectional and longitudinal studies. While >30 strains were initially used, most survived to the 12-month time point, but not all survived to the planned age for cross-sectional testing that is 20 months of age. Data were captured with the Mouse Disease Information System, 61,73 which provided a computable system for compiling and analyzing the subsequent massive data sets based on genome-wide association studies (GWASs). Representative photomicrographs from this and other studies can be accessed on public repositories (Mouse Tumor Biology Database, http://tumor.informatics.jax.org/ and Pathbase, http://www.pathbase.net). 3,31,58,59,61 These data provide a unique resource and background information that can be an aid to correct interpretation of lesions seen in old mice.
Materials and Methods
Mice
The following inbred strains, obtained from 1 vendor (The Jackson Laboratory, Bar Harbor, ME, USA), were selected for their interstrain genetic diversity: 53 129S1/SvImJ, A/J, AKR/J, BALB/cByJ, BTBRT +tf/J, BUB/BnJ, C3H/HeJ, C57BL/10J, C57BL/6J, C57BLKS/J, C57BR/cdJ, C57L/J, CAST/EiJ, CBA/J, DBA/2J, FVB/NJ, KK/HIJ, LP/J, MRL/MpJ, NOD.B10Sn-H2b/J (a congenic strain with the NOD genetic background but with a histocompatibility locus from a diabetes-resistant strain), NON/ShiLtJ, NZO/HlLtJ, NZW/LacJ, P/J, PL/J, PWD/PhJ, RIIIS/J, SJL/J, SM/J, SWR/J, and WSB/J. Three of the strains were not evaluated because of lymphoma development (AKR/J) confirming a well-documented strain-specific phenotype, 68,69 lack of sufficient mice for analysis (CAST/EiJ), or self-mutilations resulting in euthanasia for humane purposes (SJL/J). Therefore, GWAS analyses were performed in 28 strains of mice.
Mice were maintained in a humidity-, temperature-, and light cycle– (12:12 hours) controlled vivarium under specific pathogen–free conditions (http://jaxmice.jax.org/genetichealth/index.html). Mice were housed in double-pen polycarbonate cages (330-cm2 floor area), with 4 mice per pen. Mice were allowed free access to autoclaved food (NIH 31, 6% fat; LabDiet 5K52, Purina Mills, St Louis, MO, USA) and acidified water (pH 2.8–3.2).
All work was done with the approval of The Jackson Laboratory Animal Care and Use Committee (approval No. 06005). Mouse handling and care were followed according to the Public Health Service animal welfare policies.
Aging Colonies
The study was divided into 2 parts: (1) a cross-sectional study to evaluate onset and diversity of diseases at 12 and 20 months of age; (2) a longitudinal, or life span, investigation, to determine how long each strain survived. Mice in the longitudinal study were maintained until they became moribund, at which point they were euthanized by CO2 asphyxiation.
Mice in the cross-sectional study were euthanized in groups by strain during a 4-month period at 12 months ± 28 days and 20 months ± 28 days of age. Blood was collected at the time of necropsy from the submandibular vein for hematology (Siemens Advia 2120 Hematology Analyzer, Siemens Healthcare Diagnostics, Newark, DE, USA) and blood chemistry (Beckman Coulter 600 Pro Chemistry Analyzer, Beckman Coulter Corp, Brea, CA, USA). Just prior to necropsy, body mass was determined with a PIXImus machine (Lunar PIXImus, GE Medical Systems, Hermosa Beach, CA, USA). All mice were subjected to a complete necropsy. 66 All physiologic data, including clinical pathology data, are available on the Mouse Phenome Database (http://phenome.jax.org/).
The Cross-sectional Study
Two cohorts were maintained from all the strains described above—each consisting of 15 females and 15 males and each with a designated survival end date: 12 months and 20 months. Mice that survived to their respective time points were euthanized by CO2 asphyxiation, and a complete necropsy was performed. 66
The Longitudinal Life Span Study
A cohort of 32 females and 32 males for each of the 28 strains was initially set up. A second cohort of 32 females was subsequently added, totaling 96 mice/strain, which was calculated to provide enough statistical power to detect a 10% difference in life span. 91 Complete necropsies 66 were done on moribund mice but not on mice found dead, as autolysis precluded a meaningful interpretation.
Tissue Collection
Tissues were collected in a systematic manner and fixed by immersion in Fekete’s acid alcohol formalin solution. 66 In our experience, Fekete’s acid alcohol formalin solution provides the optimal fixation for histology and especially for antigen preservation for immunohistochemistry of mouse tissues. 40 After overnight fixation, tissues were transferred to 70% ethanol and later trimmed, processed routinely, embedded in paraffin, sectioned at 6 μm, and stained with hematoxylin and eosin. Pancreata were fixed attached to the intestines. Additional samples of pancreata were removed without any attached tissues other than mesentery, fixed in Bouin’s solution, washed in running water overnight, processed as above, but stained with aldehyde fuchsin to evaluate pancreatic islets for insulin production. Skulls and the vertebral column were also fixed in Bouin’s solution, allowed to fix and decalcify for a week, and then processed as above to generate hematoxylin and eosin–stained slides. The neurologic tissues were evaluated separately and are not reported here. 74 Additional special stains—such as toluidine blue for mast cell tumors, phosphotungstic acid hematoxylin for rhabdomyosarcomas, or alizarin red or von Kossa for mineralization—were used as needed to aid in reaching a definitive diagnosis. More extensive workups were done on specific diseases, which will be or were reported separately. Twenty-four slides were generated per mouse, with an average of about 3 tissues per slide.
These included “Swiss Rolls”
42
of the duodenum, jejunum, ileum, and colon (with anus and perineal skin) Longitudinal section of the stomach with esophagus and cecum (inflated with fixative) Cross sections of the left lateral and medial lobes of liver (to include the gall bladder), spleen, right kidney with adrenal glands, the left kidney was trimmed lengthwise so that it could be differentiated from the right kidney, the reproductive organs (testis, epididymis, accessory sex organs for males; ovary, uterine tube, uterus, mammary glands for females), the preputial gland for males / the clitoral gland for females, the salivary gland cluster with cervical lymph nodes, the heart, the esophagus and trachea with thyroid and parathyroid glands, and the tongue Longitudinal sections of the center of the lobes of both lungs, dorsal skin, ear skin (pinna), ventral skin, muzzle skin, eyelid, and left kidney Longitudinal section of hind leg, including stifle/knee joint Longitudinal section of front leg, including shoulder and elbow joints Longitudinal section of hind foot (soft tissues, bone, and nail unit/footpad) Longitudinal section of front foot (soft tissues, bone, and nail unit/footpad) Longitudinal and cross sections of the lumbar spine Longitudinal and cross sections of the tail Longitudinal and cross sections of the skull Sections of the lower jaw to include teeth
9
Bones fixed in Fekete’s acid alcohol formalin solution were processed in Cal-Ex (Fisher, Pittsburgh, PA, USA). Bones fixed in Bouin’s solution had no additional decalcification done.
Data Management
Diagnoses were entered into an electronic medical record system consisting of a relational database and a user interface for the pathologist to seamlessly enter specific findings for each mouse. This database, the Mouse Disease Information System, 73,78 uses the Mouse Anatomy Ontology to code for the organ 25 and the Mouse Pathology Ontology 61 to code for the disease process (diagnosis). A severity score was recorded (0, normal; 1, mild; 2, moderate; 3, severe; or 4, extreme) to account for differences in the size, severity, and biological behavior of lesions.
GWASs and Variant Identification
Phenotype-genotype association analysis was performed via the expedited efficient mixed-model association algorithm. 29 Frequencies and mean severities of the selected disease phenotypes in 28 strains of mice were used as input phenotype data. Four million single-nucleotide polymorphisms (SNPs), which are freely available at http://mouse.cs.ucla.edu/mousehapmap/full.html, were used as input genotype data. Each SNP was evaluated individually and recorded as –log(P values) as the strength of the genotype-phenotype associations in Manhattan plots. SNPs were annotated with the Variant Effect Predictor by Ensembl (http://useast.ensembl.org/info/docs/variation/vep/index.html). Coding region SNPs known to change the amino acid sequence of the protein (ie, nonsynonymous SNPs) were tested for functional importance via the PolyPhen-2 algorithm (http://genetics.bwh.harvard.edu/pph2/). 1 Identified genes were evaluated by extensive literature searches and evaluations of protein-protein interactions and potential biological pathways.
Results
Numbers of Mice Examined
While most inbred mouse strains survived to 12 months of age, it became rapidly apparent with the longitudinal study that few mice would reach 24 months of age, the initial goal for the older cohort. As such, the second group was only aged to 20 months of age. There was wide variation in the numbers of individual mice that reached 20 months of age, which was reported previously. 74 All AKR/J mice died after the 12 months of age exclusively due to development of lymphomas, a known strain-specific lesion due to an endogenous retrovirus. 68,69 All AKR/J mice had structural abnormalities of their hair shafts, unrelated to age, due to a mutation in the Soat1 gene. 23,86 SJL/J fought extensively, especially but not exclusively the males, requiring many to be euthanized. As a result, no SJL/J mice were available for necropsy at 20 months of age. Other diseases, such as alopecia areata, were found in old mice in strains previously identified to develop the disease. 39,75
A total of 1742 full necropsies were performed. Others were found dead and not necropsied due to advanced autolysis. A total of 20 885 different diagnoses were made, with an average of 12 diagnoses per mouse in the study. A summary of the most common lesions identified by strain, excluding neurologic lesions, is shown in Tables 1 and 2. Detailed summaries of all lesions or specific cancer types by strain are published elsewhere. 2,74
Condensed Summary of Diseases in 20-Month-Old Female Mice Representing 28 Inbred Strains of Mice.a


aFrequently diagnosed diseases in almost all mice of a strain are in darker gray tones. Intermediate grey tones indicate that about half the mice were affected. Light gray tones indicate that at least 1 mouse was affected in the group. Data are shown only for lesions that affected at least 1 strain at an intermediate level. Neurologic diseases are not reported here.
 very common diagnosis;
very common diagnosis;  moderately common diagnosis;
moderately common diagnosis;  occasional diagnosis.
occasional diagnosis.
Condensed Summary of Diseases in 20-Month-Old Male Mice Representing 28 Inbred Strains of Mice.a

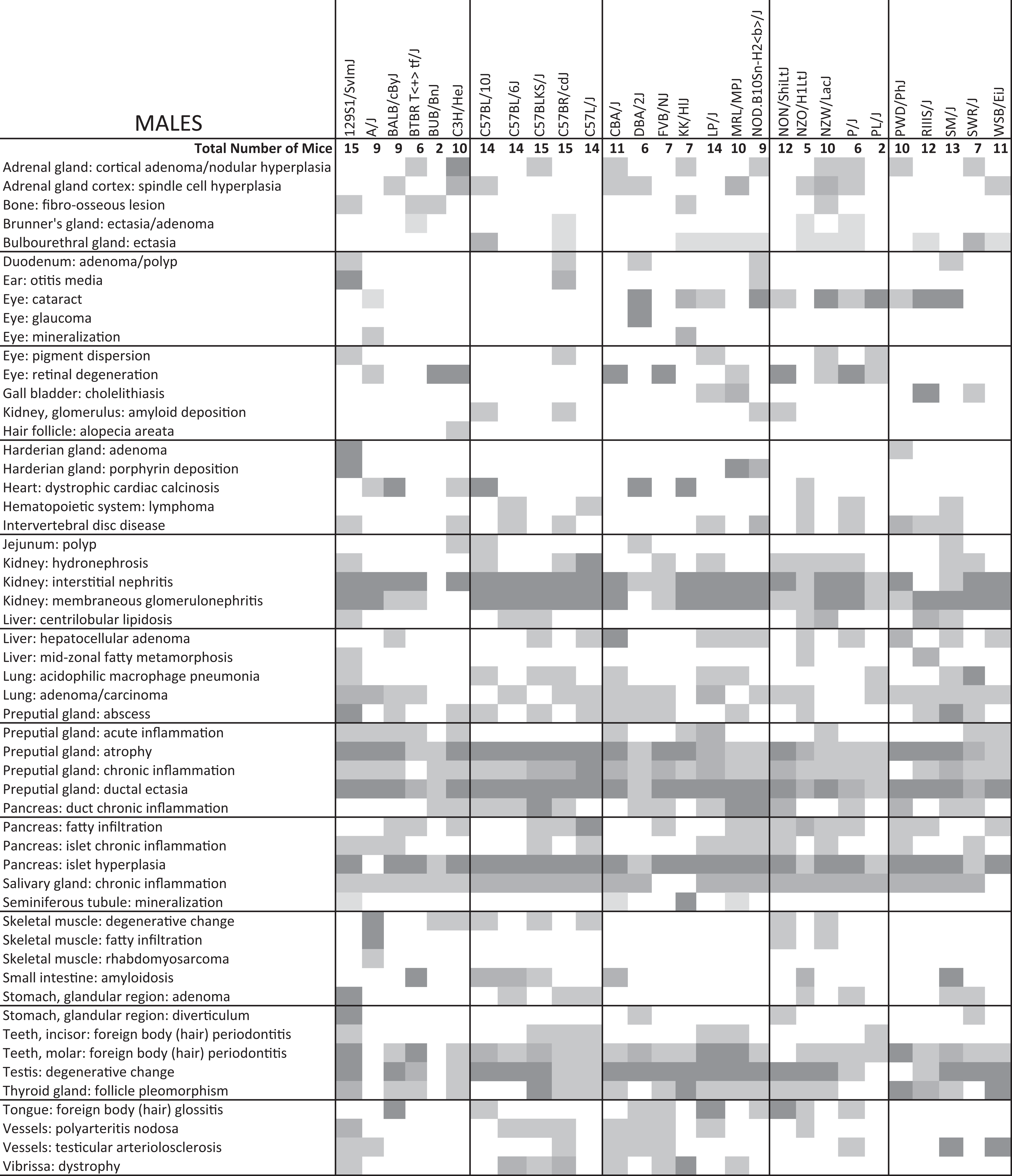
aFrequently diagnosed diseases in almost all mice of a strain are in darker gray tones. Intermediate gray tones indicate that about half the mice were affected. Light gray tones indicate that at least 1 mouse was affected in the group. Data are shown only for lesions that affected at least 1 strain at an intermediate level. Neurologic diseases are not reported here.
very common diagnosis; moderately common diagnosis; occasional diagnosis.
Diseases and Causes of Death
The mice selected at 12 and 20 months of age were, for the most part, in reasonably good health and were euthanized by CO2 asphyxiation. However, detailed histologic analyses of all organ systems revealed a wide variety of chronic diseases, the most frequent of which are listed in Tables 1 and 2. These tables compress variations in disease presentations (eg, dystrophic cardiac calcinosis, which includes epicardial and myocardial mineralization and fibrosis [Berndt A, Li Q, Sundberg BA, et al, “Mouse Genome-wide Association Study Identifies Polymorphisms on Chromosomes 4, 11 and 15 for Age-Related Cardiac Fibrosis,” unpublished data]) that are defined in greater detail in other focused studies. Neurologic lesions were reviewed by others and are not reported here.
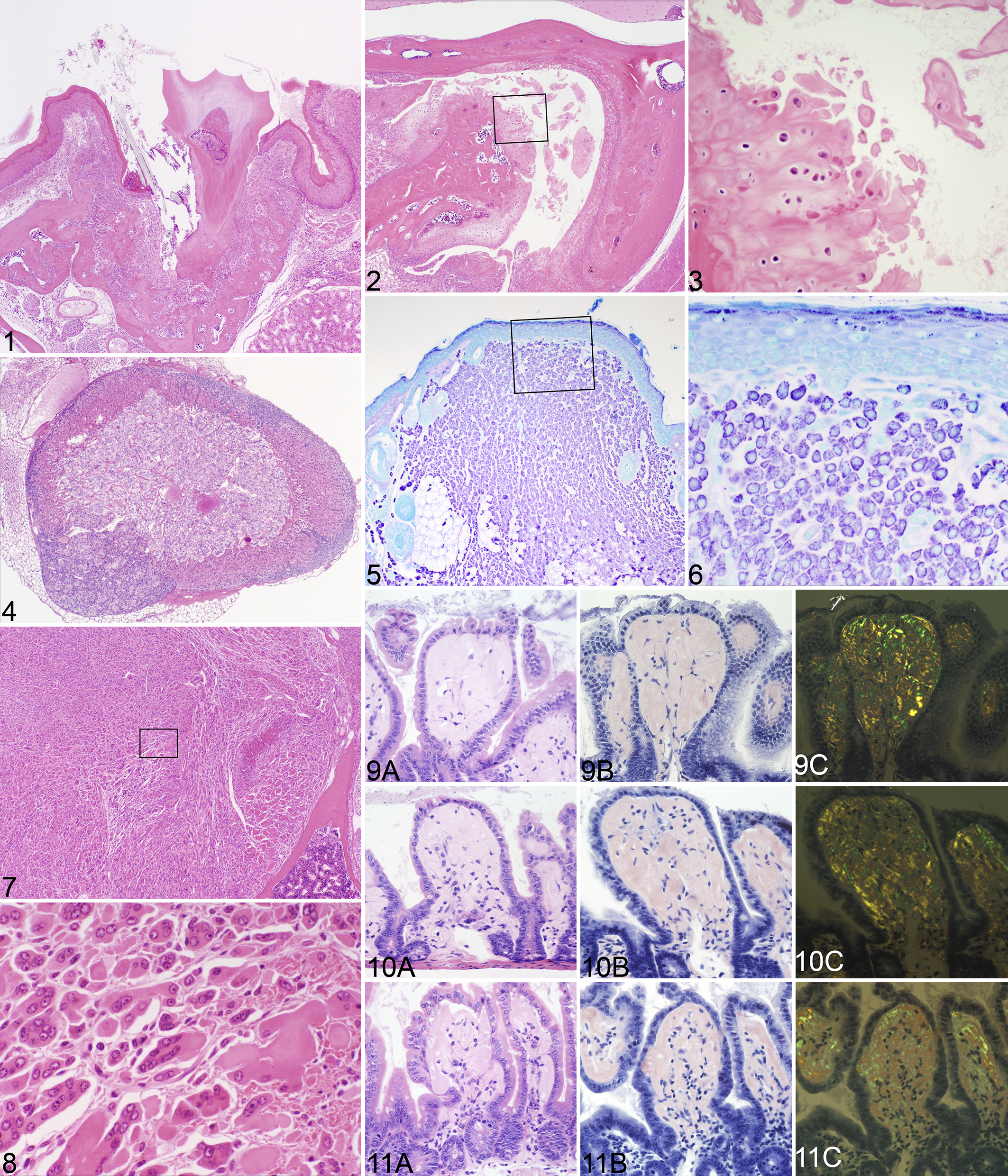
Some lesions were found in all or almost all mice regardless of strain or sex, such as oral foreign body impactions (hair impaction secondary to grooming and food material impacted around teeth or in the lingual gland duct, causing acute to chronic inflammation and, in some cases, avulsion of teeth; Fig. 1), 36 degenerative changes of the preputial and clitoral glands (ductal ectasia, sebocyte atrophy, and various degrees of acute to chronic inflammation or abscessation), 63 temporomandibular joint disease in very old mice in the longitudinal study (Figs. 2, 3), or pancreatic islet hyperplasia. 9 Other diseases, especially cancer, were for the most part not consistent in any one strain, with the following exceptions (Figs. 4–8): lymphomas in AKR/J, 68,69 adrenal cortical adenomas in C3H/HeJ (Fig. 4), 26 and rhabdomyosarcomas in very old A/J mice (Figs. 7, 8). 65 Mast cell tumors are extremely rare spontaneous findings in inbred mice, limited in this study to single cases in FVB/NJ (Figs. 5, 6) and NZW/LacJ mice. Detailed summaries for cancer diagnoses by strain are reported elsewhere. 2 Eye lesions have been well described in laboratory mice, and these were similar in the aged mice. 67
GWAS Analysis
Not surprising, because of how inbred strains were developed, 53 groups of related strains often had similar or identical diseases. Due to the large amount of data collected, it is not possible to include detailed analyses on all diseases in an overview of this kind; therefore, individual workups integrating GWAS analyses with detailed phenotyping are being reported elsewhere (Berndt A, Li Q, Sundberg BA, et al, “Mouse Genome-wide Association Study,” unpublished data). 5 –9 To illustrate how these computational methods can be applied to aging disease data sets, amyloidosis of the small intestine, well known in some C57BL/6 substrains, 37,55 is used as an example. Amyloidosis of the small intestine was found in about half of the strains in this study (Figs. 9–12), not just C57BL/6J. Genome-wide scans were performed with the expedited efficient mixed-model association algorithm based on a 4 million–SNP panel from the National Institute of Environmental Health Sciences. Twenty-month-old mice, females and males, were evaluated and scored for small intestinal amyloidosis in 28 inbred strains. 74 Frequency of intestinal amyloidosis lesions was most strongly associated (P < 10−9) with genome variations on chromosomes 1 and 10: Chr 1 at 13.549419 Mb (MM38-1-13549419:NES15822440; P = 4.86·10−10) and Chr 10 at 12.92 Mb (NES11975634; P = 4.86325E-10), 12.95 Mb (NES11974532; P = 4.86325E-10), 13.023 Mb (NES11972003; P = 4.86325E-10), and 13.024 Mb (NES11971958; P = 4.86325E-10), which contained 3 candidate genes: Tram1, Stx11, and Sf3b5 (Table 3).
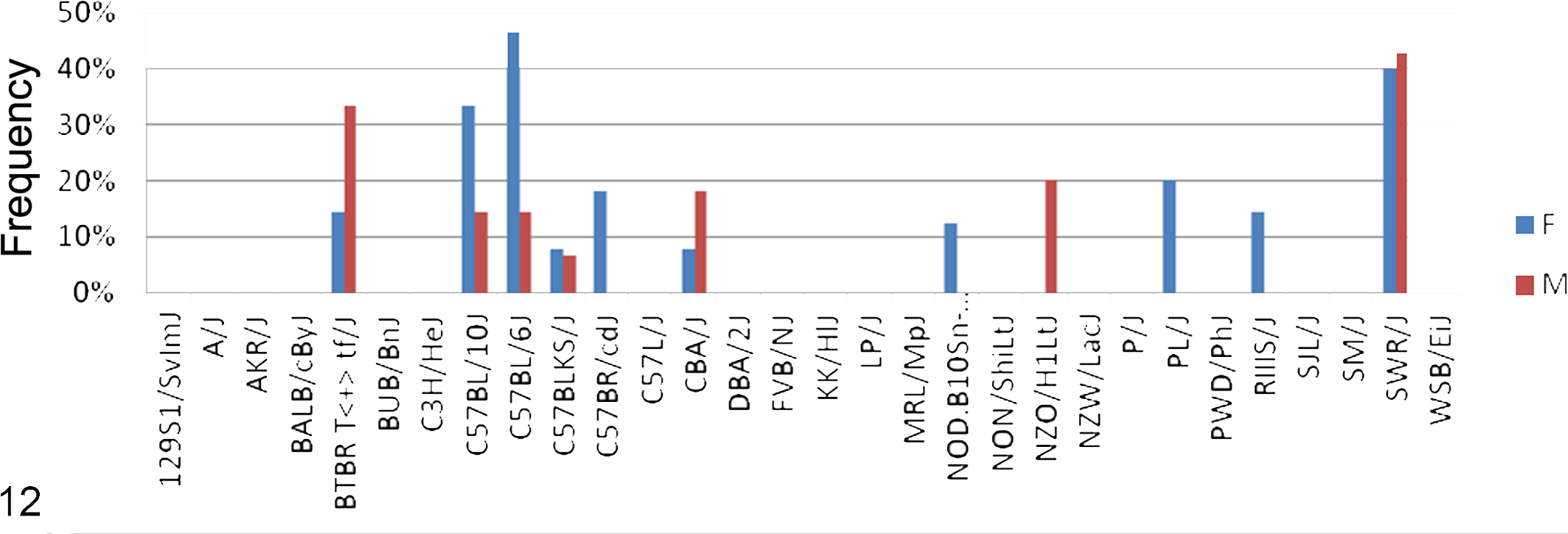
Frequency (percentage of affected mice by strain) of amyloidosis of the small intestine in 28 inbred strains of mice at 20 months of age. F, female; M, male. Note that the C57 strains and substrains are commonly affected.
Candidate Genes for Amyloidosis of the Small Intestine as Identified by Genome-wide Association Studies of 28 Aged Inbred Mouse Strains.a
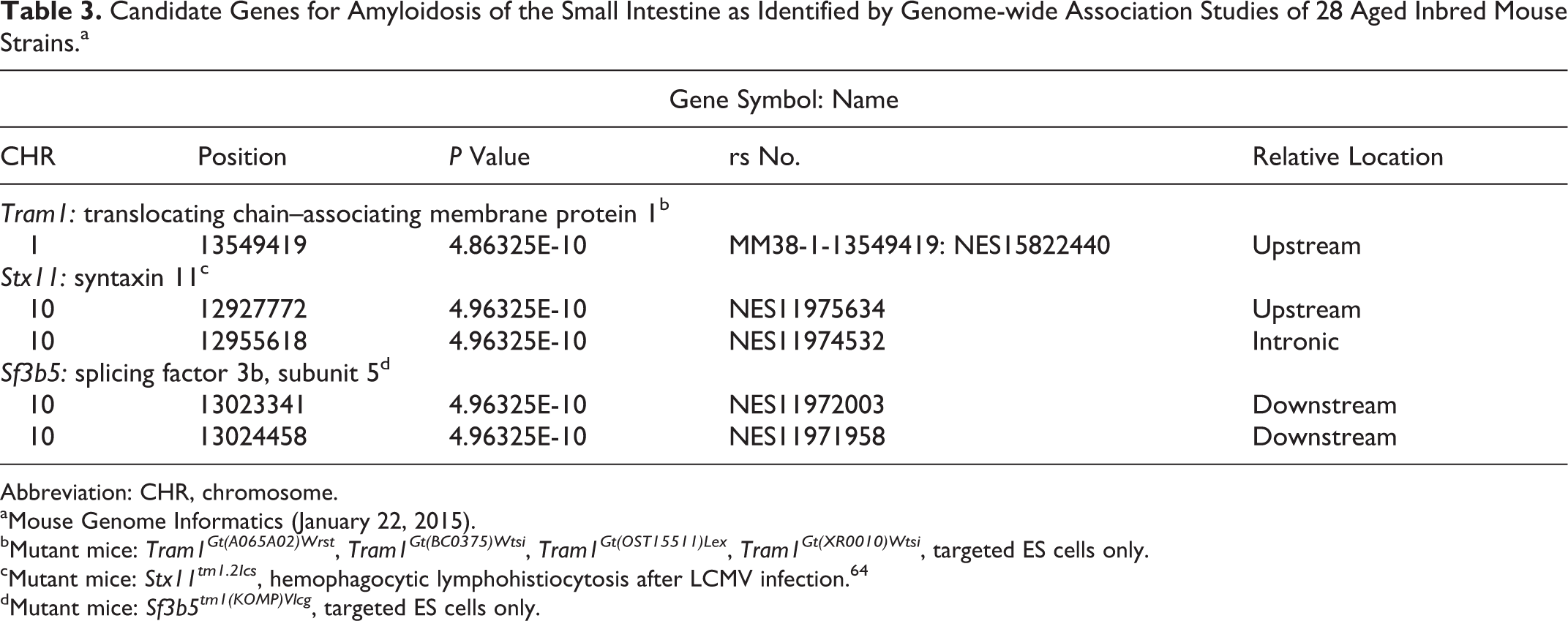
Abbreviation: CHR, chromosome.
aMouse Genome Informatics (January 22, 2015).
bMutant mice: Tram1Gt(A065A02)Wrst , Tram1Gt(BC0375)Wtsi , Tram1Gt(OST15511)Lex , Tram1Gt(XR0010)Wtsi , targeted ES cells only.
cMutant mice: Stx11tm1.2Ics , hemophagocytic lymphohistiocytosis after LCMV infection. 64
dMutant mice: Sf3b5tm1(KOMP)Vlcg , targeted ES cells only.
Discussion
While the data gathered here are enormously valuable for understanding the types and diversity of diseases found in commonly used inbred mouse strains, one of the most valuable outcomes was an understanding of how to set up these types of histopathology-based studies of aging. Debates continue regarding whether (1) to focus on moribund mice to establish a mean life span with detailed histopathology near the time of death or (2) to collect samples from mice at specific ages or time points. Based on experience doing large-scale disease surveillance based on limited, lesion-specific diagnostic workups rather than complete necropsies, disease trends can be found in young mice up to 8 months of age. 76 However, for very detailed multiorgan histologic studies, only small numbers of old mice are practical due to animal maintenance costs and the perceived high cost of histopathology. 79 Diseases were identified in 12-month-old mice that were also found in 20-month-old group but less frequently. Some diseases became obvious only at a very old age. For example, rhabdomyosarcomas changed from a rare to a common disease in A/J mice >20 months of age. 65 By contrast, AKR/J mice are highly prone to lymphomas and rarely survive to 20 months of age, and those that do have few, if any, other lesions. 68,69 Evaluating mice at standardized ages does provide a cross-sectional profile of all strains at a particular age; however, it is not the best way to determine what diseases are life-threatening and potentially the ultimate cause of death in any particular strain. Aging mice, until they become moribund, provide a higher chance of discovering strain-specific disease predisposition and correlating these diseases to mean life span. However, many of these mice die before they can be necropsied, making generation of population data more difficult. Regardless, both approaches yield valuable information.
Another concern is whether histopathology is a useful, cost-effective tool for evaluating phenotypes or if physiologic (in vivo) phenotyping 60 (ie, panels of functional diagnostic tests) provide an adequate and more cost-effective means of evaluating mouse health. 41 Histopathology, when done by experienced personnel, has for >150 years been the stand-alone standard for arriving at a diagnosis. By contrast, physiologic phenotyping approaches reflect human clinical practice and, in the context of model organisms, are often applied later in hypothesis-driven approaches to understanding phenotypes as confirmatory assays selected on the basis of histopathology findings. A problem with physiologic assays is that they provide data out of context. For example, high red blood cell counts were observed in some members of a cohort of a single strain (SM/J females; Table 1). While statistical analyses would suggest that this might be an error in the hematologic assay and exclude the data, the histologic diagnosis that affected mice had an erythropoietic neoplasm revealed the real significance of these data. Histopathology in most cases can be used independently to arrive at a definitive answer, often with direct correlation with a human disease.
The value of these histopathologic data sets is dependent on the skill of the pathologist or investigator evaluating the study. For example, teratomas 19,20 and sebaceous gland tumors 44,92 reported in aged mutant mice were actually misdiagnoses of common, nonneoplastic, normal “aging changes” of the preputial and clitoral glands (Tables 1 and 2), organs found in mice but not humans. 63 Hair shaft impaction around the incisors and especially the molars, causing foreign body periodontitis, was supposedly associated with ulcerative dermatitis in C57BL/6 strains and hybrids 16 but is found in almost all mice regardless of their hair coat or cutaneous ulcers 36,77 and was commonly diagnosed in C57BL/6J and many other strains in this aging study in mice with no concurrent alopecia or other skin condition. Many other examples are abundant in the literature. 28
While the interest of pathologists is and should be in the variety of types of lesions that old mice develop, especially in strains beyond those commonly used, it is fascinating to see that groups of inbred strains, sometimes closely related but not necessarily, have essentially the same diseases while others do not. As an example, amyloidosis of the small intestine is a well-described finding in aging mice, particularly in some of the C57BL/6 substrains. 37,55 This diagnosis was confirmed in the study reported here. By treating this as a binary trait—that is, either a strain had the disease or it did not—GWAS analyses identified 3 candidate genes for amyloidosis of the intestine (Tram1, Sf3b5, and Stx11).
Only 1 of the 3 candidate genes identified has a known role in inflammation or amyloidosis (Stx11), and this is discussed below. An approach to further establishing whether genes associated with particular phenotypes may have a hitherto unknown role is to consider known protein-protein interactions and network membership—the so-called guilt-by-association approach. 47 Based on the modularity of disease phenotypes, 50 this approach has now become widely used. Using protein-protein and genetic interaction data, we explored the immediate interactome surrounding our candidate genes and report significant molecular or regulatory associations that might explain their genetic association with amyloidosis. 93
Tram1 presents an interesting candidate for involvement in amyloidosis and amyloid-accumulating diseases such as Alzheimer disease. So far Tram1 has not been implicated genetically in amyloid-associated degenerative disease; however, a search of BioGRID 13 for interacting partners presents an abundance of evidence for its potential involvement (Figure 13, Suppl. Table 1). TRAM1 is an integral transmembrane glycoprotein of 36 kDa, which forms part of the translocon in the endoplasmic reticulum, is involved in membrane insertion of nascent proteins, and has a role in the disposal of misfolded proteins in the endoplasmic reticulum. 45 This is a mechanism common to amyloid accumulation in many tissues irrespective of the protein involved. TRAM1 interacts directly with amyloid precursor protein (APP), 45 as shown in an interactome screen in Alzheimer disease, as well as with the SEC61P complex. It has been shown to bind SEC61B by direct crosslinking. 30 SEC61B has been shown to be a critical protein in the cytoplasmic export of amyloid beta. 30 In Alzheimer disease and other amyloidosis syndromes, the misfolded peptide accumulates both in extracellular plaques and in the cytosol. Exported proteins are normally degraded in the cytosol by protease-dependent and protease-independent pathways. Other interacting partners of TRAM1, such as BCAP31 and DERL1, are also involved in protein translocation and endoplasmic reticulum processing and degradation, which is a precursor for amyloid accumulation. 27,49,62,82,83
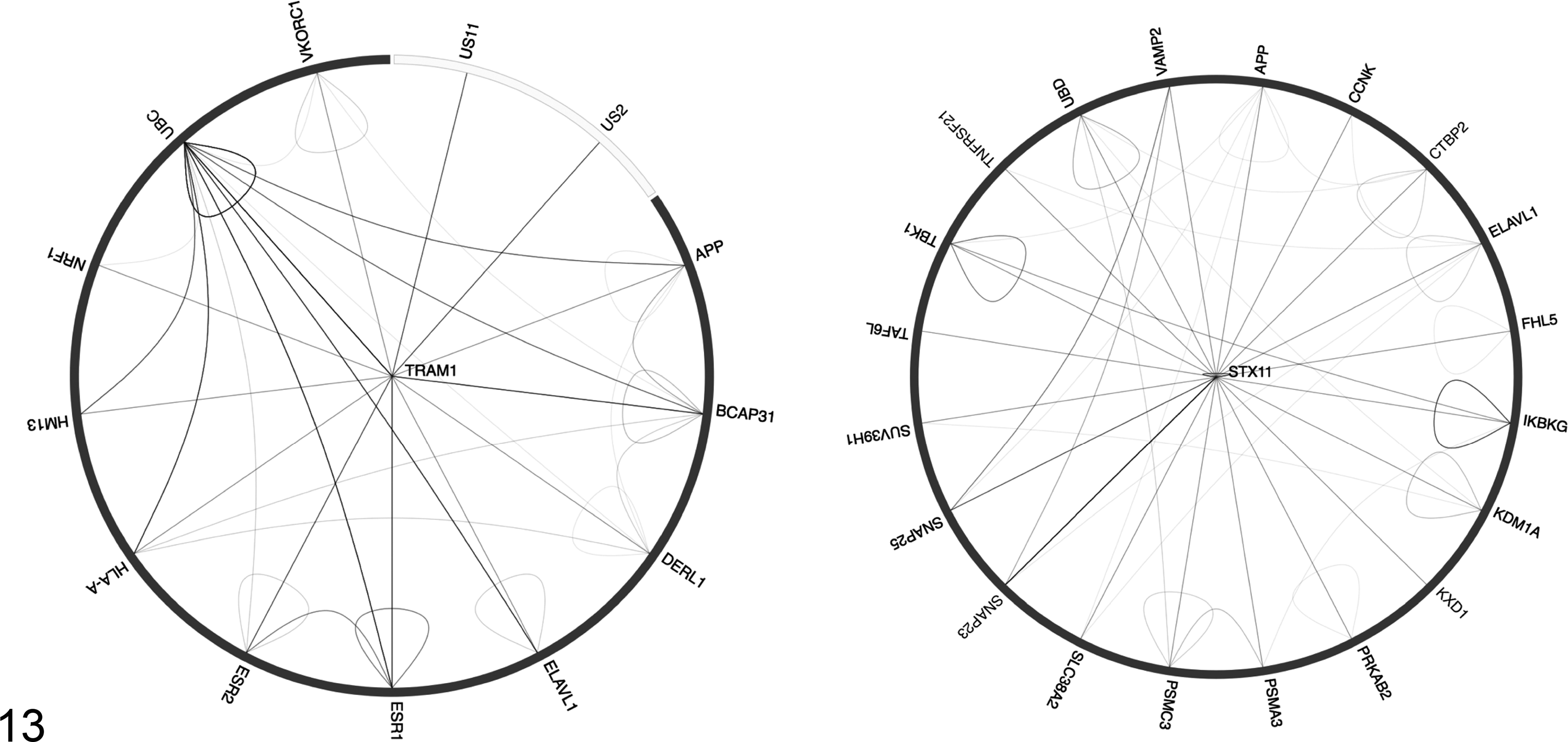
Interaction diagrams from BioGRID for STX11 and TRAM1. BioGRID 3.3.123 searched 44 772 publications for 823 466 raw protein and genetic interactions from major model organism species. The diagram shows physical interactions as edges connecting gene products (ie, protein-protein interactions) for all of the interspecies interactions, self-interaction, and interactions between interactors. The raw data and evidence are shown in Supplemental Tables 1 and 2.
TRAM1 also interacts with HM13. 49 Previously known as presenilin-like protein 3, HM13 is a GxGD class of proteinase of the same family as the presenilins and acts as a transmembrane ion channel. 32 The presenilins are responsible for cleavage of the cleavage of APP and are implicated in Alzheimer disease, 87 although direct involvement of HM13 in other amyloid accumulation diseases has not yet been shown.
TRAM1 functions as a cofactor to promote efficient cytomegalovirus US2- and US11-dependent dislocation of major histocompatibility complex class I heavy chains and seems to hijack the translocation complex, also involving DERL1. 49 Recently, a suggestive link has been made between Alzheimer disease and cytomegalovirus susceptibility. 35 It may be interesting to pursue the relationship between cytomegalovirus infection and apparently sporadic amyloidosis, with the hypothesis that subvention of the TRAM1-associated translocation complex may compete for translocation of other proteins, such as amyloid beta retrotranslocated protein.
Finally, the direct interaction with VKORC1, the vitamin K epoxide reductase complex subunit 1, 57 is intriguing. While VKORC1 does have direct interaction with TRAM1, APLP2, and BCAP31, the gene has never been directly linked to amyloid accumulation, being a primary determinant of blood-clotting efficiency and sensitivity to warfarin. 54 However, the Vkorc1tm1.1Jobg knockout mouse shows phenotypes consistent with cerebral amyloid angiopathy as well as the expected clotting defects. 70
In humans, SF3B5 interacts with survival motor neuron domain containing 1 (SMNDC1), which is genetically associated with Crohn disease and therefore may be involved in the site specificity of intestinal amyloidosis. 89 Targeted embryonic stem cells have been made for Sf3b5; however, live mice have not yet been produced. It is possible that they will be created as part of the International Mouse Phenotyping Consortium on the C57BL/6N background, but even in those studies, mice will be aged only to 15 weeks, too early to detect any effect on intestinal amyloidosis.
Stx11 is a member of the soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) family, which function as executors of exocytosis during CTL cell killing and are important for the fusion of lytic granules at the immunologic synapse. 24 In humans, a deficiency in syntaxin 11 (STX11) causes familial hemophagocytic lymphohistiocytosis type 4 (FHL4). 12,94 Genetically engineered Stx11tm1.2Ics null mice on a C57BL/6 background had a normal immune system when compared with wild-type controls with no evidence of amyloidosis, but this was not the focus of the study. Disease similar to FHL4 in humans could be reproduced in these mutant mice by infection with lymphocytic choriomeningitis virus. 64
A search of BioGRID (Fig. 13, Suppl. Table 2) revealed that the immediate interactome of STX11 contains several genes of potential interest, including amyloid beta A4 protein. 46 The interactions with Vamp2 (synaptobrevin), Snap23 (synaptosomal-associated protein 23), and Psma3 (Proteasome subunit alpha type-3), a component of the proteasome, reflect the involvement of STX11 in vesicle transport. 72 However, their interactions are modified by amyloid resulting in modulation of neurotransmitter release. 56
Interestingly STX11 interacts with TNFRSF21, 81 a member of the TNF-receptor superfamily also called death receptor 6 (DR6), which has an important role in inflammation and both B- and T-cell function and proliferation 4 and functions with APP in axonal branching control. 48 Two allelic mutations exist in mice (Stx11tm1.2Ics and Stx11tm1Bupa , as well as several ES cell lines, MGI:2151075). These have been mainly studied from the standpoint of immune and vascular function or neurobiology, so it is not clear whether extra–CNS amyloidosis would have been detected and reported.
Overall, the associations that we have found between TRAM1 and STX11 and their interacting partners suggest possible novel pathogenetic roles for these proteins based on their activity as part of larger complexes and networks. That there may be several predisposing factors to the pathogenesis of intestinal amyloidosis is consistent with what we know about its pathobiology, and the analysis offered here provides new candidates for further mechanistic investigation.
This study demonstrated how the combination of defined and extensive genetic variation in inbred strains, with detailed phenotyping, can generate hypotheses about the genetic basis for relative predisposition to the phenotypes or diseases of interest. The genes that we have identified therefore provide hypotheses about the underlying pathogenetic process. Association does not necessarily imply causation, and as with all GWAS analyses, hypothesis-driven experimentation is now needed to support or reject the genetically derived hypothesis. The causative genes associated with identified SNPs are not necessarily the closest, and it is very rare that the SNPs themselves are the cause of the altered gene function. 17 Use of informatics, annotations, and extensive literature searches can provide clues regarding which associations reflect true involvement, 33 but we are far from having enough phenotype and genotype data for this to be useful in all cases. Until we have all of the systematic phenotyping from the International Mouse Phenotyping Consortium completed, the gaps and biases in data will render any prioritization approach suboptimal but valuable. Finally, using whole genome variant analysis instead of imputed SNP sets may eventually provide targeted identification of true genomic variants.
Footnotes
Acknowledgement
P.N.S. thanks the Warden, Fellows, and Scholars of Robinson College, Cambridge, for travel support.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: JPS, KAS, and VEK have a research contract with Bioniz, LLC, and a research grant from NAAF for AA preclinical trials. All other authors state that they have no conflicts of interest.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Ellison Medical Foundation, Parker B. Francis Foundation, National Institutes of Health (AG25707, for the Shock Aging Center; CA89713, for the Mouse Tumor Biology Database). The Jackson Laboratory Shared Scientific Services were supported in part by a Basic Cancer Center Core Grant from the National Cancer Institute (CA34196, to The Jackson Laboratory).