
Abstract
The authors documented gross and microscopic morphology of lesions in corals on 7 islands spanning western, southern, and eastern Micronesia, sampling 76 colonies comprising 30 species of corals among 18 genera, with Acropora, Porites, and Montipora dominating. Tissue loss comprised the majority of gross lesions sampled (41%), followed by discoloration (30%) and growth anomaly (29%). Of 31 cases of tissue loss, most lesions were subacute (48%), followed by acute and chronic (26% each). Of 23 samples with discoloration, most were dark discoloration (40%), with bleaching and other discoloration each constituting 30%. Of 22 growth anomalies, umbonate growth anomalies composed half, with exophytic, nodular, and rugose growth anomalies composing the remainder. On histopathology, for 9 cases of dark discoloration, fungal infections predominated (77%); for 7 bleached corals, depletion of zooxanthellae from the gastrodermis made up a majority of microscopic diagnoses (57%); and for growth anomalies other than umbonate, hyperplasia of the basal body wall was the most common microscopic finding (63%). For the remainder of the gross lesions, no single microscopic finding constituted >50% of the total. Host response varied with the agent present on histology. Fragmentation of tissues was most often associated with algae (60%), whereas necrosis dominated (53%) for fungi. Two newly documented potentially symbiotic tissue-associated metazoans were seen in Porites and Montipora. Findings of multiple potential etiologies for a given gross lesion highlight the importance of incorporating histopathology in coral disease surveys. This study also expands the range of corals infected with cell-associated microbial aggregates.
Keywords
Coral reefs face a variety of threats globally, 9 including overfishing, 15 terrestrial pollution, 10 global climate change, ocean acidification, 22 and disease. Coral cover in the western Atlantic has declined almost 80% in the last 30 years, 11 leading to major shifts in reef structure and species composition in the region 4 ; diseases, particularly those that cause tissue loss, are suspected to have played an important role in this decline. 6,8,26 Diseases in corals are also being documented more frequently in the Pacific, 35 with recent examples including tissue loss diseases in Acropora sp from the Great Barrier Reef and the Marshall Islands, 31 Montipora sp from Hawaii, 2 and Porites from the Philippines. 28
Lesions such as tissue loss, growth anomalies, or discoloration in corals are manifestations of disease. 39 Systematic descriptions of lesions at the gross and microscopic levels provide the foundational information for the case definition of animal diseases and provide a deductive process to assign potential causation to particular diseases. 24,44 However, coral disease investigations have not traditionally followed a deductive approach and often lack descriptions of lesions at the microscopic level. 36 This limits the types of information available to interpret gross lesions and can hamper the understanding of coral disease. Now recognizing the utility of histopathology, recent studies are starting to include histology in their methods. 30
Field surveys are a staple of coral disease investigations and provide important demographic data on status and trends of various types of lesions in corals in the Atlantic 34 and Pacific. 2 To understand changes at the cellular level, combining field surveys with histopathology of gross lesions provides a robust method for disease investigations in corals. However, our knowledge of coral disease in the Pacific, particularly in remote atolls and regions of the South Pacific, is rudimentary to nonexistent. Only 1 coral disease survey exists for Micronesia, and it is limited to the island of Guam. 20 As part of an interdisciplinary survey of coral reefs in the Federated States of Micronesia, our objective was to systematically characterize lesions encountered in corals in the region at the gross and microscopic levels. We report here on a variety of host responses and potential etiologic agents of disease in multiple species of corals, including the discovery of heretofore undescribed potential symbionts.
Methods
We surveyed corals for lesions in 7 islands in Federated States of Micronesia spanning the entire archipelago (Fig. 1). Two 25-m transects were laid end to end, separated by about 10 to 15 m and parallel to the reef crest at depths of 6 to 10 m, and corals were surveyed for lesions within a 6-m wide swath along the transect. All gross lesions on the transect were photographed and classified into 3 broad categories, including tissue loss, discoloration, and growth anomaly. Tissue loss was subdivided as follows: acute, where a distinct margin of tissues was apposed to bare white skeleton; subacute, where a distinct margin of tissues was apposed to a variably sized band of bare white skeleton that became progressively overgown by turf algae, with increasing distance from the tissue margin; and chronic, where tissues were apposed to skeleton completely overgrown by epibiota. 39 Discoloration was classified as bleaching, dark, or other discoloration. Bleaching comprised tissues that were white, and it was subdivided as diffuse or localized. Dark discoloration comprised variably sized distinct irregular dark brown to black areas. Other discoloration included all other forms of abnormal tissue pigmentation. Growth anomalies were categorized as umbonate, exophytic, rugose, or nodular. 41
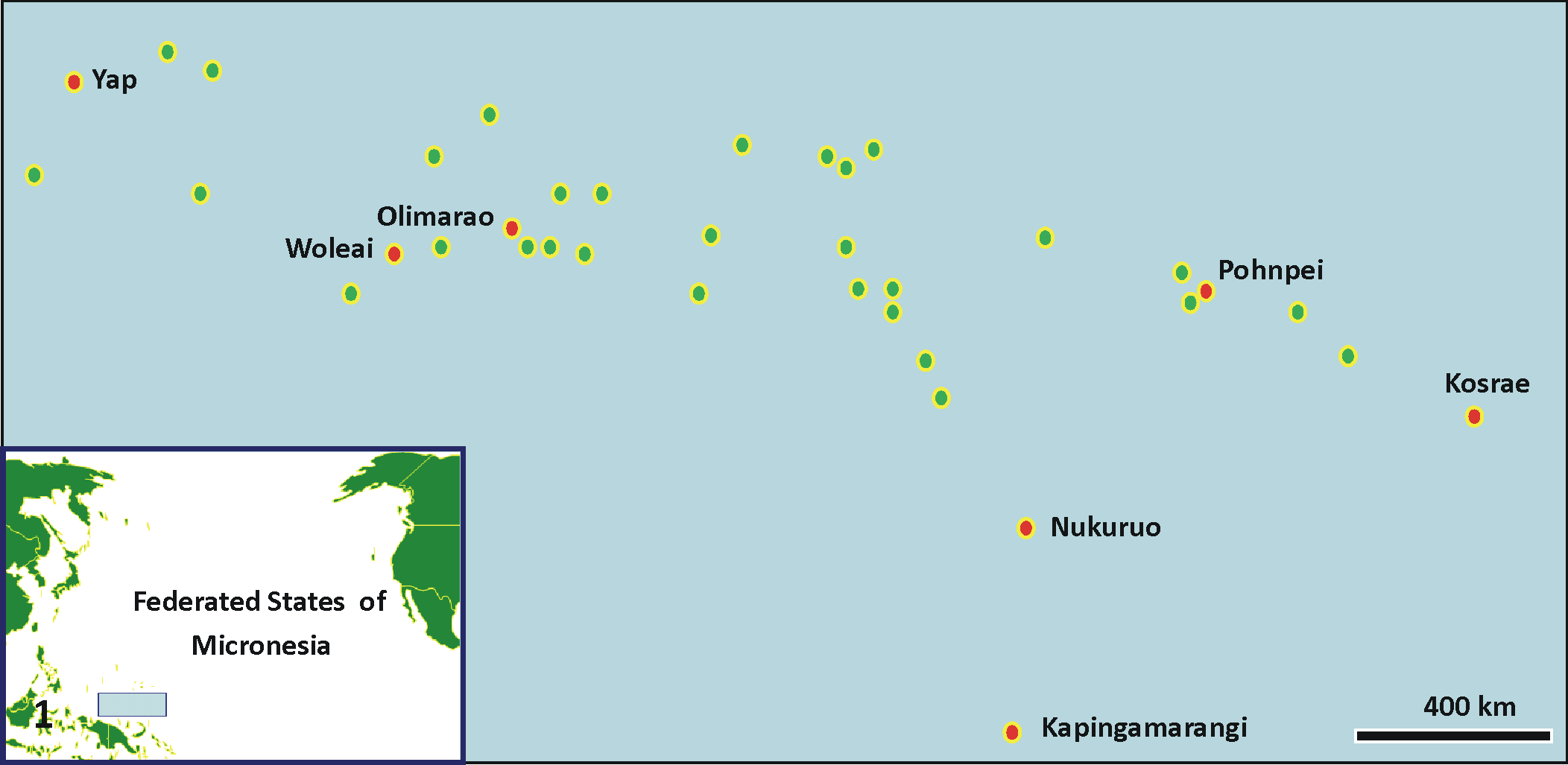
Map of the islands of Federated States of Micronesia (circles), located north of Papua New Guinea (see blue rectangle in inset map of Pacific). Red circles are islands where surveys were done.
Coral fragments (2–5 g) were collected with chisel or bone shears and placed into individually numbered Whirl-Pak bags in seawater. Fragments with lesions were collected ensuring inclusion of the border between normal and lesional tissues. When available, paired grossly normal fragments were also collected. Fragments were fixed in Z-Fix (Anatech Ltd) diluted 1:5 with seawater and decalcified in dilute formic acid/formaldehyde solution (Cal-Ex II, Fischer Scientific). Tissues were then embedded in paraffin, sectioned at 5 µm, and stained with hematoxylin and eosin. 38 To confirm presence of fungi or bacteria, Grocotts methenamine silver or Gram stains were used, respectively. 27
Microscopic changes were broadly categorized by agent associated with cell pathology, if present, and host response. Skeletal spaces were differentiated from gastrovascular canals based on cells lining the space; spaces lined by calicodermal cells were classified as decalcified skeleton, whereas those lined by gastrodermis or containing mesenterial filaments were classified as gastrovascular canals. Agents were identified according to their microscopic morphology and included sponges or cnidarian, 13 helminths, 14 algae, 18 fungi, 17 or crustacean. 29 Sponges consisted of metazoa with a matrix containing spicules, choanocytes, and presence or absence of zooxanthellae; cnidaria were metazoa with nematocysts; helminths were vermiform metazoa with or without a gut; algae were metazoa with cell walls; fungi were elongate branching filamentous structures with or without septa; and crustacea were metazoa with gut, muscle, reserve inclusion cells, cuticle, hepatopancreas, and segmented appendage. Host response or changes included tissue fragmentation, suspect wound repair, hyperplasia of basal body wall, necrosis, or inflammation. 38,40,41,45 Tissue fragmentation comprised variably sized clumps of intact cells near the main intact tissue section; suspect wound repair comprised regeneration of epidermis on exposed mesoglea of basal body wall or epidermal metaplasia of exposed calicodermis; hyperplasia of basal body wall comprised widespread proliferation of gastrodermis, mesoglea, and calicodermis of basal body wall with reduced formation or absence of mesenterial filaments and lack of polyp structures such as actinopharynx or tentacles; necrosis comprised cells manifesting cytoplasmic hypereosinophilia or shrinkage associated with pyknosis or karyorrhexis; inflammation comprised infiltrates of tissues with larger-than-normal amounts of mesogleal cells. Microscopic changes were not mutually exclusive, and in such cases, if an agent was associated with a lesion, it took priority when categories were assigned. If a host response was associated with only the lesion, the most severe change took priority during category assignment. Finally, we noted the presence/absence of symbiotic cell-associated microbial aggregates in the upper or basal body wall. 37 These are characterized by variably sized cell-associated clusters of symbiotic gram-negative bacteria in the gastrodermis or epidermis with no associated host response.
To aid in the interpretation of lesions, a series of supplemental figures are available highlighting diverse aspects and variations of normal microscopic anatomy of corals (see Supplemental Figs. S1–S12).
Results
We sampled a total of 76 colonies from 7 islands—including 17 colonies each from Kapingamarangi and Kosrae, 16 from Woleai, 12 from Pohnpei, 9 from Yap, 3 from Nukuoro, and 2 from Olimarao—comprising 30 species of corals among 18 genera, with Acropora, Porites, and Montipora dominating. Of 76 colonies examined, tissue loss constituted the majority of lesions that we encountered (41%), followed by discoloration (30%) and growth anomaly (29%). Of 31 cases of tissue loss, most lesion samples were subacute (48%; Fig. 2), followed by acute (Fig. 3) and chronic (Fig. 4; 26% each). Of 23 samples with discoloration, 40% were dark discoloration (Figs. 5, 6), with bleaching (Fig. 7) and other discoloration (Fig. 8, 9) each comprising 30%. Of 22 growth anomalies, umbonate (Figs. 10, 11) composed half of those seen in 10 species of 6 genera. In addition, exophytic growth anomalies (Fig. 12) were seen in 1 each of Acropora cerealis and Acropora surculosa; nodular growth anomalies, in 3 Lobophytum crassus and 2 Montipora grisea; and rugose growth anomalies, in 1 each of M. grisea, Montipora sp, Porites evermanni, and Hydnophora microconos (Supplemental Table 1).

Subacute tissue loss, Galaxea fascicularis from Kosrae. Note band of bare white skeleton bereft of tissues (black arrow) progressing to green algal turf cover (white arrow).

Multifocal pink discoloration, Porites sp from Kapingamarangi. Algae and inflammation were seen on microscopy.
For 9 cases of dark discoloration, fungal infections predominated (7 of 9, 77%); for 7 bleached corals, depletion of zooxanthellae from the gastrodermis made up a majority of microscopic diagnoses (4 of 7, 57%); and for growth anomalies other than umbonate, hyperplasia of the basal body wall was the most common microscopic finding (7 of 11, 63%). For the remainder of the gross lesions, no single microscopic finding composed >50% of the total.
Host response varied when agent was considered a primary microscopic diagnosis. For 10 cases with algae as the primary agent (Figs. 13, 14), the dominant host response was fragmentation (6 of 10 cases, 60%), whereas necrosis was the dominant host response associated with cases having fungi as the primary agent (10 of 19, 52%; Figs. 15, 16). Crustacea (Fig. 17) and bivalves (Fig. 18) did not manifest an evident host response, and the single case of helminth infestation was associated with fragmentation.
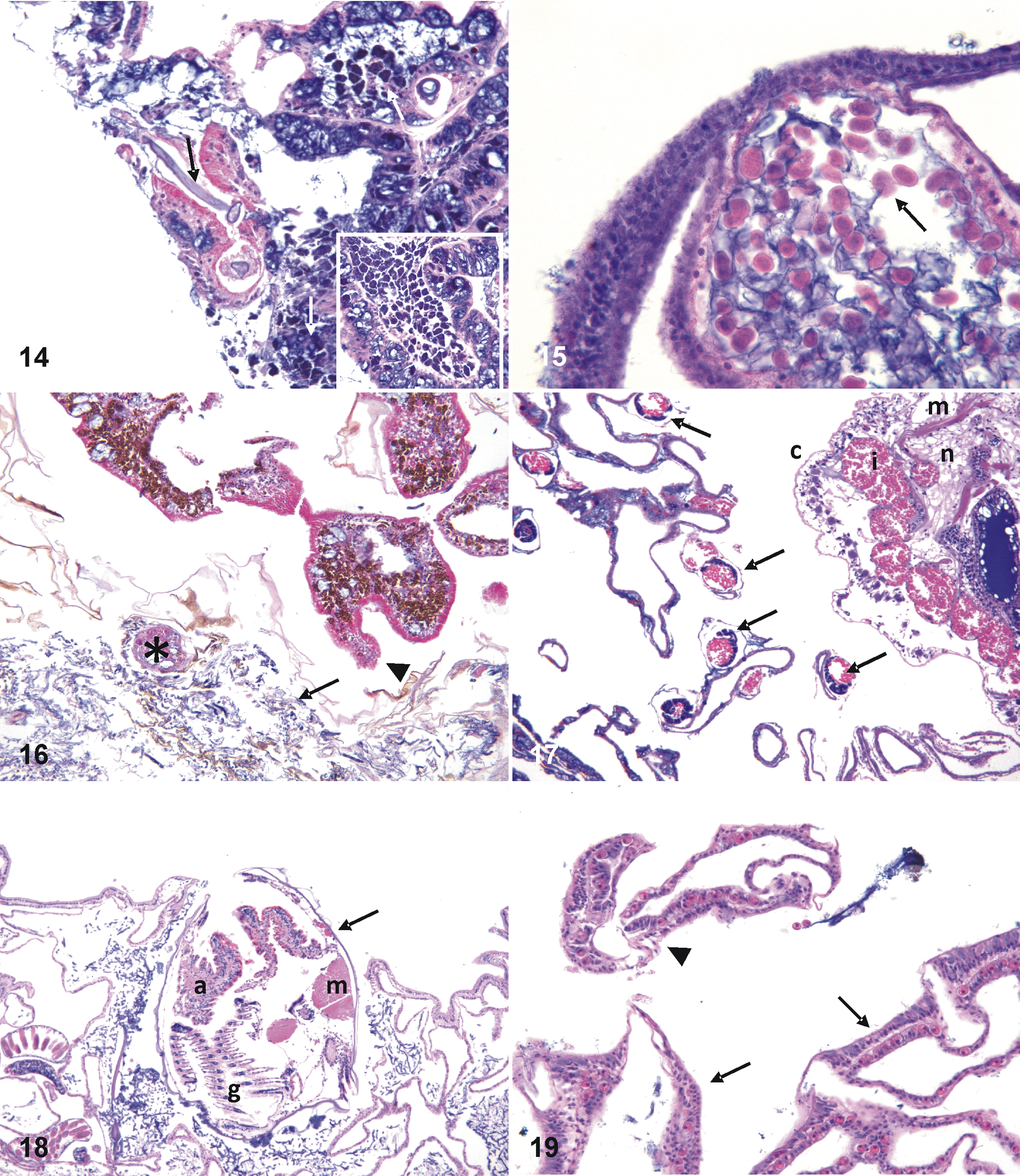
Multifocal pink discoloration, Porites sp (Fig. 8). Note filaments of algae (black arrow) surrounded by hypereosinophilic fragmented tissues and marked adjacent infiltrates of granular brown cells (white arrow). Inset: higher magnification of granular brown cells that, on other studies,
21
have stained positive with melanin. Hematoxylin and eosin (HE).
For primary diagnoses comprising host responses, there was variation in associated microscopic lesions. Of 11 cases of fragmentation, wound repair was the most common associated finding (6 of 11, 54%; Fig. 19). In cases of depletion of zooxanthellae, atrophy was the most common associated finding (4 of 6, 66%; Fig. 20). Most cases of necrosis (7 of 8, 87%; Fig. 21) had no associated lesions, and 7 of 10 (70%) cases of hyperplasia of the basal body wall (Fig. 22) had no associated microscopic findings. Hyperplasia of the epidermis was seen in only a single umbonate growth anomaly (Fig. 23). Of 12 corals manifesting microscopic evidence of inflammation, Porites (5 of 12, 41%) and Montipora (4 of 12, 33%) composed the majority of genera, with Isopora, Lobophytum and Acropora having 1 instance each.
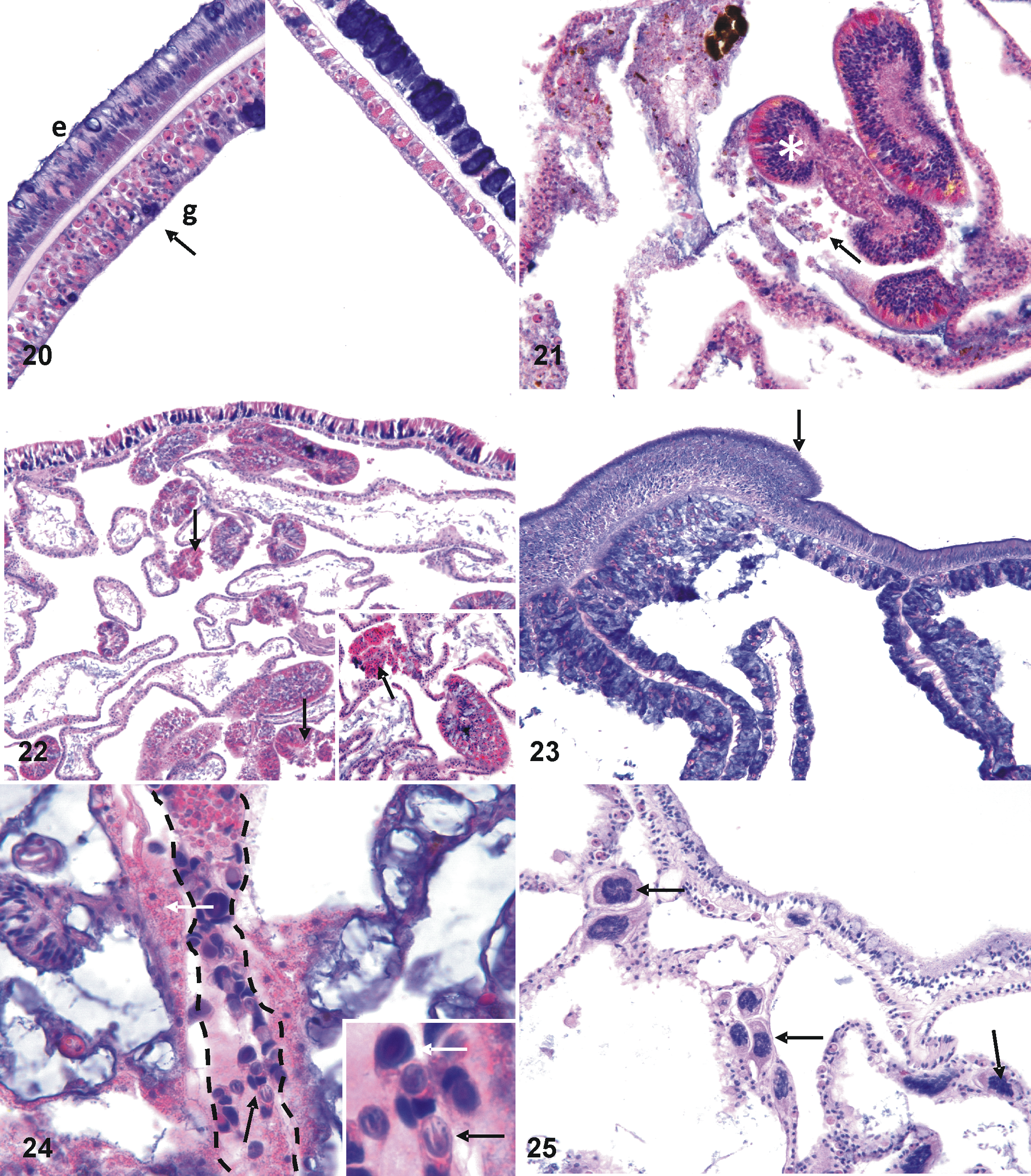
Bleaching, Mycedium robokaki (Fig. 7). Left panel is normal upper body wall with tall columnar epithelium (e) with plump gastrodermis (g) replete with zooxanthellae (arrow); contrast with right panel, showing depleted zooxanthellae in thin gastrodermis and thin epidermis where cells have mostly atrophied and are effaced by basophilic mucocytes.
Two metazoans closely associated with coral tissues elicited no host response in both normal and diseased tissues. One metazoan was elongate with numerous nematoblasts and nematocytes compatible in morphology with microcnidiaria (Fig. 24). These were found in 2 Porites sps from Pohnpei, 1 M. grisea from Kapingamarangi, and 1 Porites rus from Kosrae. Another apparent symbiont comprised unidentified multicellular (2–4 nuclei) organisms within the mesoglea of a single M. grisea from Kapingamarangi (Fig. 25).
Of 67 paired normal tissues, 27, 22, and 18 originated from colonies with tissue loss, growth anomalies, and discoloration, respectively. Seventy-one percent of paired normal fragments had no microscopic lesions, whereas lesions were seen for 7 of 27 (26%) paired normal fragments for tissue loss, 8 of 22 (36%) for growth anomaly, and 4 of 18 (22%) for discoloration. For the 7 paired normal fragments from tissue loss colonies, 3 had fragmentation, and 1 each had fungi, algae, sponges, or inflammation. Of the 8 paired grossly normal fragments from growth anomaly colonies, 5 had fungi, and 3 had necrosis. Of the 3 paired normal fragments from discolored colonies, 2 had fungi, and 1 had inflammation.
Cell-associated microbial aggregates were seen in corals from Woleai, Kosrae, Kapingamarangi, Nukuruo, and Pohnpei, with A. cerealis, A. hyacinthus, A. surculosa, Galaxea fascicularis, Hydnophora exesa, Platygyra daedala, Porites cylindrica, and P. evermanni infected.
Discussion
The findings in this study reinforce a concept that distinct gross lesions in corals have multiple potential etiologies and host responses, thus highlighting the utility of histopathology in coral disease surveillance. In this study, the most common agents associated with tissue loss were algae and fungi, and host response was most commonly fragmentation and necrosis. In other studies of tissue loss in Acropora from the Pacific, algae, fungi, helminths, and sponges were associated with various host responses, including wound repair, fragmentation, and necrosis. 38 A longitudinal study of tissue loss in Montipora capitata in Kaneohe Bay Hawaii 45 revealed ciliates commonly associated with rapidly progressing tissue loss (acute tissue loss) and helminths or chimeric parasites 43 associated with slowly progressing tissue loss (subacute tissue loss). Thus, a pattern continues to be confirmed with tissue loss diseases in corals in the Indo-Pacific that are associated with a variety of hosts responses and potential etiologic agents, even when the condition is examined for a single species and a single location.
Discoloration included bleaching, dark discoloration, pink multifocal discoloration in Porites, and discoloration secondary to burrowing molluscs. As per other instances of bleaching in corals, 7 the typical histologic finding for this lesion was depletion of zooxanthellae often associated with atrophy of tissues. Prolonged bleaching leads to loss of tissue biomass in other coral species. 32 Dark discoloration was associated with fungal infections in 80% of the corals. Fungi were associated with foci of necrosis and typically invaded the lower and upper skeleton, often with formation of structures compatible in morphology with fruiting bodies near the surface body wall. These findings are consistent to those found in Montipora and Pavona from Hawaii and American Samoa, where corals with dark discoloration manifested endolithic hypermycosis. 42 Hence, this represents a range extension of this condition to Micronesia. Future studies for this disease should focus on identifying the fungi associated with these lesions and the drivers that promote overgrowth of skeleton. The presence of fruiting bodies only near the epidermis was an interesting phenomenon. Given that light does not influence distribution of endolithic fungi, 12 other drivers must be influencing fruiting body production near the upper body wall in corals, and these merit further exploration. Other discolorations included multifocal pink discoloration in Porites associated with algal infiltrations and multifocal dark pinpoint cavities in Montipora sp. that appeared as little dark spots associated with endolithic bivalves. Other endolithic organisms, such as barnacles, associated with multifocal punctate cavities have been documented in a variety of corals in the Pacific and Atlantic. 3 Our findings emphasize that pink spots in corals are not invariably associated with trematode larvae, 1 as pointed out elsewhere. 5 Porites trematodiasis has a very distinct “pink spot” that is greatly swollen and never has pinpoint cavities, sediment, or algae observed on the swelling. Careful observation of pink spots in the field, followed by histologic examination, is required to discriminate trematodiasis from other causes.
Growth anomalies were common lesions in corals throughout Micronesia. Like growth anomalies in Acropora elsewhere in the Pacific, 41 growth anomalies in Acropora from Micronesia manifested as hyperplasia of the basal body wall occasionally associated with necrosis. A similar finding predominated in nodular and rugose growth anomalies in other species of corals, and others have found hyperplasia of basal body wall in the Caribbean. 25 However, umbonate growth anomalies seen in a variety of species here had a greater range of microscopic manifestations above and beyond hyperplasia of the basal body wall, including epidermal hypertrophy, bleaching, fragmentation, fungal infections, and endolithic crustacea. The crustacea were seen exclusively in Turbinaria, and their morphology with a series of rootlets distributed locally within the skeleton is consistent with rhizocephalans, parasitic barnacles often found in crabs that also grow rootlets throughout their host. 33 The coral skeleton surrounding the rootlets was thicker than surrounding normal tissues, a finding also observed with crabs that cause growth anomalies in corals. 16 Determining the identity of this crustacean and whether it caused the skeletal growth anomaly would require additional investigations.
We also identified 2 new organisms in coral tissue, regardless of disease state, that were not associated with any evident host cell pathology. As such, we suggest that these organisms are new putative symbionts in corals. The first of these were found in Porites and Montipora and contained distinct nematocysts (stenoteles) characteristic of hydrozoa. 13 Symbiotic hydroids have been found attached to the surface body wall of corals, 19,23 but no cnidaria have been reported deep in the skeleton as seen here. The other putative symbiont was a distinct, small, multicellular organism (metazoan) in the mesoglea of M. grisea. This metazoan was smaller and within the mesoglea, distinguishing it from another metazoan in this genus, a chimeric parasite from M. capitata that is located within gastrovascular canals and is associated with tissue loss. 43 Elucidating the identity of both organisms may be difficult, as they were found infrequently and in low numbers.
The pattern of prevalence of cell-associated microbial aggregates in corals in Micronesia mirrored that seen elsewhere in the Pacific, 37 with Platygyra, Acropora, and Porites being commonly infected with total absence of these organisms in Montipora. This study expands the range of species infected with these aggregates to include A. cerealis, A. surculosa, G. fascicularis, and H. exesa. Cell-associated microbial aggregates are tissue-associated bacteria that are found in high prevalence in dominant genera of corals in the Indo-Pacific, such as Pocillopora, Acropora, and Porites. They are thought to be facultative symbionts that are important to coral health and immunity; they could also have an important evolutionary role, possibly contributing to the dominance of specific coral genera on coral reefs. 37 The documentation of their presence in Micronesia expands their range in the Pacific, and confirming their role in coral biology merits further study.
Twenty-eight percent of paired grossly normal fragments had a microscopic lesion. This phenomenon has been documented elsewhere and indicates that some coral lesions at the microscopic level will extend to tissues that appear grossly normal. 45 For example, in this study, 11% of paired normal fragments from lesions with fungi also had fungal infections associated with cell pathology, suggesting that fungal infections may be more widely disseminated than suggested by gross lesions alone. In other studies, certain microscopic lesions—such as chimeric parasites in M. capitata with tissue loss (in Hawaii)—were found to be more systemically distributed and likely to be seen in both normal and lesional tissues than ciliates or helminths, which were more restricted to grossly abnormal tissues. 45 Therefore, gross observations do not always provide clear-cut results between healthy and diseased tissues. However, the findings here provide important baseline information on coral disease and should serve as a foundation for future investigations to understand the role of particular agents in causing lesions.
Footnotes
Acknowledgements
Dr Douglas Fenner graciously assisted with the identification of corals. We thank Ray Dalio and the Dalio Family Foundation for their support of the Woods Hole Oceanographic Institution Access to the Sea program, through which this work was partially funded, and the crew of the R/V Alucia for logistical support. Susan Knowles and anonymous reviewers provided constructive comments. Mention of products or trade names does not imply endorsement by the US government.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.