
Abstract
Extremely poor growth of some individuals within a birth cohort (runting) is a significant problem in crocodile farming. We conducted a pathological investigation to determine if infectious disease is associated with runting in farmed saltwater crocodiles (Crocodylus porosus) and to look for evidence of other etiologies. In each of 2005 and 2007, 10 normal and 10 runt crocodiles, with an average age of 5.5 months and reared under identical conditions, were sampled. Laboratory testing included postmortem; histological examination of a wide variety of tissues (with quantitation of features that were noted subjectively to be different between groups); hematology; serum biochemistry (total protein, albumin, globulins, total calcium, phosphorus, and iron); bacterial culture of liver and spleen (2005 only); viral culture of liver, thymus, tonsil, and spleen using primary crocodile cell lines (2007 only); and serum corticosterone (2007 only). The only evidence of infectious disease was mild cutaneous poxvirus infection in 45% of normal and 40% of runt crocodiles and rare intestinal coccidia in 5% of normal and 15% of runt crocodiles. Bacterial and viral culture did not reveal significant differences between the 2 groups. However, runt crocodiles exhibited significant (P < .05) increases in adrenocortical cell cytoplasmic vacuolation and serum corticosterone, decreased production of bone (osteoporosis), and reduced lymphoid populations in the spleen, tonsil, and thymus. Runts also exhibited moderate anemia, hypoalbuminemia, and mild hypophosphatemia. Taken together, these findings suggest an association between runting and a chronic stress response (hyperactivity of the hypothalamic-pituitary-adrenal axis).
Keywords
In the crocodilian farming industry, extremely poor growth of some hatchlings compared with their birth cohort (runting) is a major problem worldwide, affecting up to 30% of animals. 3,21,32,33,48,49,55,58 Runting is a cause of major economic loss, in an industry where juvenile growth and survival are important economic factors. 33
The cause of runting in farmed crocodiles is unknown. Although various possibilities have been proposed, none have been thoroughly investigated. Proposed factors associated with runting include egg incubation conditions; 38,69 poor yolk absorption posthatching; 22,32 inappropriate diet or problems with diet assimilation; 11,22,26,55 posthatching environment, including temperature, stocking density, and behavior of conspecifics; 26,32,39,48,58 and failure to adapt to the captive environment. 3,64 Poor growth has been found to be clutch related in that some clutches produce relatively high numbers of runts. 26,33,48,58 However, future crocodile runts are not identifiable at hatching, since body size at hatching is a poor predictor of posthatching growth. 26,34,38,69
There is limited information on diseases and pathology exhibited by runt crocodiles. A few studies mention bacteremia, mycotic dermatitis, 64 hepatitis, 22,64 atrophy of the liver and intestine and ascites, 21,32 hepatic lipidosis, 21,49 or pancreatic atrophy. 22 There has been no in-depth pathological investigation comparing runt crocodiles with their normal counterparts from the same cohort and raised under identical conditions. The purpose of this study was to conduct a direct and thorough pathological comparison between runt and normal crocodiles to rule out the involvement of infectious disease and to survey for other lesions or conditions that could suggest a cause for the runting.
Materials and Methods
The study location was a large saltwater crocodile (Crocodylus porosus) farm 40 km south of Darwin, Australia. The weather in this region is tropical, with a “wet” season having high humidity and monsoonal rains, from November to April, and a “dry” season, characterized by low humidity and no rain, from May to October. The daily maximum temperature is 33°C during both seasons, but the average daily minimum is 24°C in the wet season compared with 18°C in the dry season. Saltwater crocodiles nest in northern Australia from late October to April, corresponding to a hatching period from February to June. Eggs were obtained both from the wild (13 runt and 10 normal crocodiles; average estimated embryo age at collection was 21 days) and from captive females (7 runt and 10 normal crocodiles; average estimated embryo age was 6 days). At the farm, eggs were incubated at 32°C and 99% to 100% humidity until hatching. Each hatchling was identified to clutch of origin by scute cutting. 30 The hatchlings used in this study originated from 35 clutches (1 from each clutch, except for 5 clutches from which 2 study animals originated from each clutch).
Runt crocodiles were not identifiable at hatching. On crocodile farms, normal management practice involves “grading” animals to ensure minimal size variation within each pen to reduce intraspecific aggression and maximize equal opportunity to access food. Grading on our study farm commences when disparity in growth starts to become apparent, at approximately 2 months of age, and is performed as required but at least every month within each pen. Thus, through systematic grading, the runt crocodiles used in this study were continually maintained in pens of similar sized animals and were not selected from pens of larger animals whereby exogenous factors such as competition for food or thermoregulatory ability could be a cause of the runting. Runt crocodiles were reared under identical husbandry conditions to normal crocodiles of the same birth cohort, with pens containing crocodiles of either of the 2 groups interspersed in the same shed. Stocking density in all pens was approximately 14 animals/m2. Within each completely enclosed shed, pens were shaded, of concrete construction, with shallow water (approx. 70% of pen area; 30–50 cm deep) at one end with a gradual ascent to a dry feed-deck (approx. 30% of pen area) at the other end. Hide-boards were provided within each pen. Water temperature in the pens was kept at approximately 32°C. The diet consisted of finely minced red meat (horse or buffalo) fortified with 2% vitamin-mineral mix that included vitamin D3 (25 000 IU/kg; Monsoon Crocodile Premix, Darwin, Australia) and 1.5% calcium carbonate. The animals were fed, in excess, 5 times per week until they were 3 months of age when they were fed every second day. Food was dispersed evenly over the land area in the afternoon and left overnight. Crocodiles were not observed for individual food intake, and measurement of remaining food as an indicator of how much the crocodiles in each pen were eating was impractical, due to dispersion of the food into the water by the crocodiles. Crocodiles were fasted for 48 hours prior to sampling. Ten runt and 10 normal crocodiles were sampled in each of 2 study years: on November 8 to 11, 2005 (case Nos. 1–20) and on July 10 and 12, 2007 (case Nos. 21–40). Sampling of animals was done when the grading process had resulted in entire pens containing subjectively obvious runt crocodiles, which was at approximately 7 and 5 months of age in 2005 and 2007, respectively. The only difference between the “runt” and “normal” animals was their body size and condition; all animals used in the study were subjectively judged to be bright, responsive, and active.
The initial suite of laboratory testing for 2005 samples was designed to be relatively broad to maximize the chance of detection of any disease or condition that could be related to runting. The purpose of the second sample in 2007 was to substantiate potentially significant findings from the initial 2005 sample, and therefore testing in 2007 differed slightly and was generally not as broad (specific differences in testing between the years is detailed in relevant sections below).
Necropsy and Histopathology
Following blood sampling (see below), crocodiles were euthanized by overdose (80 mg/kg) of pentobarbitone sodium injected into the dorsal tail vein (Lethabarb euthanasia injection, 325 mg/ml; Virbac Animal Health, Milperra, NSW, Australia). A full necropsy was performed on all crocodiles. In 2005, liver weights were taken on all crocodiles (not repeated in 2007 due to no difference between groups in 2005; see Results).
Tissues were fixed in 10% neutral buffered formalin, processed in standard fashion for histological examination, and 5-μm sections stained with hematoxylin and eosin (HE). Sections of bone were decalcified in 10% neutral buffered formalin with 9% formic acid prior to processing. Sections trimmed for histological examination were standardized with respect to location in, and orientation of, the organ/tissue. Organs/tissues examined only in 2005 were heart, lung, trachea, kidney, gallbladder, esophagus, stomach, coelomic fat body, thyroid gland, skin, femorotibial joint, spinal cord, brain, and eye. These tissues did not display notable differences between runt and normal crocodile groups (see Results) and therefore were not examined in the second set of samples taken in 2007. Organs/tissues examined in both years were pituitary gland, adrenal gland, spleen, thymus, tonsil, bone (mid-sagittal section of proximal tibial metaphysis, including bone marrow), skeletal muscle, liver, pancreas, duodenum, jejunum, colon, and gonad. Histological examination of the gonad was used to determine sex. Sections of parathyroid gland were available for examination in 4 normal and 9 runt crocodiles. Perls’s stain for ferric iron was used to identify the nature of the green-brown pigment present in splenic macrophages. 9
All histology slides were examined by one person (C.S.) unaware as to whether the tissues were from a normal or runt crocodile. Following initial screening of all slides, aspects of tissues that subjectively seemed to differ among crocodiles were quantified. Features that were scored as 0 (none), 1 (mild), 2 (moderate), or 3 (marked) were degree of cytoplasmic vacuolation of adrenocortical (interrenal) cells and hepatocytes, amount of zymogen in pancreatic acinar cells, and amount of globular green-brown pigment in splenic macrophages. Bone marrow total percent cellularity and the relative proportion of erythroid cells (compared with myeloid cells) were subjectively estimated.
Digital images were taken for the following histological features (MicroPublisher 3.3; Q Imaging, Surrey, BC, Canada) and the feature measured using a program calibrated to the magnification of the microscope (analySIS Five; Soft Imaging System GmbH, Münster, Germany). Skeletal muscle fiber width was measured by taking the average width of 10 fibers in the same region of cranial thigh muscle in each crocodile. In 2005, the muscle was sectioned transversely across the fibers, while in 2007, the muscle was sectioned longitudinally. The amount of the primary spongiosa in the proximal tibial growth zone was quantified by determining the proportion of the total width of mineralizing cartilaginous trabeculae immediately beneath the growth zone compared with the width of the medulla at the same level. The area of periarteriolar lymphoid sheaths was measured by taking the area of each of 10 lymphoid sheaths, minus the area of the arteriole each surrounded, and calculating an average. The 10 sheaths were the first 10 discrete sheaths encountered in a transect of the histological section starting from the capsule moving toward the center at the greatest diameter of a section taken of the mid-region of the spleen. The amount of lymphoid tissue in the tonsils was measured as a proportion of the area of the tonsil occupied by lymphocytes compared with the total area of the folds of the tonsil observed at low power in a standard complete transverse section taken from the mid-tonsil. Thymus tissue was quantified by measuring the total area of thymus lobes in a transverse section containing all the tissues bound by the mediastinum at the level of the proximal primary bronchi. To take into account the smaller overall size of tissues of runt crocodiles, the area of the thymus lobes is presented as a proportion of the area of the adjacent primary bronchus.
Ancillary Diagnostic Testing
Blood was sampled from the occipital venous sinus and the initial 0.5 ml of blood placed into EDTA anticoagulant for hematology, with the remainder placed into serum separator gel tubes (BD Vacutainer; Becton Dickinson, Franklin Lakes, NJ). EDTA was used as the anticoagulant since in our experience, it does not cause hemolysis in saltwater crocodiles and is not potentially associated with leukocyte and platelet clumping, and thus inaccurate cell counts, as has been reported with lithium heparin anticoagulated blood. 5 Blood samples were obtained 2 to 4 hours after removal from the pen for crocodiles in 2005 and within 3 minutes of removal from the pen, between 0800 and 0900 in the morning for crocodiles in 2007. The strict timing of blood sampling for 2007 was to minimize any effects of circadian rhythm or acute handling stress on serum corticosterone level. 23,37,42,44 Serum corticosterone was measured in the 2007 samples in an effort to expand on the apparent histological difference noted in adrenal glands between the runt and normal crocodile groups noted in 2005 (see Results). Frozen serum was not available to retrospectively test the 2005 crocodiles. Serum corticosterone was measured using a high-sensitivity enzyme immunoassay according to kit directions (Corticosterone HS EIA; Immunodiagnostics Systems Ltd, Boldon, Tyne & Wear, UK). Values that exceeded the 20-ng/ml upper limit of kit accuracy were set at this limit for statistical analysis.
Selected clinical pathological parameters were determined to assist interpretation of aspects of the gross and/or histopathology. Hematology included total and differential white blood cell counts conducted using a hemocytometer with an eosinophil Unopette system (Becton-Dickinson, Rutherford, NJ) and blood smear examination according to standard reptile protocol. 6,10 Packed cell volume was determined by centrifugation of blood in microhematocrit tubes. Serum biochemical parameters were determined on an automated analyzer (Konelab 20; Thermo Electron, Victoria, Australia). Parameters measured (followed in parentheses by a brief analytical basis for the measurement provided by Thermo Electron) were total protein (biuret method), albumin (bromcresol green dye binding method), globulins (by subtraction of albumin from total protein), total calcium (reaction with the metallochromogen Arsenazo III), inorganic phosphorus (formation of phosphomolybdate and subsequent reduction to molybdenum blue), and iron (hydroxylamine hydrochloride reduction and subsequent reaction with liquid ferrozine).
Bacterial culture of liver and spleen was performed on all crocodiles in 2005 but not in 2007 because of a lack of significant bacteriological differences between the 2 groups in 2005 (see Results). Culture was performed on samples obtained aseptically during necropsy. To collect samples aseptically, the skin of the crocodile was cleaned with 100% ethanol and incised using scalpel and forceps that had been sterilized by dipping them in 100% ethanol and flaming them over a Bunsen burner. Following the skin incision and reflection of the skin, the instruments were again sterilized and the coelom opened. Instruments were sterilized a third time prior to sampling tissue. In addition, yolk was cultured from case No. 1 (runt) and case No. 11 (normal) with enlarged internal yolk sac remnants and a swab of the subcutis from the swollen forelimb of case No. 16 (normal). For these samples, the tissue was entered aseptically as described above and sampled using sterile swabs. Bacterial culture was performed using standard veterinary bacteriology phenotypic and biochemical techniques. Briefly, samples were homogenized and plated onto sheep blood agar (Oxoid Australia, Thebarton, Australia) and MacConkey agar (Oxoid Australia, Thebarton, Australia) and incubated at 35°C for 48 hours. The bacterial isolates were initially characterized using Gram’s stain, colony morphology, and relevant preliminary tests, including oxidase and catalase, and then the appropriate commercial kits were used for speciation (api 20 Strep, api Coryne, bioMérieux, Marcy-l’Etoile, France; Microbact Gram-Negative Identification System, Oxoid Ltd, Basingstoke, Hants, UK).
Virus isolation was attempted on liver, thymus, tonsil, and spleen from all crocodiles in 2007 using primary crocodile liver and kidney cell lines developed at Berrimah Veterinary Laboratories. 50 Culture was not performed on samples from 2005 since the crocodile cell lines had not been developed yet and frozen stored samples from 2005 were no longer available. In 2007, samples were stored at –70°C until processing. Briefly, tissue samples were homogenized, clarified by centrifugation, and the supernatant filtered through a 0.45-μm filter. The filtered supernatant was then inoculated into 25-cm2 flasks containing confluent primary cell line monolayers. Each sample was inoculated into 2 different cell lines. The flasks were incubated at 28°C and examined for viral growth every 3 days for 21 days. The cultures were then passaged into fresh flasks for another 21 days and examined every 3 days. This process was repeated a final time, and if no growth was observed, the culture was deemed negative. Viral growth was recognized by a cytopathic effect characterized by loss of confluence of the cell monolayer, rounding up of cells, and, in some cases, syncytia formation. 50 The virus isolations were part of a larger study investigating the viral flora of saltwater crocodiles, 50 and further characterization of isolated viruses was beyond the scope of this study.
Statistical Analyses
To generate a single continuous variable that described overall body size relative to age (a growth index), we first conducted a principal component analysis to combine 2 measures of body length (snout-vent length and vent-tail tip length) with body weight. The resultant first principal component (PC1) incorporated 99% of the variation in the 3 measures and was used as a single measure of overall body size. This PC1 body size measure was then corrected for crocodile age by calculating the residuals from a linear regression of PC1 on age. These residual values form the growth index, as they provide a composite measure of how large each crocodile was for its age. There was no overlap in this value for crocodiles assigned to the normal and runt groups, verifying its validity in distinguishing runts from normal crocodiles (Table 1). This continuous variable was used as an explanatory covariate in all statistical analyses. However, for ease of interpretation, summary statistics of basic measurements are presented as 2 groups: normal and runt crocodiles.
Body Size, Age, Sex, Hematological, and Biochemical Parameters for All Normal and Runt Crocodiles in 2005 (Case Nos. 1–20) and 2007 (Case Nos. 21–40).a
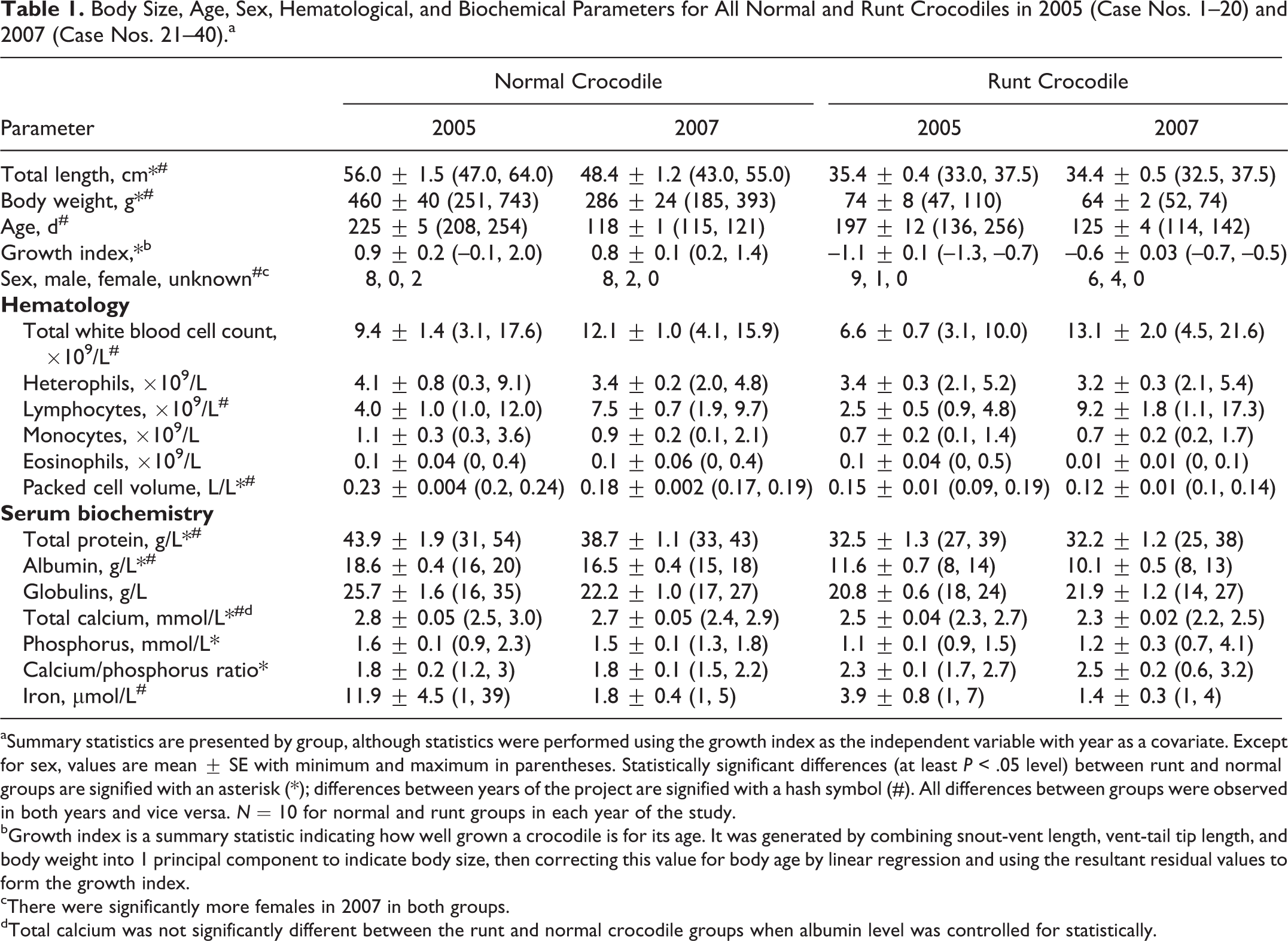
aSummary statistics are presented by group, although statistics were performed using the growth index as the independent variable with year as a covariate. Except for sex, values are mean ± SE with minimum and maximum in parentheses. Statistically significant differences (at least P < .05 level) between runt and normal groups are signified with an asterisk (*); differences between years of the project are signified with a hash symbol (#). All differences between groups were observed in both years and vice versa. N = 10 for normal and runt groups in each year of the study.
bGrowth index is a summary statistic indicating how well grown a crocodile is for its age. It was generated by combining snout-vent length, vent-tail tip length, and body weight into 1 principal component to indicate body size, then correcting this value for body age by linear regression and using the resultant residual values to form the growth index.
cThere were significantly more females in 2007 in both groups.
dTotal calcium was not significantly different between the runt and normal crocodile groups when albumin level was controlled for statistically.
Response variables that were measured as ordinal categories (eg, degree of cytoplasmic vacuolation of adrenocortical cells) were analyzed using logistic regression with year and growth index as explanatory variables. Measurements made on a continuous scale (eg, hematology parameters, size of splenic periarteriolar lymphoid cuffs) were analyzed using multiple regression, with year and growth index as explanatory variables.
Since 40% to 45% of total calcium is transported in serum bound to protein (principally albumin), serum albumin level may influence total calcium level. 1,51 To correct for this, we regressed total calcium on albumin measures. The residual values from this regression were then compared between the runt and normal crocodiles using analysis of covariance.
Results
Necropsy
The runt and normal crocodile groups differed significantly in total length and body weight in both years, with runt crocodiles being, on average, 33% shorter and 82% lighter than normal crocodiles (Fig. 1, Table 1). There was no significant difference in the ages of runt vs normal crocodiles; however, both groups were older in 2005. The growth index differed significantly between the runt and normal crocodile groups, with no overlap in range, confirming significantly smaller body size for age (ie, poorer growth) in runt crocodiles. Overall, there was no difference in sex ratio between the 2 groups, but in 2007, there were more females in both groups than there had been in 2005 (Table 1). There was no significant difference in the origin of the eggs (wild vs captive nest) between the 2 groups.

Normal crocodile Nos. 21 to 25 (left) and runt crocodile Nos. 26 to 30 (right), sampled July 10, 2007.
Visually, runt crocodiles exhibited reduced muscling, prominence of bony protuberances, reduced amount of adipose tissue in the base of the tail, and markedly reduced size of the coelomic fat body (which was not grossly visible in many of the runts). These subjective necropsy findings indicate poor body condition in runt compared with normal crocodiles. Compared with normal crocodiles, gallbladders of runt crocodiles were relatively large and distended with bile. Bones were subjectively judged to be of comparable strength between the 2 groups. Grossly, lobes of the thymus were generally unapparent in runt crocodiles while they were usually obvious in normal crocodiles. The tonsils of runt crocodiles had less prominent folds than those of normal crocodiles. Case No. 1 (runt) and case No. 11 (normal) had 1-cm diameter yolk sac remnants protruding from the outer wall of the mid-jejunum. The remainder of the crocodiles either had a small (1–2 mm diameter) yolk sac remnant or the remnant was not grossly appreciable. One normal crocodile (case No. 16) had moderate diffuse enlargement and edema involving 1 forelimb. Except for small fecal pellets in the rectum, the gastrointestinal tracts were largely empty in both groups. There were rare to occasional 0.3- to 1.0-mm round white cutaneous foci, typical of mild superficial poxvirus lesions, in case Nos. 2, 6, 13, 14, 27, 29, 30, and 32 (8 runt crocodiles) and case Nos. 5, 9 to 12, 16, and 38 to 40 (9 normal crocodiles). Liver weight as a percentage of body weight averaged 2.4% in both the runt and normal crocodile groups.
Histopathology
Except for features that are expanded upon below, there were no differences in histological appearance of organs/tissues among the crocodiles. The significance of findings between normal and runt crocodiles applies to both years of the project unless otherwise stated.
There were rare sporulated coccidia oocysts in the mucosal epithelium of the jejunum and/or colon in case Nos. 2, 6, and 15 (runts) and case No. 25 (normal). Histologically, the poxvirus lesions appeared as discrete slightly raised foci of markedly hypertrophic epithelium containing large eosinophilic intracytoplasmic inclusion bodies, typical of mild crocodile poxvirus lesions. 33,35 Histologically, the single swollen leg present in case No. 16 (normal) exhibited marked subcutaneous and intramuscular edema, associated moderate heterophil and macrophage infiltration, and numerous macrophages containing abundant intracytoplasmic bacterial cocci.
Statistics for histological features that were quantified are presented in Table 2. Degree of vacuolation of adrenocortical cells was significantly greater in runt compared with normal crocodiles and slightly greater in both groups in 2007 (Figs. 2, 3). The degree of hepatocyte vacuolation was slightly but significantly less in runt crocodiles and less in both groups in 2005. The degree of pancreatic zymogen was significantly decreased in runt crocodiles. Skeletal muscle fiber width was significantly less in runt crocodiles compared with normal crocodiles in both years. Muscle fiber width was significantly less in both groups in 2007, presumably artifactually, due to the muscle being sectioned longitudinally in 2007, compared with transversely in 2005. Bone marrow cellularity was reduced in runt compared with normal crocodiles with no difference in the proportion of erythroid cells between the 2 groups. There was an increase in the proportion of erythroid cells in the bone marrow in both groups in 2005.
Histological Features Quantified for All Normal and Runt Crocodiles in 2005 (Case Nos. 1–20) and 2007 (Case Nos. 21–41).a
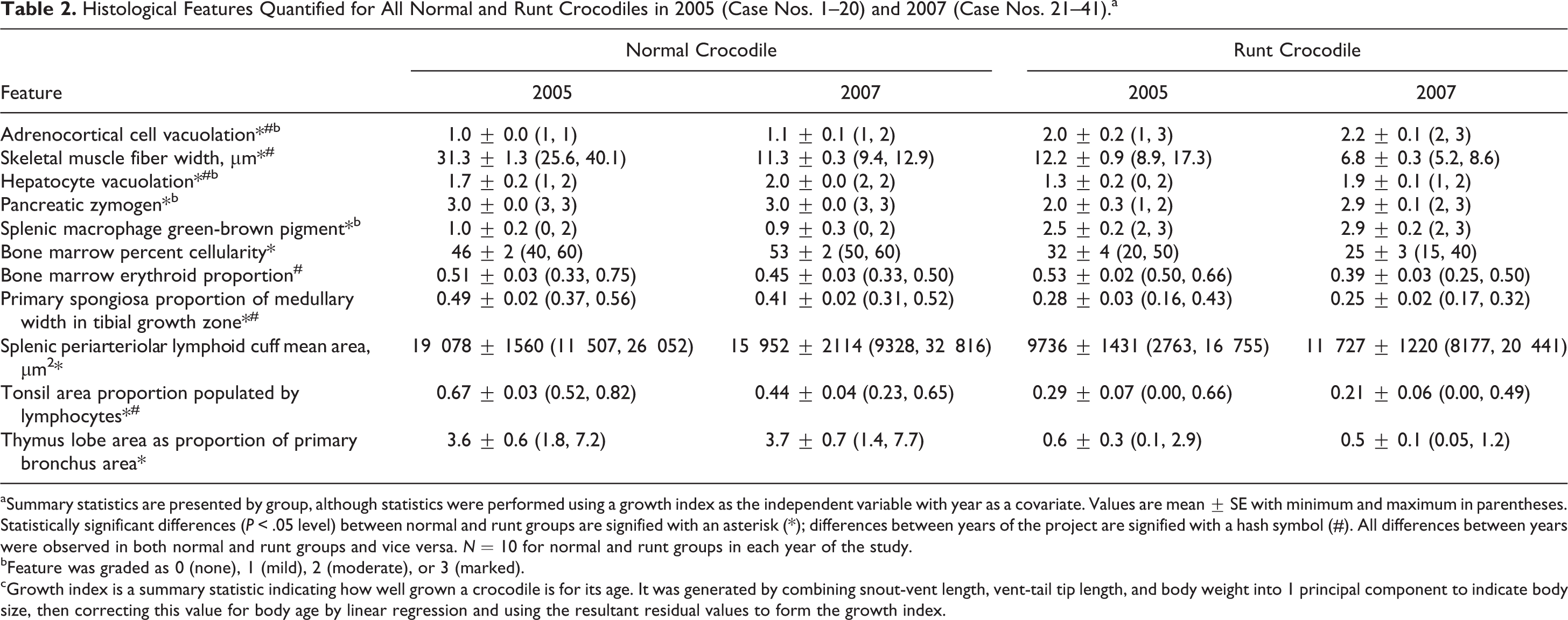
aSummary statistics are presented by group, although statistics were performed using a growth index as the independent variable with year as a covariate. Values are mean ± SE with minimum and maximum in parentheses. Statistically significant differences (P < .05 level) between normal and runt groups are signified with an asterisk (*); differences between years of the project are signified with a hash symbol (#). All differences between years were observed in both normal and runt groups and vice versa. N = 10 for normal and runt groups in each year of the study.
bFeature was graded as 0 (none), 1 (mild), 2 (moderate), or 3 (marked).
cGrowth index is a summary statistic indicating how well grown a crocodile is for its age. It was generated by combining snout-vent length, vent-tail tip length, and body weight into 1 principal component to indicate body size, then correcting this value for body age by linear regression and using the resultant residual values to form the growth index.
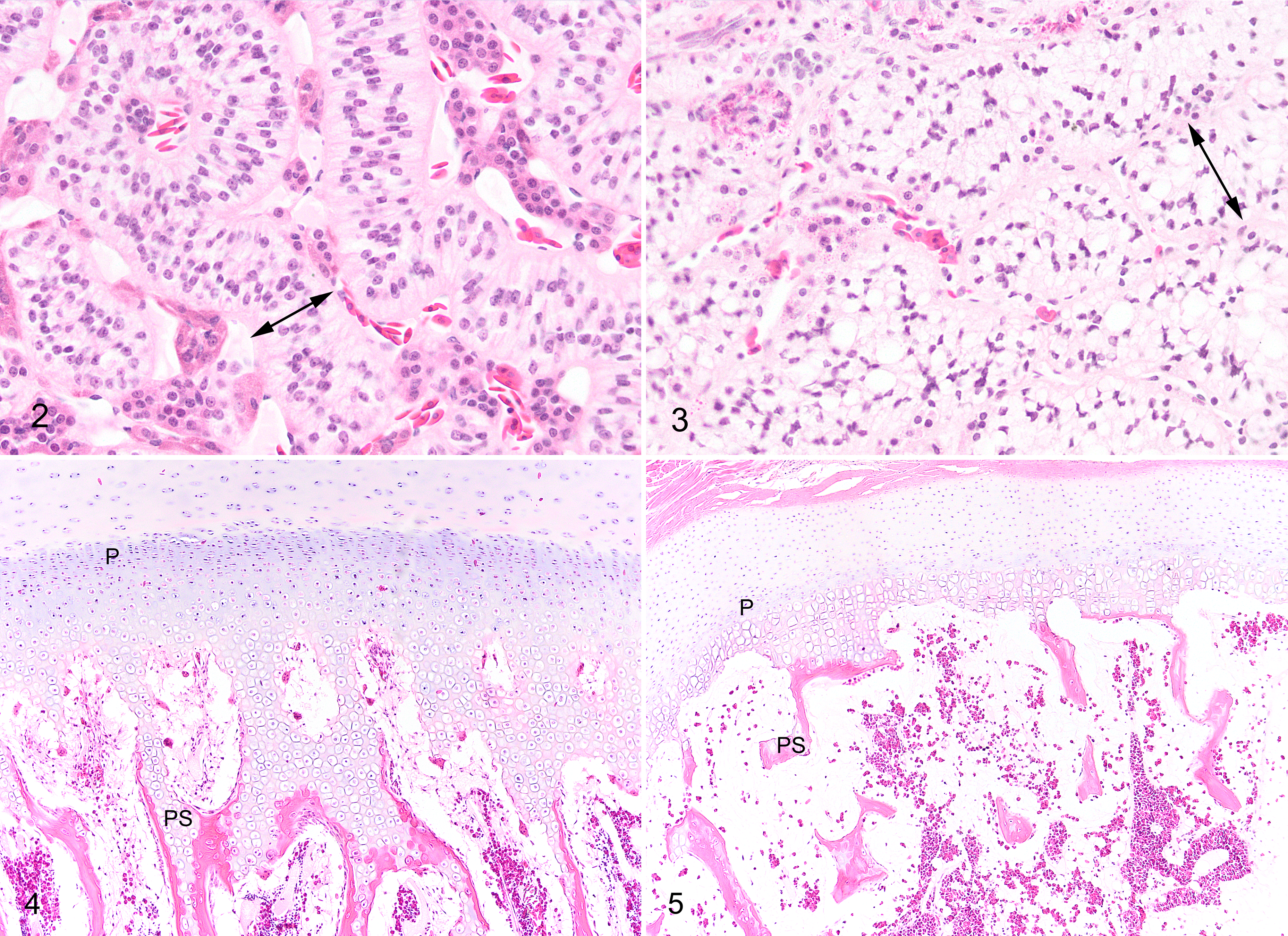
Proximal tibial bone growth zones differed subjectively in histological appearance between runt and normal crocodiles. While the growth zones of runt crocodiles retained all normal histological components with no additional lesions, runts had generally narrower and less distinct zones of proliferating, maturing, and hypertrophic chondrocytes and decreased production of primary spongiosa (Figs. 4, 5). Quantitation of the amount of primary spongiosa confirmed decreased production of bone at the growth zone in runt compared with normal crocodiles (Table 2).
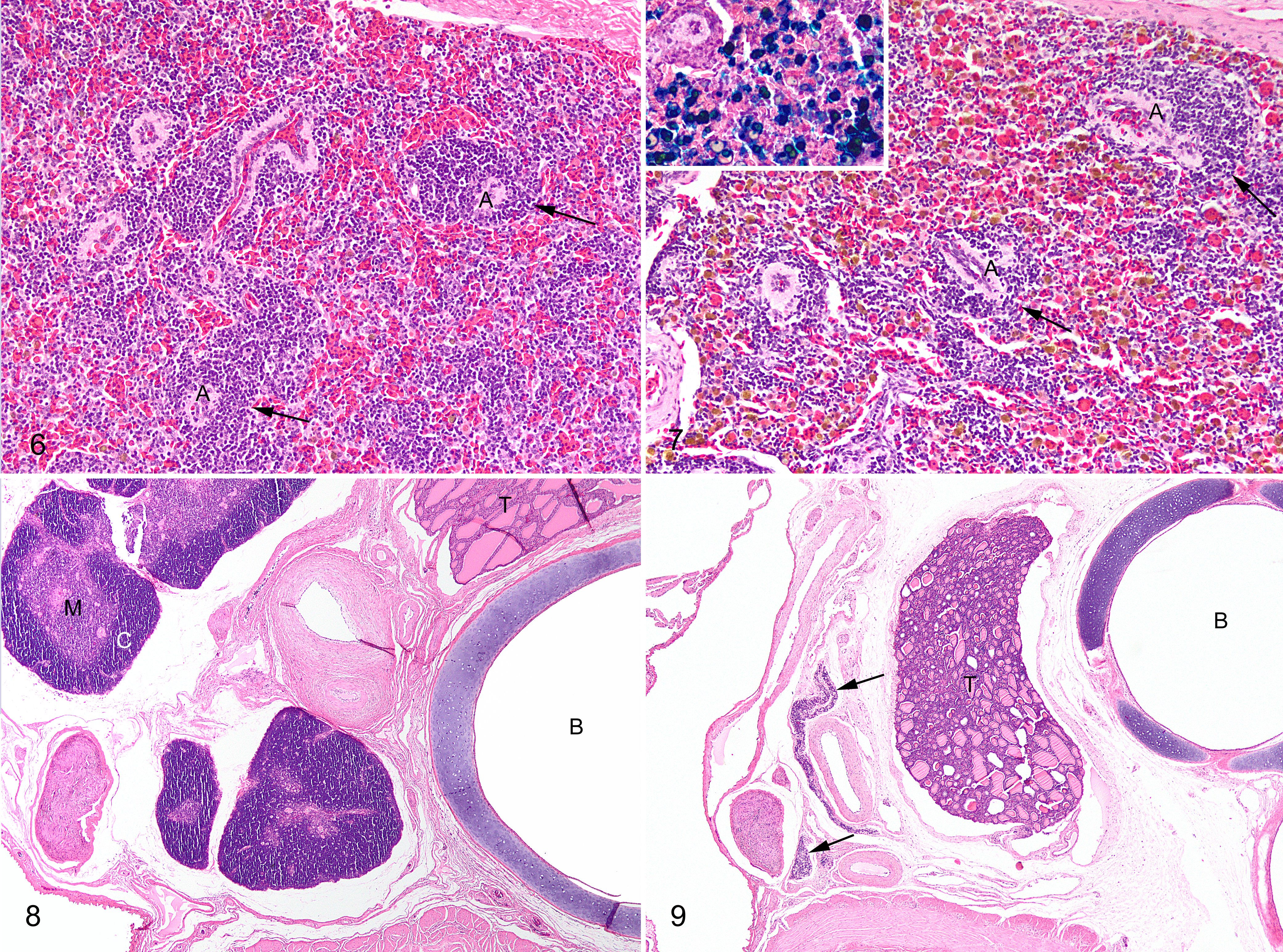
Runt crocodiles had significantly reduced lymphoid populations compared with normal crocodiles in all tissues in which this was evaluated, including, spleen (Figs. 6, 7), tonsil, and thymus (Figs. 8, 9). Tonsils had a greater lymphoid component in both groups in 2005. The amount of green-brown pigment in splenic macrophages was greater in runt compared with normal crocodiles. The majority of this pigment was strongly positive with Perls’s stain for ferric iron (Fig. 7, inset).
Ancillary Testing
In 2007 (testing was not performed in 2005), mean serum corticosterone levels were significantly higher in runt (mean [SE], 17.1 [1.1] ng/ml; range, 10.2–20) compared with normal crocodiles (mean [SE], 9.4 [2.3] ng/ml; range, 0.5–20).
Hematologically, there were no significant differences between runt and normal crocodiles in white blood cell parameters, although both groups had significantly higher total white blood cell and lymphocyte counts in 2007. Mean packed cell volume was significantly lower in runt compared with normal crocodiles and significantly lower in both groups in 2007 (Table 1). Examination of blood smears did not reveal evidence of hemoparasites in crocodiles in either group. There were low to moderate numbers of polychromatophilic red blood cells in peripheral blood smears in 11 runt and 17 normal crocodiles.
Serum biochemistry (Table 1) revealed significantly lower total protein and albumin in runt compared with normal crocodiles in both years, and these parameters were higher in both groups in 2005. Serum total calcium and phosphorus were significantly lower in runt compared with normal crocodiles, with total calcium being slightly but significantly higher in all crocodiles in 2005. However, when the residuals of a regression of total calcium vs albumin were compared between the runt and normal crocodile groups, there was no significant difference, indicating that the difference in total calcium between the 2 groups was attributable to their differing serum albumin levels. The relatively greater decrease in phosphorus in runt crocodiles compared with total calcium resulted in significantly higher total calcium to phosphorus ratio in runt compared with normal crocodiles (Table 1). There were no statistically significant differences between runt and normal crocodiles in serum iron, although this parameter exhibited substantial variation within the groups and was on average higher in both groups in 2005.
Bacterial culture results in 2005 were negative except as follows: Streptococcus agalactiae was isolated in 3 normal crocodiles (from the liver and spleen of case No. 4, the liver of case No. 8, and the swollen forelimb of case No. 16). Edwardsiella tarda was isolated from the enlarged yolk sac remnant of case No. 11 (normal). Corynebacterium sp was isolated from the liver and spleen of case No. 14 (runt) and Morganella morganii from the liver and Salmonella sp from the enlarged yolk sac remnant in case No. 1 (runt).
Cytopathic effect in viral culture suggestive of viral growth was observed in 2007 from the liver and tonsil of case No. 22 (normal) and from the tonsil of case Nos. 21, 24, and 25 (normal). No cytopathic effect in viral culture was detected from runt crocodiles.
Discussion
Runt crocodiles in this study exhibited markedly smaller body size and weight for their age compared with normal crocodiles, indicating very poor growth as noted in other studies. 3,21,32,48,49,55,58 In addition, multiple parameters indicated inanition in runt crocodiles, including their moderately decreased serum albumin, smaller skeletal muscle fiber width, and decreased pancreatic zymogen. Liver weight as a percentage of body weight was the same in runt and normal crocodiles, suggesting that a significant degree of hepatic atrophy was not present in runt crocodiles. Histologically, runt crocodiles had slightly decreased hepatocyte cytoplasmic vacuolation compared with normal crocodiles. In reptiles, mild to moderate hepatocyte vacuolation, usually due to lipid, is commonly present in reptiles in good body condition and is considered within physiological normal, rather than a degenerative change. 30,59 Thus, the mildly decreased hepatocyte vacuolation in runt compared with normal crocodiles likely reflects their generalized paucity of body fat and is another parameter supporting inanition in the runt crocodiles.
Runt crocodiles had significantly lower packed cell volumes than normal crocodiles. The anemia in runts was nonregenerative, being unaccompanied by evidence of erythroid hyperplasia in bone marrow or evidence of regeneration in peripheral blood beyond the mild to moderate red blood cell polychromasia that was observed in both runt and normal crocodiles. This may be at least in part due to the tendency of young reptiles to exhibit a greater degree of polychromasia than adults. 5 Serum iron was inconsistent between the 2 years, with 2005 crocodiles having significantly higher serum iron levels than 2007 crocodiles regardless of whether they were runt or normal animals. Both groups also had lower packed cell volumes in 2007, possibly reflecting lower serum iron. In the runt crocodiles, there appeared to be sequestration of iron in the form of splenic hemosiderin. Splenic hemosiderosis occurs with starvation or other catabolic states in all species and, in reptiles, is generally considered secondary to other conditions, such as chronic inanition. 46 Thus, it is likely that inanition is the main contributor to the anemia in runt crocodiles. 62,67
One of the main objectives of this study was to investigate whether chronic infectious disease could be associated with runting. Despite a thorough examination of all organ systems in runt vs normal crocodiles, there were no gross or histological lesions to suggest a significant presence of infectious disease. The only parasite detected in the study was rare sporulated oocysts in the intestinal epithelium of 3 runt crocodiles and 1 normal crocodile, consistent with the Goussia-like coccidia described in saltwater crocodiles in Australia. 25,40 The presence of this parasite as a contributing factor to runting is dubious, given that the coccidia were evident in only 3 of 20 runt crocodiles, and then only rarely in the intestinal epithelium with no associated histological lesions.
Bacterial culture of 2 filtering organs (liver and spleen) was conducted to detect bacterial sepsis, since this may be difficult to detect histologically and is a common cause of morbidity and mortality in juvenile crocodiles. 3,41 Bacterial isolates were limited to a total of 5 species in 2 runt crocodiles and 4 normal crocodiles, with no particular pattern to suggest a relationship with runting.
Histological evidence of virus infection was limited to mild cutaneous poxvirus infection present in several crocodiles, both normal and runts. Virus isolation was attempted for multiple internal organs in all crocodiles in 2007 to further substantiate lack of virus involvement and because of the association of viral infection with atrophy of lymphoid tissues and stunted growth in other species. Examples include mammalian parvoviruses and pestiviruses, 47 as well as retroviruses of the reticuloendotheliosis virus group in poultry. 18 However, our isolation attempts yielded only a cytopathic effect suggestive of 5 isolates from 4 normal crocodiles. This, as well as the lack of histological lesions such as necrosis or inflammation that could substantiate a significant virus infection, suggests that viral infection was not related to runting in our study.
A second objective of the study, to look for histological clues of other disease processes, revealed some associations with runting and could help suggest an alternate etiology to infectious disease. One notable finding was the increased cytoplasmic vacuolation of adrenocortical cells in runt compared with normal crocodiles in both phases of the study. Variation in the degree of vacuolation is evidence of altered activity of the adrenocortical cells in reptilians and is evidence of increased activity of the cells in mammals. 7,24 To further investigate this finding, in 2007, serum level of corticosterone, the main glucocorticoid stress hormone in crocodilians, 37,43 was measured and found to be significantly elevated in runt crocodiles. Chronic elevation of corticosteroids results in a general catabolic state 7,61 and anorexia in crocodiles, 32 and it inhibits both growth hormone secretion and action in mammals. 31 A negative correlation between corticosterone level and growth rate has been noted previously in saltwater crocodiles 58,65 and alligators, 15 and extremely poor growth has been experimentally reproduced in alligators by chronic elevation of corticosterone. 43,53
Another notable histological finding in runt crocodiles was the presence of bone growth zones that had all the components of normal growth zones but were relatively quiescent and exhibit decreased production of primary spongiosa. The runt crocodiles were mildly hypocalcemic and moderately hypophosphatemic compared with normal crocodiles. Since approximately 40% to 45% of total calcium is transported in serum bound to protein (principally albumin), the mild hypocalcemia in the runt crocodiles was suspected to be a reflection of their concurrent substantial hypoalbuminemia (runt crocodiles had on average 38% lower albumin than normal crocodiles). 1,51 This was confirmed by statistically correcting for the level of albumin. Measurement of the ionized (unbound, biologically active) portion of total calcium would have been an additional parameter we could have used to investigate serum calcium in this study 1,51 but was judged to not be worth pursuing given the lack of hypocalcemia in the runts once albumin was taken into account.
The mild to moderate hypophosphatemia in runt crocodiles is likely at least in part attributable to prolonged lack of dietary intake, a common cause in reptiles. 4 Since runt crocodiles were offered the same diet as the normal crocodiles, the deficit may be due to a lack of food intake rather than a deficient diet per se. Also, increased glucocorticoid hormone has been shown in humans to result in decreased renal resorption and gastrointestinal absorption of phosphate. 57 A final possible cause for the relative hypophosphatemia in runt crocodiles may be that it is simply reflecting decreased bone formation, as serum phosphorus tends to be higher in growing animals. 1
The formation of endochondral bone in growth zones is essentially similar in mammals, birds, and reptiles. 28 In mammals, abnormal bone growth in young animals due primarily to severe prolonged lack of dietary phosphorus generally results in rickets or osteomalacia, characterized by deformities of bone shape with excessive production of unmineralized osteoid and cartilaginous matrix, 63 lesions that were not evident in runt crocodiles. The histology of the poor bone growth in the runt crocodiles is best described as osteoporosis, with relative inactivity of the growth zone and decreased production of bone being typical findings in mammals or reptiles undergoing inanition. 28,63 Another factor in reduced activity of growth zones in runt crocodiles may be elevated corticosterone, since elevated glucocorticoid hormones leads to osteoporosis and decreased bone formation in some mammals. 56,57,63
The decreased lymphoid populations in the major immune tissues of crocodiles, 19 including spleen, tonsil (gut-associated lymphoid tissue), and thymus in runt crocodiles, indicate generalized lymphoid hypoplasia or atrophy. Marked lymphoid involution in reptiles can be seasonal or may result from stressors such as starvation or disease. 10 Corticosterone has been shown to be associated with lymphoid involution and immunosuppression in a wide variety of species, including crocodilians. 43,54,60,70 Despite the lymphoid involution evident in runt crocodiles, there was no overt evidence of immunosuppression in the form of an increased prevalence of infectious disease. This may be partly related to the sampling procedure, in that only bright, responsive, and active runt crocodiles were selected for the study. Runt crocodiles may be predisposed to eventual morbidity/mortality due to infectious disease, but this would have gone undetected in the present study and would be a useful avenue for further research. Perhaps even better would be research aimed at detecting subtle evidence of immunosuppression in runt crocodiles, such as response to phytohemagglutinin injection or other immunoassays. 20,27
The marked histological difference in lymphoid tissues of runt vs normal crocodiles was not reflected in numbers of circulating lymphocytes in this study. This is in contrast to other studies in reptiles in which tissue lymphoid depletion was correlated with a decrease in circulating lymphocytes. 62 Furthermore, the stress response, characterized by increased activity of the adrenocortical cells, has been found to be associated with a decrease in circulating lymphocytes along with an increase in heterophils/neutrophils in animals generally, including crocodilians. 5,29,53,56 Despite the histologically appreciable adrenocortical hyperplasia and significantly elevated corticosterone in runt crocodiles in this study, this was not correlated with alterations in circulating leukocytes compared with normal crocodiles. However, a few other studies have noted variable or absent alterations in numbers of circulating lymphocytes and heterophils with stress in crocodilians, indicating that measurement of circulating leukocytes may be an unreliable indicator of adrenocortical activity in these species. 43,65
In summary, there is no significant evidence of infectious disease in runt crocodiles in this study. Rather, there is evidence of inanition, lymphoid atrophy, and quiescent growth zones in bones. Regarding inanition and poor growth, although food intake of the runt crocodiles in this study was not measured, because we found no evidence of diseases or conditions that might result in poor food conversion, it seems likely that the runt crocodiles, while being offered equal access to food, were not eating as much as the normal crocodiles. As discussed previously, decreased appetite could be a consequence of a stress response. 32 Alternatively, it is possible that the food was inherently unacceptable to runt crocodiles (eg, they required the additional stimulation of movement of prey to eat), they were unable to learn to eat the captive diet, or they had altered behavior that prevented them from approaching the food (eg, excessively fearful disposition). Supporting these as possibilities is the finding by some investigators that changing the diet will ameliorate runting in some circumstances. 11,22,26,55 One avenue for further research into runting in crocodiles could thus be confirmation that runt crocodiles do eat less than their normal conspecifics and alteration of the diet or feeding environment in an attempt to ameliorate the runting.
Another possibility is that the inanition, lymphoid atrophy, and quiescent growth zones of bones are a result of a chronic adrenocortical stress response, which was demonstrated by the adrenocortical cell vacuolation and elevation of corticosterone in runt crocodiles. Chronic stress could be a secondary response to some unknown stressor. For example, one possible cause of secondary stress could be related to a perceived lack of suitable food in runt crocodiles, as discussed above. Since runt crocodiles were reared in pens containing other similarly sized runts, a chronic stress response is unlikely to be secondary to aggression from larger pen-mates. Although runt crocodiles were reared under identical husbandry conditions as normal crocodiles, it is possible that there could be something inherently different in runt crocodiles that results in them being stressed by a feature of the husbandry or environment that is not stressful to normal crocodiles. Further research along these lines could involve trials altering the features of runt pens, such as design (eg, provide more hides, variation in water depth), lowering stocking density, or providing a wider spectrum of available temperature (eg, provide heat lamps for basking). Thus, runt crocodiles could be suffering from a “maladaptation syndrome” suggested by a few other investigators and recognized as a feature of stress in reptiles. 3,12,65
Rather than being a secondary response in individual hatchlings to some aspect of the farming situation, the stress response could be a primary feature inherent in a runt crocodile. It has been established in many animals, from fish to birds and mammals, that maternal stress may influence the hypothalamic-pituitary-adrenal axis of the offspring, resulting in altered behavioral development and/or ill thrift in the offspring for several months after birth. 2,8,13,16,17,36,45 In lizards, treatment of reproductive females or eggs with corticosterone can affect hatchling body size, growth rate, behavior (eg, dispersal and antipredator responses), and even sex. 52,66,68 Likewise, in crocodilians, limited studies suggest that high plasma corticosterone in females may result in relatively poor egg quality and decreased hatchling survival. 14,61
The clutch effect in crocodile runting 26,33,48,58 suggests that crocodiles in specific clutches are hatched with a tendency to become runts, although more research needs to be done on the degree of clutch effect and variation within a clutch. A clutch effect could be due to genetics, maternal circumstances, egg incubation conditions, or, where members of a clutch are reared together in some degree of isolation from other clutches, their rearing environment. These considerations are beyond the scope of this study, given the large number of clutches the study animals originated from and the mixture of eggs from wild vs captive nests, but may be valuable avenues for further research. Exploration of the possible role of genetics in causing runting would require controlled breeding experiments in captive pairs over multiple years. Maternal circumstances, including size and body condition, geographic location, diet, food availability, and stress level, would also be useful avenues for further research. Egg incubation conditions in this study were uniform and controlled at the farm once eggs were collected but not necessarily prior to collection, particularly for eggs originating from wild nests. An effect of incubation conditions on posthatching growth has been noted in a few studies of crocodilians, 38,69 but its role in runting is an area that requires further research. Finally, if it is established that there is a strong clutch effect to runting and predictors discovered, investigation into either preventative measures or altered rearing practices for at-risk clutches might ameliorate the runting.
Footnotes
Acknowledgements
This research was approved by the University of Sydney Animal Ethics Committee (reference number N00/9-2005/3/4204).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This project was funded in part by the Australian Government, Rural Industries Research and Development Corporation and findings briefly summarized in RIRDC Publication No. 09/135 Improving Australia’s Crocodile Industry Productivity—Understanding Runtism and Survival.