
Abstract
The past decade has seen remarkable technical advances in infectious disease diagnosis, and the pace of innovation is likely to continue. Many of these techniques are well suited to pathogen identification directly from pathologic or clinical samples, which is the focus of this review. Polymerase chain reaction (PCR) and gene sequencing are now routinely performed on frozen or fixed tissues for diagnosis of bacterial infections of animals. These assays are most useful for pathogens that are difficult to culture or identify phenotypically, when propagation poses a biosafety hazard, or when suitable fresh tissue is not available. Multiplex PCR assays, DNA microarrays, in situ hybridization, massive parallel DNA sequencing, microbiome profiling, molecular typing of pathogens, identification of antimicrobial resistance genes, and mass spectrometry are additional emerging technologies for the diagnosis of bacterial infections from pathologic and clinical samples in animals. These technical advances come, however, with 2 caveats. First, in the age of molecular diagnosis, quality control has become more important than ever to identify and control for the presence of inhibitors, cross-contamination, inadequate templates from diagnostic specimens, and other causes of erroneous microbial identifications. Second, the attraction of these technologic advances can obscure the reality that medical diagnoses cannot be made on the basis of molecular testing alone but instead through integrated consideration of clinical, pathologic, and laboratory findings. Proper validation of the method is required. It is critical that veterinary diagnosticians understand not only the value but also the limitations of these technical advances for routine diagnosis of infectious disease.
Keywords
Traditionally, pathogen identification for infectious disease was mostly based on isolation in culture, and this was also critical for fulfilling Koch’s postulates. 24 Molecular identification methods have been used for infectious disease diagnosis since the 1980s, and “molecular Koch’s postulates” were applied to bacterial pathogenicity at the gene rather than the whole-organism level. 21,22 Molecular guidelines for establishing microbial disease causation were described as a reconsideration of Koch’s postulates. 24
Among the nonculture methods, polymerase chain reaction (PCR) and DNA sequencing have been widely used to identify infectious pathogens. Recently, massive parallel DNA sequencing has rapidly developed for the identification of both described and new viral pathogens. The technology will likely revolutionize bacterial disease diagnosis in the near future. Not only have many nonculturable or difficult-to-culture bacterial pathogens become detectable, but many readily isolated bacterial infections can now be identified with nonculture methods. Molecular identification methods can be both more sensitive and quicker than traditional culture methods, as well as adding value at relatively low cost, for example, for epidemiological purposes through identification of individual strains. This review focuses on the molecular identification methods commonly used in veterinary bacterial disease diagnosis. Since many molecular assays have been developed and published, this review will use selected examples and to illustrate advances in different areas. Experiences at a provincial veterinary diagnostic laboratory will be used to demonstrate how molecular assays are being routinely used. Immunohistochemistry is not discussed.
PCR Assays
Currently, PCR is the first line of molecular methods for identification of pathogens directly from different samples, including tissue samples. PCR assays require a set of primers specific for the targeted pathogen. In the past, the amplified PCR products were revealed by electrophoresis in an agarose gel (gel-based PCR). Over the last few years, most gel-based PCR assays have been replaced by real-time PCR, where the PCR product is monitored in each cycle of amplification (ie, in real-time) by the use of a double-stranded DNA fluorescent dye or a fluorescein-labeled probe. Real-time PCR has the advantage over conventional PCR of speed, as well as being less prone to cross-contamination because it is performed in a closed system. Veterinary diagnostic laboratories in North America can now offer PCR tests for more than 50 different pathogens, mostly for difficult-to-culture or unculturable pathogens, or for those that are difficult to characterize using other methods. For example, for bacterial pathogens, our laboratory offers PCR assays for the detection of Brachyspira spp, Chlamydophila abortus, Chlamydophila psittaci, Coxiella burnetii, avian Coxiella-like organism, Lawsonia intracellularis, Mycobacterium avium subsp paratuberculosis, different species of Mycoplasma, and Streptococcus equi subsp equi.
A good example of the use of PCR to identify difficult-to-culture bacteria is for molecular detection of Brachyspira, since these bacteria are not only difficult to culture but are also difficult to identify to species. Brachyspira spp have been isolated from pigs, namely, Brachyspira hyodysenteriae, Brachyspira innocens, Brachyspira intermedia, Brachyspira murdochii, and Brachyspira pilosicoli. 48 Recently, novel Brachyspira species identified in Europe and North America were provisionally designated “Brachyspira suanatina” and “Brachyspira hampsonii” by phenotypic and molecular methods. 17 The different species of Brachyspira have different virulence; for example, B. hyodysenteriae causes swine dysentery, a severe intestinal disease, whereas B. pilosicoli causes more mild disease, and B. innocens and B. murdochii are considered nonpathogenic. 48 In addition, B. pilosicoli is a potentially zoonotic pathogen. 34,70 Therefore, it is important not only to detect Brachyspira spp in diagnostic specimens but also to identify them accurately to species level. Both tasks are difficult phenotypically, since culture of Brachyspira requires strict anaerobic conditions and is time-consuming and labor-intensive. Ten years ago, a PCR–restriction fragment length polymorphism (PCR-RFLP) assay was developed to amplify the nicotinamide adenine dinucleotide (NADH) oxidase gene (nox) and generate species-specific restriction patterns using the restriction endonucleases BfmI and DpnII. 54 Applying this assay to 2050 Brachyspira isolates from pigs allowed identification and differentiation of B. hyodysenteriae, B. innocens, B. intermedia, B. murdochii, B. pilosicoli, and B. suanatina. 53 It remains to be determined whether this assay can identify the newly described B. hampsonii. Since DNA sequencing is becoming easier and inexpensive, this PCR-RFLP assay can be adapted as a PCR-sequencing assay, which may generate additional polymorphism information and thus be more accurate and potentially of greater value. For rapid identification, the assay could also be adapted as a real-time PCR assay using species-specific probes, which could replace the time-consuming RFLP. In the past few years, several real-time PCR assays have been published for the detection of Brachyspira from animal samples. For example, a multiplex PCR with specific probes simultaneously identified L. intracellularis, B. pilosicoli, and B. hyodysenteriae with a detection limit of less than 1000 cells/g of pig fecal samples. 71 Compared with gel-based PCR, real-time PCR assays can be more sensitive 1 in addition to being rapid and quantitative. 71
Another example of the use of PCR to identify difficult-to-culture bacteria is molecular detection of Mycoplasma species. Mycoplasma are either nonculturable (eg, hemotropic mycoplasma) or take days to isolate and often are overgrown by other bacteria in the samples. PCR assays have played important roles in the detection of Mycoplasma spp. For example, by using a procedure with automatic DNA extraction and a high-throughput real-time PCR instrument integrated with a laboratory information management system (LIMS), we were able to use a Mycoplasma iowae real-time PCR 13 to test tens of thousands of cloacal swabs within a short period of time in an eradication project recently. The sample volume was far beyond the capacity of traditional mycoplasma culture, not to mention that culture of M. iowae from cloacal swabs is usually unsuccessful because of overgrowth of other bacteria.
A PCR assay for detection of S. equi subsp equi in the diagnosis of strangles in horses is another valuable diagnostic tool. Although the organism is readily culturable, S. equi subsp equi can be difficult to phenotypically differentiate from other β-hemolytic streptococci, notably S. equi subsp zooepidemicus. A PCR assay targeting the gene for the virulence factor SeM of S. equi subsp equi has been demonstrated to be specific 62 and has been very useful for characterizing isolates as well as for detection of the pathogen directly from diagnostic specimens. Furthermore, sequence analysis of the SeM gene and the other virulence factor gene SzP can be used to distinguish wild-type S. equi subsp equi from a modified live vaccine strain. 33
Culture of Coxiella and Chlamydophila requires the use of a Biosafety Level 3 laboratory, but PCR detection can be performed in a Level 2 laboratory. Our laboratory has offered PCR detection of C. burnetii and Chlamydophila for several years. Recently, we identified C. burnetii by PCR in 69% and 75% of sheep and goat abortion submissions, respectively. By combining this information with knowledge of histologic lesions and other clinical and laboratory data, Coxiella was considered to be a cause of the abortion in 10% and 21% of the sheep and goat cases that tested positive, respectively. 26 Similarly, we identified C. abortus in 26% and 59% of sheep and goat abortion submissions, respectively, and this pathogen was considered significant in 38% and 63% of sheep and goat submissions that tested positive, respectively. Using a quantitative real-time PCR approach, we found that only in infections with a high bacterial load of C. burnetii or C. abortus could these agents be diagnosed as the cause of small ruminant abortion. 26 Similarly, a recent review summarized the progress of Clostridium difficile diagnosis and pointed out that detection of the bacteria in a clinical specimen may or may not be diagnostic of the disease. 11 These examples highlight that, because of its sensitivity, a positive PCR result by itself may not indicate a disease and that combination of quantitative PCR with other clinical and laboratory findings is critical in PCR-based disease diagnosis.
PCR assays are also useful when fresh tissue is not available for culture. For example, enteropathogenic Escherichia coli and Enterococcus spp infections have a similar histologic appearance, with bacteria adherent to the apical surface of enterocytes of the small intestinal villi. Although a presumptive identification of Enterococcus sp is possible by Gram-positive staining of tissue, PCR amplification with confirmation by fluorescent in situ hybridization (discussed further below) was used for specific identification as Enterococcus hirae. 43 DNA extracted from formalin-fixed, paraffin-embedded (FFPE) tissue is also useful in retrospective studies. Mycoplasma bovis DNA was identified in archived FFPE tissue, confirming that lesions of caseonecrotic bronchopneumonia caused by this pathogen were present but unrecognized 25 years ago. 25
Quality Control in Molecular Diagnosis
The wide availability of PCR equipment and methodology, the ease of designing new primers, and the ability to run multiple assays on a single sample create a situation in which rapidly implemented tests could yield spurious results. In practice, veterinary practitioners and pathologists tend to accept the results of unfamiliar laboratory tests with little question, even though it is a well-accepted principle that laboratory data must be interpreted in the context of the clinical and pathologic findings. Thus, a newly designed but flawed molecular assay can easily result in false clinical diagnoses, and quality control both during test development and in routine use is therefore critical.
One of the essential points is to avoid false-negative tests, which can be caused by PCR inhibitors in DNA samples. There are commercially available internal control kits for detection of inhibitors. Alternatively, as an internal control, DNA and primers for a reference gene can be added to the PCR reaction reagent mix, along with sample DNA and primers for the target gene. Failure to adequately amplify the internal control DNA indicates the possible existence of PCR inhibitors.
When inhibitors are suspected, serial dilution of the sample DNA extract usually can reduce the inhibitory effects. 52 Similar to a multiplex PCR, amplification of the internal control can compete with that of the target DNA and reduce the assay sensitivity. One solution is to test a 1- or 2-log dilution of the DNA sample directly, without amplification of the internal control. With availability of high-throughput PCR thermocyclers, this approach can actually save time and reagent costs. Our laboratory detects Geomyces destructans, the agent of white nose syndrome of bats, using a TaqMan real-time PCR assay. 40 Although no PCR inhibitors were found in bat skin samples, 40 we did find inhibitors in soil samples. We found that use of a commercial internal control kit reduced the PCR sensitivity by about 1 log. By serially diluting the DNA template (eg, 10 and 100 times), it can help to identify the existence of PCR inhibitors (undiluted samples were negative, whereas diluted samples were positive in the assay); this approach may have better sensitivity than using an internal control kit (Table 1). We found that some commercial DNA extraction kits (such as the Ultraclean Soil DNA kit; MoBio Laboratories, Carlsbad, CA) are more effective than others in removing PCR inhibitors from soil samples. The occurrence, properties, and removal of PCR inhibitors have been reviewed in detail. 56 These experiences indicate some of the care required to optimize PCR identification for particular samples and circumstances. False-positive PCR tests can have a major impact by leading to unnecessary or inappropriate therapy or control efforts, by jeopardizing animal movement and trade, and by incorrectly diagnosing diseases that have public health significance.
Detecting the Existence of PCR Inhibitors by Using an IPC or by Diluting DNA Extracts.
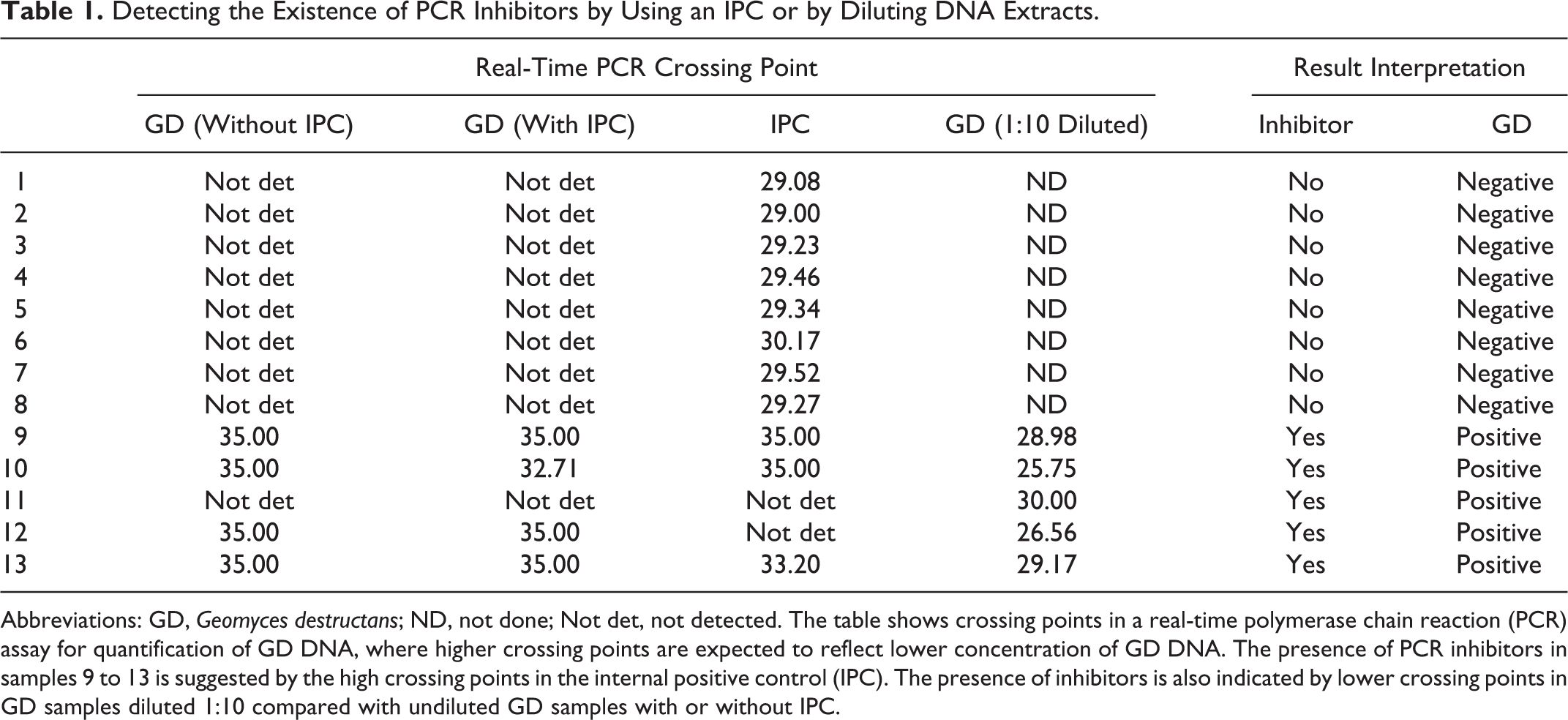
Abbreviations: GD, Geomyces destructans; ND, not done; Not det, not detected. The table shows crossing points in a real-time polymerase chain reaction (PCR) assay for quantification of GD DNA, where higher crossing points are expected to reflect lower concentration of GD DNA. The presence of PCR inhibitors in samples 9 to 13 is suggested by the high crossing points in the internal positive control (IPC). The presence of inhibitors is also indicated by lower crossing points in GD samples diluted 1:10 compared with undiluted GD samples with or without IPC.
Inadequate quality of extracted DNA is another potential cause of false-negative tests. Concentration and purity of extracted DNA is routinely estimated by the ratio of absorbances at 260 and 280 nm. Amplification of a reference gene, as described above, is also useful as an alert to poor sample quality. Negative controls routinely include both an absence of DNA in the PCR reaction and DNA extraction from a negative sample. Primer specificity is tested by analysis of melting curves of the real-time PCR product and by sequencing of the amplified product.
Another critical quality control point is to avoid false-positive reactions, which are usually caused by cross-contamination between samples or from previous PCR products. There have been many reviews outlining the various procedures required to prevent and detect false positives, which include using separate rooms for different steps of the reaction, including negative controls at every PCR step, and regular decontamination of working areas with ultraviolet light or bleach. 7,10,37,38
Use of Formalin-Fixed Tissues for PCR Assays
FFPE tissues can be used for PCR assays and 16S ribosomal RNA (rRNA) gene sequencing. However, since formalin treatment can fragment DNA, assays should be designed to work with smaller fragments. Specifically, it has been found that PCR worked well with 400–base pair (bp) fragment targets, but it was difficult to amplify 600-bp fragments when extracting DNA from FFPE tissue with commercial kits. 29 A recent study comparing 10 different extraction protocols and 2 commercial kits found the QIAamp DNA FFPE Tissue Kit (Qiagen, Valencia, CA) was most effective, being able to extract DNA and amplify 122- to 171-bp fragments from tissues that had been achieved for almost 200 years. 45 If cost is a concern, a manual procedure with protein K digestion may be used, since this method was shown to have results similar to the QIAamp DNA FFPE Tissue Kit for PCR-based amplification of 208- to 390-bp fragments but was more cost-effective. 65
In our laboratory, we compared DNA and RNA quality for PCR assays on FFPE vs frozen tissues using the QIAamp DNA FFPE Tissue Kit (Qiagen) and RNeasy FFPE Kit (Qiagen) for C. psittaci real-time PCR (78-bp target) and bovine viral diarrhea viruses type 2 (BVDV-2) RNA virus and porcine reproductive and respiratory syndrome virus (PRRSV) real-time RT-PCR using commercial PCR kits (fragment size unknown). Similar quantitative results were obtained for FFPE and frozen tissue samples (unpublished data). Other studies have found reduced sensitivity using FFPE compared with fresh tissue for a gel-based PCR assay to detect Helicobacter spp in feline tissues, and these differences in sensitivity affected the outcome of the study with respect to tissue distribution of the pathogen. 59 However, the assay sensitivity presumably depended on the DNA extraction method used. Using FFPE samples, we routinely perform 16S rRNA gene sequencing (350–500 bp) to detect an avian Coxiella-like organism and use PCR to detect bovine papillomavirus (122-bp target) with procedures described recently. 5,67
Cross-contamination of FFPE from other cases in the postmortem room, as well as from processing and sectioning tissues for histopathology, is a concern for molecular testing of FFPE tissue samples. As an example, Bartonella DNA contamination was found from cleaned microtome blades and other parts of the microtome, on the postmortem tissue trimming board (after cleaning), and in used paraffin from the tissue processor. 68 These findings suggest that, at least for PCR-based detection of Bartonella sp, it would be difficult to conclusively rule out cross-contamination in routinely processed blocks of FFPE tissue from diagnostic cases. Confirming the findings by an alternative method such as immunohistochemistry, or using a method to localize nucleic acid within tissues such as in situ hybridization, could be used to rule out cross-contamination as a reason for positive PCR results on FFPE tissues.
Multiplex PCR Assays
In general, the use of multiplex PCR assays, which identify more than 2 pathogens, toxin genes, or other targets, can reduce both the cost and time to obtain results. For example, by charging a little more than for a regular single PCR assay, our laboratory offers an assay detecting multiple virulence factors of enterotoxigenic E. coli (ETEC) using a multiplex gel-based PCR that is highly specific and sensitive. 16 Another gel-based multiplex PCR assay designed to simultaneously detect B. hyodysenteriae, B. pilosicoli, and L. intracellularis had substantial to almost perfect agreement with a nested PCR assay for Brachyspira and moderate agreement with the immunofluorescence test for Lawsonia. 42
In addition to a gel-based platform, multiplex PCR can be done in a real-time PCR format, with determination of melting temperature to differentiate multiple targets. There are only a few assays developed on this platform for animal pathogen detection. For example, a real-time multiplex PCR was developed to screen for common Salmonella serotypes by measuring the melting temperature of the PCR amplicons with SYBR Green, a double-stranded DNA dye. 49 Another multiplex real-time PCR based on the same principle was described for the detection of porcine circovirus 2, porcine parvovirus, pseudorabies virus, and Torque teno sus viruses 1 and 2 in pigs. 46 Compared with gel-based PCR, this type of assay is fast and relatively inexpensive since gel electrophoresis is not involved. Although the specificity of the real-time PCR can be improved by using high-resolution melt analysis (using a second-generation dye to detect double-stranded DNA), 50,64 in general, it is recognized that using specific probes rather than double-stranded DNA dyes makes a real-time PCR assay more specific. A good example is a panel of 2 probe-based multiplex real-time PCR assays for rapid and simultaneous detection of the 5 major bovine enteric pathogens: bovine coronavirus, group A bovine rotavirus, Salmonella spp, E. coli K99(+), and Cryptosporidium parvum. This assay is highly sensitive with a detection limit of 0.1 50% tissue culture infective dose (TCID)50 for viruses, less than 5 colony-forming units (CFU) for bacteria, and 50 oocysts for Cryptosporidium. 18 Another example is a real-time PCR with different sets of primers and probes for the detection of Chlamydia spp, C. burnetii, and Neospora caninum in ruminant abortion samples. 51 Instead of using multiple set of primers, a multiplex real-time PCR assay can use 1 set of universal primers and multiple specific probes to identify different targets. For example, a PCR assay was able to amplify all ETEC with 1 set of primers and identify F4 (K88), F4ab, F4ac, and F4ad variants with 3 probes. 12 One should be aware that when there are mixed infections, the universal primers can amplify the abundant bacteria and result in false-negative results for the bacteria present in scanty numbers. For example, we found that a M. bovis real-time PCR assay, which amplifies 16S rRNA gene with universal primers and detects M. bovis with specific probes, 14 can generate false-negative results from mastitic milk samples with mixed infection of M. bovis and high numbers of Staphylococcus aureus (unpublished data). Development of additional sensitive and validated multiplex real-time PCR assays for the detection of multiple pathogens in species-specific clinical syndromes would be welcomed by veterinary practitioners.
DNA Microarray
Multiplex PCR for detecting multiple bacterial pathogens has not been widely used in animal health diagnostic laboratories, possibly because these assays are difficult to optimize and fully validate. DNA microarray is a more advanced tool for detection of multiple pathogens. Probes for multiple targets are anchored on a solid surface such as a glass slide or a microcentrifuge tube to form a solid microarray, or on microbeads to form a liquid array. Specific DNA from samples is amplified and labeled by PCR with universal primers or by multiplex PCR and then is added to the microarray or liquid array for hybridization. After washing, the hybrids of positive PCR amplicons and probes display labeled fluorescence or chemiluminescence. A rare example in the animal health arena is a microarray that consists of probes for 55 different bacteria developed for the identification of Mycoplasma spp, including Mycoplasma mycoides subsp mycoides. 63 The current form of the DNA microarray requires PCR and hybridization and is therefore expensive and relatively time-consuming. The technique has not been applied in routine veterinary diagnosis. However, a microarray may be adapted to a portable device for routine or even pen-side diagnosis when manufacturing costs and the size of the device are reduced. The relatively small size of the animal health compared with the human medical diagnostic market may be one factor hindering the development of commercially available diagnostic microarrays.
Fluorescent In Situ Hybridization
Fluorescent in situ hybridization (FISH) is a well-established non–culture-based procedure for identifying bacteria that uses fluorescent oligonucleotide probes based on 16S rRNA sequences specific to the species, genus, or other phylogenetic level of interest. It is rapid and specific and has had particular application as a research tool to visualize specific and often hard-to-identify bacteria within complex mixtures of other bacteria; FISH combines the precision of a molecular tool with the ability to visualize bacteria within their broader microbial ecological context. Although the procedure is straightforward, processing is labor-intensive and the probes are relatively expensive. An excellent review of the technique is available. 8
FISH can be used directly on clinical samples or on FFPE. The protocols for FISH typically involves 4 steps: (1) sample fixation and permeabilization (to allow penetration of the fluorescent probes into the cell and their protection against endogenous RNases), (2) hybridization with fluorescein-labeled 15- to 30-bp oligonucleotide probes under stringent conditions, (3) washing to remove unbound probe, and (4) mounting, visualization, and photographic analysis of the probed materials, often using antifading materials to reduce loss of fluorescence.
While the FISH procedure is straightforward, it must be optimized for different samples and probes. For example, methods for optimal attachment of bacteria to glass surfaces may need to be compared and Gram-positive bacteria may require treatment with lysozyme or lysostaphin to remove cell-wall peptidoglycan that would otherwise interfere with probe penetration into the cell. Peptide nucleic acid probes, although expensive, are often used to overcome problems associated with probe penetration, since their lack of charge prevents electrostatic repulsion by the negatively charged sugar-phosphate backbone of DNA. Depending on optimization, different probes labeled with different fluoresceins can be used to identify bacteria at several different levels within bacteria populations, such as the bacterial kingdom, order, family, genus, or species, depending on the region of 16S rRNA on which the probe is based. The choice of the probe will depend on the availability of a sequence that is discriminatory for the bacterium of interest.
Model applications of the use of FISH in veterinary diagnosis include direct identification of Flavobacterium psychrophilum, 60 spirochetes in bovine digital dermatitis, 15 canine and feline Helicobacter spp, 30 feline enteroadherent E. hirae, 44 adherent and invasive E. coli, and other bacteria in murine models of Crohn disease, 20 bacterial genera on the equine gastric mucosa, 47 and bacteria in archived FFPE cases of canine endocarditis. 31 As noted, these applications have largely been focused on research questions, but there is obvious potential for FISH in rapid diagnosis in veterinary laboratories working with difficult-to-culture bacteria in complex microbial specimens, notably the intestine.
Universal DNA Sequencing and Microbiome Profiling
With specific PCR assays or DNA microarrays, one can detect only previously described pathogens, and these assays may therefore fail to identify novel or emerging pathogens. Using universal primers to amplify and sequence genes in all bacterial species can overcome this drawback. Under some conditions, this technique can be applied directly to tissue samples to identify nonculturable bacterial pathogens. The most commonly used universal gene for bacterial identification is the 16S rRNA gene. Although far less commonly used, alternatives to the 16S rRNA gene have been suggested, which include the genes for chaperonin-60 (cpn60); 35 recombination and repair protein (recN) and DNA polymerase III subunits γ, τ (dnaX); 77 and the β-subunit of RNA polymerase (rpoB) and esterase (est). 39 Since the 1980s, sequencing of the 16S rRNA gene has been used as an important tool for phylogenetic analysis and classification of bacteria. 32 The 16S rRNA gene contains regions that are well conserved in all organisms, making this ideal for primer design, PCR-based sequencing, and sequence alignment. It is possible to design universal primers for most bacteria. The gene also contains species-specific variable regions that can allow species identification.
When a sample is suspected to contain an nonculturable bacterial pathogen, universal primers can be used for PCR amplification and sequencing of the 16S rRNA gene. The unknown bacteria can be identified by comparing its 16S rRNA gene sequences with those in a database such as GenBank (http://www.ncbi.nlm.nih.gov/). For example, using the 16S rRNA gene sequencing approach, Coxiella-like bacteria were identified from psittacine birds, toucans, and lorikeets, which had disease resembling that caused by C. burnetii. The samples were negative using a PCR assay specific for C. burnetii but had 16S rRNA gene sequences with about 98% similarity to that of C. burnetii. 58,67,72
One needs to keep in mind that the public database of 16S rRNA gene sequences is not complete. Although 16S rRNA gene sequences are available for most bacterial-type strains that can be sequenced, some are still missing. 76 In addition, 16S rRNA gene sequence analysis can identify many species only to the genus level, so that sequencing other genes (or use of PCR specific for these genes) would be necessary for identification to the species level.
Universal gene sequencing is reliable only with samples containing a single bacterial species, since multiple bacteria will lead to a mixed DNA sequence chromatogram that cannot be analyzed. There is also a well-known bias in the relative amplification efficiency of 16S rRNA from heterogeneous samples. 61 This is a major limitation for the use of the technique on tissue samples, so this approach is used mainly for identification of cultured bacteria. However, another study used cloning of PCR products and software to resolve mixed chromatograms to identify mixed bacterial infections in blood samples, 73 illustrating the potential to overcome the difficulties of identifying polymicrobial infections.
An alternative method to contend with this problem with universal gene sequencing uses high-throughput 16S rRNA gene sequencing for identification of a whole community of microorganisms. The technique and its application to food animal microbiome profiling has been reviewed. 27 The advantage of high-throughput 16S rRNA gene sequencing is that it can identify polymicrobial infections, since it can amplify all microbial nucleic acid sequences with only limited bias.
Compared with PCR assays and DNA sequencing, massive parallel DNA sequencing requires a higher quality of nucleic acid. However, there have been reports of the successful use of FFPE tumor tissue samples for massive DNA sequencing. 3,57,74 For pretreatment of FFPE tissues for genomic studies, methods have been described that repair gaps in degraded DNA, ligate short DNA fragments, and remove chemical adducts. 28 Most recently, the full genome of the 1918 influenza virus and the associated bacteria that caused pneumonia were determined by massive DNA sequencing using RNA and DNA extracts from FFPE lung tissue samples, 75 suggesting that application of such methods for veterinary diagnosis and research may be technically feasible.
We foresee that in the coming years, high-throughput 16S rRNA gene sequencing, together with other high-throughput DNA sequencing applications, will be used more and more for identification of new and nonculturable bacterial pathogens. Similar to other types of high-throughput DNA sequencing, the disadvantages of high-throughput 16S rRNA gene sequencing include the expense, complicated data analysis, and relatively long sample preparation and sequencing procedures. However, as the instruments become more affordable, operating costs decline, and analytical software becomes easier to use, the technique will become more accessible to veterinary diagnostic laboratories. For example, a rapid (same day) and inexpensive procedure for routine clinical use was recently described. 55 In this study, multiplexing massive DNA sequencing was performed on sputum samples from patients with cystic fibrosis, using primers designed to target the variable regions 1 and 2 (V1–V2) of the 16S rRNA gene sequences at a cost of about US$60 per sample. Massive DNA sequencing can amplify 16S rRNA gene fragments for all bacteria species in the samples without bias and therefore can identify all mixed infection simultaneously. In general, the procedures described in this study are feasible for animal health laboratories in terms of turnaround time and cost.
Molecular Typing of Bacterial Pathogens Directly From Samples
Epidemiological analysis often requires molecular typing of bacterial isolates for comparison within and between outbreaks. For nonculturable or difficult-to-culture pathogens, a molecular typing assay must be effective on DNA samples extracted directly from clinical samples or tissues. Recently, multiple-locus-variable number of tandem repeats analysis (MLVA) was used for molecular typing of Bordetella pertussis directly from clinical specimens. 36 We have also used MLVA to perform molecular typing of C. burnetii DNA extracts from abortion samples (unpublished data, Shu Chen, University of Guelph). MLVA is robust and the results are “portable” (reproducible between laboratories), but it requires setting up PCR reactions for specific bacterial species and may generate artifacts in typing profiles because of PCR inaccuracy. 66
As the cost of high-throughput DNA sequencing becomes ever lower, comparing the genomes of bacterial isolates may be done for routine diagnosis in the near future. This technique has been used in numerous instances for genotyping of pure bacterial cultures. Very recently, the potential of using metagenomic data for bacterial typing directly from diagnostic specimens was illustrated by sequencing multiple bacterial genomes, including Chlamydia trachomatis, directly from vaginal swabs. 2
Identification of Antimicrobial Resistance Using Massive DNA Sequencing
In the past, it was impossible to determine the antimicrobial resistance of bacteria without isolation in culture. With whole-genome sequencing, genes for antimicrobial resistance as well as regulatory genes can be sequenced and identified. It is likely that in the coming years, high-throughput DNA sequencing directly from diagnostic specimens may be used to identify not only the infectious pathogens but also their antimicrobial resistance genes. Given the continuous evolution of resistance genes, there will, however, always be a need to establish the resistance phenotype, in addition to genotype, but for well-established resistance genes, massive DNA sequencing may have value.
Matrix-Assisted Laser Desorption/Ionization Mass Spectroscopy
Recently, matrix-assisted laser desorption/ionization–time-of-flight mass spectroscopy (MALDI-TOF MS) has been widely introduced as a rapid tool for the identification of bacterial isolates directly from cultures. 19 It has become a paradigm-changing technology for diagnostic bacteriology. It is rapid, accurate, more cost-effective than current phenotypic approaches to identification (despite the high initial cost of the machine), and simple to use. 19 Bacteria are identified literally within minutes of sample placement into the machine.
Commercial MALDI-TOF MS systems identify bacteria to genus and to species by comparing the MALDI-TOF MS spectra of ribosomal proteins, although mass spectra of other proteins of type organisms may be included in the usually proprietary identification software provided by the manufacturers. 19 The main spectra can often be expanded “in house,” since there is still considerable scope to add spectra of relatively unusual animal bacterial pathogens. MALDI-TOF MS offers a robust and rapid method of identifying bacteria, without even doing a Gram stain; most reports of errors in identification relate to inadequate databases. 19 Although it works well for routine bacterial speciation from pure cultures, we have found that it is difficult to identify Mycoplasma species from animals. Protocols for sample preparation and a database to cover such types of veterinary pathogens are needed, and their spectra should be included in the manufacturer’s software. Although it is said to be possible to use MALDI-TOF MS for bacterial strain typing, 4,6,41 we have found that it has relatively low discrimination, which may be improved by using a more polymorphic spectrum.
Although MALDI-TOF MS identification is currently done almost exclusively on bacterial colonies and is therefore largely outside the scope of this review, the technique can potentially be used for identifying bacterial pathogens directly from diagnostic samples. There are reports of the use of MALDI-TOF MS to detect bacterial pathogens directly in blood broth culture samples 9 and also of accurate identification of bacteria directly from human urine after removal of leukocytes by centrifugation. 23,69 We anticipate further application to direct identification of bacteria in clinical samples.
Conclusion
Currently available molecular tests, including PCR and genetic sequencing methods, have made it possible to identify nonculturable bacterial pathogens. PCR-based typing methods such as MLVA are potentially useful for molecular typing of bacterial DNA obtained directly from diagnostic specimens. As the cost becomes more affordable and the procedures become easier, massive DNA sequencing may eventually become a routine assay for identification and characterization of nonculturable bacterial pathogens. A critical issue and major challenge is the validation of the technologies to establish the sensitivity and specificity of novel techniques against pathological and clinical samples of known causation, as well as the identification of new “gold standards” based on molecular diagnosis rather than infectious agent isolation. Astute veterinary laboratory diagnosticians remain critical in the face of the seductive Aladdin’s cave of emerging molecular diagnostic tools.
Footnotes
Acknowledgements
We thank Patricia Bell-Rogers, Colin Cameron, Jason Eidt, Rebeccah McDowall, Ana Rita Rebelo, and Joanna Sawicki for their technical assistance with some of the studies noted in this review.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported in part by the OMAFRA-UG Agreement through the Animal Health Strategic Investment fund (AHSI) managed by the Animal Health Laboratory of the University of Guelph.