
Abstract
An outbreak of eastern equine encephalomyelitis (EEE) occurred in Michigan free-ranging white-tailed deer (Odocoileus virginianus) during late summer and fall of 2005. Brain tissue from 7 deer with EEE, as confirmed by reverse transcriptase polymerase chain reaction, was studied. Detailed microscopic examination, indirect immunohistochemistry (IHC), and in situ hybridization (ISH) were used to characterize the lesions and distribution of the EEE virus within the brain. The main lesion in all 7 deer was a polioencephalomyelitis with leptomeningitis, which was more prominent within the cerebral cortex, thalamus, hypothalamus, and brainstem. In 3 deer, multifocal microhemorrhages surrounded smaller vessels with or without perivascular cuffing, although vasculitis was not observed. Neuronal necrosis, associated with perineuronal satellitosis and neutrophilic neuronophagia, was most prominent in the thalamus and the brainstem. Positive IHC labeling was mainly observed in the perikaryon, axons, and dendrites of necrotic and intact neurons and, to a much lesser degree, in glial cells, a few neutrophils in the thalamus and the brainstem, and occasionally the cerebral cortex of the 7 deer. There was minimal IHC-based labeling in the cerebellum and hippocampus. ISH labeling was exclusively observed in the cytoplasm of neurons, with a distribution similar to IHC-positive neurons. Neurons positive by IHC and ISH were most prominent in the thalamus and brainstem. The neuropathology of EEE in deer is compared with other species. Based on our findings, EEE has to be considered a differential diagnosis for neurologic disease and meningoencephalitis in white-tailed deer.
Eastern equine encephalomyelitis (EEE) virus (EEEV) is a zoonotic alphavirus spread by mosquitoes that affects humans, horses, and a variety of avian and mammalian species. 3,4,8,12,16 This report describes the neuropathology of EEEV infection in free-ranging white-tailed deer (Odocoileus virginianus), including viral antigen and nucleic acid distribution. The 7 deer included in this study were part of an outbreak of EEE in Michigan deer that occurred during the late summer and fall of 2005 and for which the detailed epidemiology and a brief description of the gross and histopathologic and immunohistochemical findings have been described previously. 18 The disease also had been reported in a single white-tailed deer. 20
Between August and October 2005, an outbreak of a neurologic disease, affecting at least 22 free-ranging white-tailed deer in southwestern Michigan, was reported as part of the ongoing chronic wasting disease (CWD) survey in Michigan. 18 Clinical signs observed in affected deer included ataxia, head tilt, circling, lack of wariness to humans, drooling, poor body condition, poor hide quality, and hair loss. Necropsies were conducted by Department of Natural Resources personnel under biosafety level 3 conditions. Affected deer had no gross lesions, and brains were submitted to the Diagnostic Center for Population and Animal Health for further diagnostic investigation.
Materials and Methods
Formalin-fixed, paraffin-embedded (FFPE) brain tissue from 7 deer with EEE, as confirmed by reverse transcriptase polymerase chain reaction (RT-PCR) and nucleic acid sequencing using fresh tissue, was further evaluated in the current study. Multiple cross sections of cerebral cortex, cerebellum, diencephalon, and brainstem, including medulla oblongata, were routinely processed, paraffin embedded, sectioned at 5 μm, and stained with hematoxylin and eosin.
Additional RT-PCR assays were performed on paraffin-embedded material from all 7 deer, using a previously described SYBR Green–based real-time RT-PCR, to confirm positivity of the individual blocks. 7,15 Prior to RNA extraction, approximately 25 mg of tissue was deparaffinized in 1 ml of Citrisolv (Fisher Scientific International, Hampton, NH), then washed twice in 1 ml of 100% ethanol and air-dried for 10 minutes. Total RNA was extracted using the RNeasy Mini Kit (QIAGEN, Valencia, CA) and eluted in 50 μl of RNAse-free water. RT-PCR was performed with an IQ5 real-time PCR detection system (Bio-Rad Laboratories, Hercules, CA), using the Quantitect SYBR Green RT-PCR Kit (QIAGEN). 15
Indirect immunohistochemistry (IHC) and in situ hybridization (ISH) were performed on sections of brain from all 7 deer on BenchMark and Discovery Automated Staining Systems (Ventana Medical Systems, Tucson, AZ), respectively. 3,8,12,14,15 For IHC, polyclonal mouse ascites fluid, recognizing the 88–amino acid and about 50-kDa E1 structural protein of the EEE alphavirus, was used as the primary antibody at a concentration of 1:500 for 30 minutes. 3 A multimer-based technology detection system (ultraView; Ventana Medical Systems) and bulk buffers specifically designed for use on the BenchMark Automated Staining System (Ventana Medical Systems) were used for immunolabeling and visualization. Brown chromogen (3,3′-diaminobenzidine [DAB]) was used for single IHC, whereas red chromogen (Alkaline Phosphatase Red) was used when IHC was combined with ISH. 15 Antigen retrieval was performed using Protease 3 (Ventana Medical Systems). 3,14,15 The slides were counterstained with Ventana hematoxylin (Ventana Medical Systems). 15
For ISH, the EEE probe consisted of a 5′-digoxygenin–labeled, high-performance liquid chromatography–purified (Integrated DNA Technologies, Coralville, IA) 40–base pair DNA sequence, 5′-ACGATGTGGTCTTTCGGTGGCTTACAATC TCCTTTGCAGG-3′, complementary to a conserved region of the virus. 6,15 Proteolytic treatment was performed using Protease 3. Hybridized viral RNA was detected using an anti-digoxigenin antibody (Invitrogen Corporation, Frederick, MD) and a streptavidin-alkaline phosphatase detection system (UMap anti-Rb AP) with a BlueMap NBT/BCIP substrate kit (Ventana Medical Systems). 15 Positive controls included brain tissues from horses affected with EEE, as confirmed by RT-PCR. 15 Brain tissues from white-tailed deer with no evidence of viral encephalitis, which had tested negative by RT-PCR for EEE, and brain tissues from horses that were positive for West Nile virus (WNV) and negative for EEE by RT-PCR were used as negative controls. 15 For negative reagent controls, the primary antibody or the oligoprobe was replaced by homologous nonimmune sera for IHC and a probe for WNV for ISH, respectively.
Results
All blocks included in the study tested positive by RT-PCR for EEE on shavings from FFPE tissues. One of the 7 deer also was positive for WNV by RT-PCR and nucleic acid sequencing on fresh tissue, 18 but RT-PCR on FFPE material, as well as IHC and ISH for WNV, was all negative for this animal.
All 7 white-tailed deer had similar microscopic lesions. There was a mild to moderate polioencephalomyelitis with leptomeningitis, which was more prominent within the cerebral cortex, thalamus, hypothalamus, and brainstem. The cerebellum, choroid plexus, and ependyma were mostly spared. Scattered throughout the affected areas were variably intense inflammatory infiltrates within the Virchow-Robin spaces surrounding small- to medium-sized vessels (Fig. 1). Inflammatory infiltrates consisted predominantly of small lymphocytes with lower numbers of histiocytes. Occasional glial nodules were observed (Fig. 1). Neutrophils were also admixed within the predominantly mononuclear infiltrate in 5 of 7 cases, although in low numbers compared with the mononuclear cells. Three deer had multifocal microhemorrhages (Fig. 2) commonly surrounding smaller vessels with or without perivascular cuffing, although vasculitis was not observed. Within the neuropil, there was sporadic neuronal necrosis characterized by hypereosinophilic, shrunken neurons with pyknotic nuclei associated with perineuronal satellitosis and neutrophilic neuronophagia (Fig. 3). Occasionally, normal-appearing neurons were also surrounded by small lymphocytes, macrophages, and neutrophils. Neuronal necrosis was observed in all 7 deer and was most prominent in the thalamus and the brainstem, especially the pons. Neurons of the cerebral and cerebellar cortex, as well as the hypothalamus, were significantly less affected and often only exhibited mild chromatolysis. Cerebral and cerebellar leptomeninges multifocally contained moderate numbers of small lymphocytes, macrophages, and a few neutrophils. Less commonly observed microscopic lesions included moderate edema of Virchow-Robin spaces (5 of 7 cases), astrocyte hypertrophy (3 of 7 cases), microglial cell activation (3 of 7 cases), and multifocal glial nodules (1 of 7 cases).
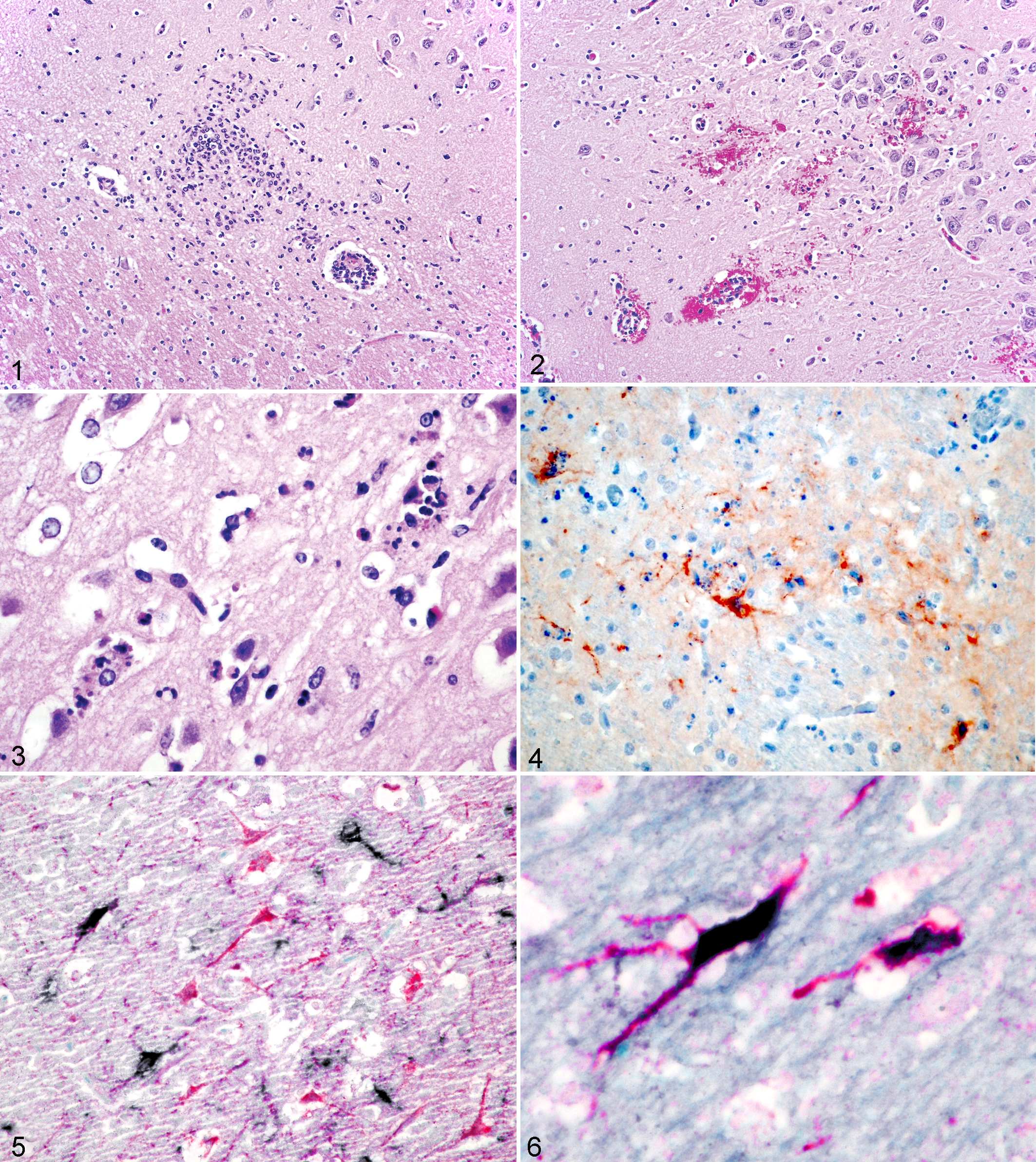
Positive IHC and ISH labeling was mainly observed in the thalamus and the brainstem, especially the pons, and to a lesser degree in the cerebral cortex of the 7 deer. There was minimal labeling in the cerebellum and hippocampus. EEEV-specific antigen was detected in the perikaryon and dendrites of necrotic and intact neurons and to a much lesser degree in glial cells and a few neutrophils (Fig. 4). Besides neurons, glial cells surrounding morphologically intact and necrotic neurons were most often positive. There was no intranuclear labeling, and not every necrotic neuron was labeled. Intracytoplasmic chromogen presented as finely granular, formed prominent clumps or diffusely labeled the whole cell body. Labeling commonly extended into axons and dendrites. Cells exhibiting positive labeling for viral nucleic acid were exclusively neurons. Positive ISH labeling was observed in the perikaryon of these neurons and extended along axons and dendrites, similar to the antigen detection by immunohistochemical labeling.
When using double labeling with IHC for viral antigen and ISH for viral RNA, dual-labeled neurons were most prominent in the thalamus and brainstem (Fig. 5). Although many neurons were positive by IHC and ISH (Fig. 6), only viral antigen was detected in other cell types. Furthermore, less than 50% of neurons positive by IHC were also positive by ISH. In contrast, less than 10% of ISH-positive neurons were negative by IHC.
Discussion
The distribution of microscopic lesions in the brain of deer naturally infected with EEE was similar to the pattern in advanced lesions of EEE observed in other species, including naturally infected horses and experimentally inoculated rodents and primates. 3,11,13,17,19,21 Neuronal necrosis was the predominant lesion and was most severe in the thalamus and pons, whereas inflammation was usually mild to moderate and characterized by mixed cell infiltrates that included neutrophils. The brainstem and thalamus have been some of the areas to consistently exhibit severe lesions of neuronal necrosis in many animal species infected by EEE, including deer. 1,3,11 –13,17,21 In contrast, microscopic brain lesions in humans and primates infected with EEE are more severe in the forebrain, especially the hippocampus. 1,11,19 Different routes of hematogenous versus neurogenic viral spread following infection of olfactory epithelial cells may account for this difference. 19
Vasculitis, as has been described for EEE infections in guinea pigs and macaques, was not a significant feature of EEE in deer. 17,19 Vasculitis has also been reported in a single case of naturally occurring EEE in a horse but was not detected in a more detailed IHC study of 2 horses. 3,5 We observed microhemorrhages in 3 deer, a lesion also reported in humans, primates, dogs, and hamsters. 1,4,11,13 Hemorrhages in hamsters were reportedly associated with vasculitis that was described as inflammatory cell infiltration into the vascular wall but otherwise lacked characteristic features such as fibrinoid necrosis. 13 We also did not detect viral antigen or nucleic acid in endothelial cells. Viral antigen, but not nuclei acid, was detected in neutrophils as has been previously reported in horses infected with EEE. 3 Similar to what we observed during our study, detection of viral nuclei acid has been limited to neurons in experimental studies in mice. 21 The inability to detect viral nucleic acid in neutrophils may simply reflect a lack of sensitivity of ISH to detect small amounts of virus or indicate an absence of replication of EEE virions in neutrophils and potential uptake of exogenous EEE virus antigen by neutrophils.
The absence of viral RNA in a large number of IHC-positive neurons may also be due to a higher likelihood of RNA degradation prior to tissue fixation or stronger cross-linking by prolonged fixation. This is especially a concern since the samples included in the study originated from deer that were euthanized or recovered in the field, and neither the time prior to tissue fixation nor the fixation time was consistent among these deer and significantly prolonged for some cases. Furthermore, the ISH probe detects genomic positive-sense viral RNA but should bind to generated messenger RNA. This would suggest that neurons at an earlier stage of EEE infection likely contain virions that are replicating at a high enough rate to meet the threshold for detection required by ISH. 15 In contrast, neurons in a later stage of replication contain large amounts of structural proteins in preparation for budding, which would also include more binding sites detectable by IHC. 15 The clinical presentation and the lesions in the brains of these deer indicate a more advanced stage of disease. This may explain the small number of ISH-positive neurons that were negative by IHC and most likely represent neurons in an early stage of infection, whereas the large number of neurons that were IHC positive and ISH negative may indicate neurons in a late stage of replication.
EEE in deer has been reported only in a single case prior to this cluster of cases. 18,20 Considering the emphasis on large-scale surveillance for CWD in many locations throughout North America over the past few years, it is not surprising that more cases of neurologic disease in cervids have been submitted for diagnostic pathologic examination. Michigan has experienced endemic EEE in birds, horses, and people for more than 60 years. 2 However, only in the past 10 years has large-scale ongoing surveillance for CWD been conducted in free-ranging deer in the state. This likely explains the detection of this localized outbreak of EEE in white-tailed deer. Interestingly, exposure of white-tailed deer to EEEV seems to be relatively common, and a recent study detected up to 19% serologic positive deer in some counties in Maine. 10 However, no viral nucleic acid was detected by RT-PCR in any of the brains collected. 10
Other important primary neurologic diseases affecting white-tailed deer include WNV, CWD, toxoplasmosis, bacterial meningitis or encephalitis, and meningeal worm infestation. 9,20 Each of these conditions may result in similar neurologic signs in deer, which include ataxia, head tilt, circling, blindness, recumbency, and loss of wariness of humans. 9,20 Although WNV can pose a significant diagnostic challenge based on microscopic examination only, the combination of routine histology with advanced molecular diagnostic testing such as RT-PCR, IHC, or ISH on sections of submitted brain can easily differentiate EEE from these diseases.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.