
Abstract
An adult male guinea pig (Cavia porcellus) with a 1-month history of hind limb paresis, torticollis, and seizures was euthanized and submitted for necropsy. Gross examination was unremarkable, but histologic examination revealed multifocal eosinophilic and lymphoplasmacytic choriomeningitis and cross sections of nematode parasites within the leptomeninges of the midbrain and diencephalon. Morphologic features of the nematode were consistent with a metastrongyle, and the parasite was identified as Parelaphostrongylus tenuis by polymerase chain reaction testing and nucleotide sequencing. Further questioning of the owner revealed that the guinea pig was fed grass from a yard often grazed by white-tailed deer (Odocoileus virginianus). To the authors’ knowledge, this is the first report of a naturally occurring P. tenuis infection in a guinea pig.
A 2.5-year-old male guinea pig (Cavia porcellus) was euthanized and submitted for necropsy following a 1-month history of progressive neurologic signs, including hind limb paresis, torticollis, and seizures. Treatment included calcium, vitamin C injections, prednisolone, and buprenorphine. The guinea pig was purchased from a pet store 2 years earlier and was fed a commercial diet supplemented with a variety of fruits and vegetables, including grass from a yard frequented by white-tailed deer (Odocoileus virginianus). For approximately 5 months prior to the onset of clinical signs, the guinea pig was housed with another healthy young male guinea pig. There was no history of trauma.
Differential diagnoses for neurologic disease in guinea pigs include trauma, pregnancy toxemia, hypocalcemia, and bacterial otitis/meningitis. Neurologic disease associated with Baylisascaris procyonis migration has been reported in guinea pigs housed on shavings contaminated with raccoon feces. 16 Although neoplastic disease is uncommon in guinea pigs, tumor metastases to the central nervous system (CNS) would be an important differential for an animal with multicentric neurologic deficits, as in this case.
A complete necropsy was performed. The calvarium was opened and the brain fixed in situ for sectioning. The entire vertebral column was fixed and decalcified, and representative cross sections were trimmed for histopathology. No gross lesions were detected during the necropsy; however, microscopic examination revealed cross sections of nematode parasites (Fig. 1) within the leptomeninges covering the midbrain, associated with eosinophilic and lymphoplasmacytic choriomeningitis (Fig. 2). The nematodes were 100 to 200 μm in diameter, with a thin, smooth cuticle; coelomyarian musculature; accessory hypodermal chords; and a large gastrointestinal tract composed of few multinucleate cells. Cross sections also contained a genital tract with either sperm or thin-shelled eggs. Multifocally, the leptomeninges of the brain and spinal cord and the choroid plexus were expanded by moderate infiltrates of eosinophils, macrophages (often containing brown cytoplasmic pigment), plasma cells, and fewer lymphocytes. In the cross section of the vertebral column at the level of T12, there were multifocal areas in all funiculi characterized by dilated axon sheathes and swollen axons. In the vertebral column section at the level of L3, the spinal cord was unremarkable, but within the epaxial and hypaxial skeletal muscles, there were multifocal areas of myofiber degeneration and a focal area of heterophilic myositis.
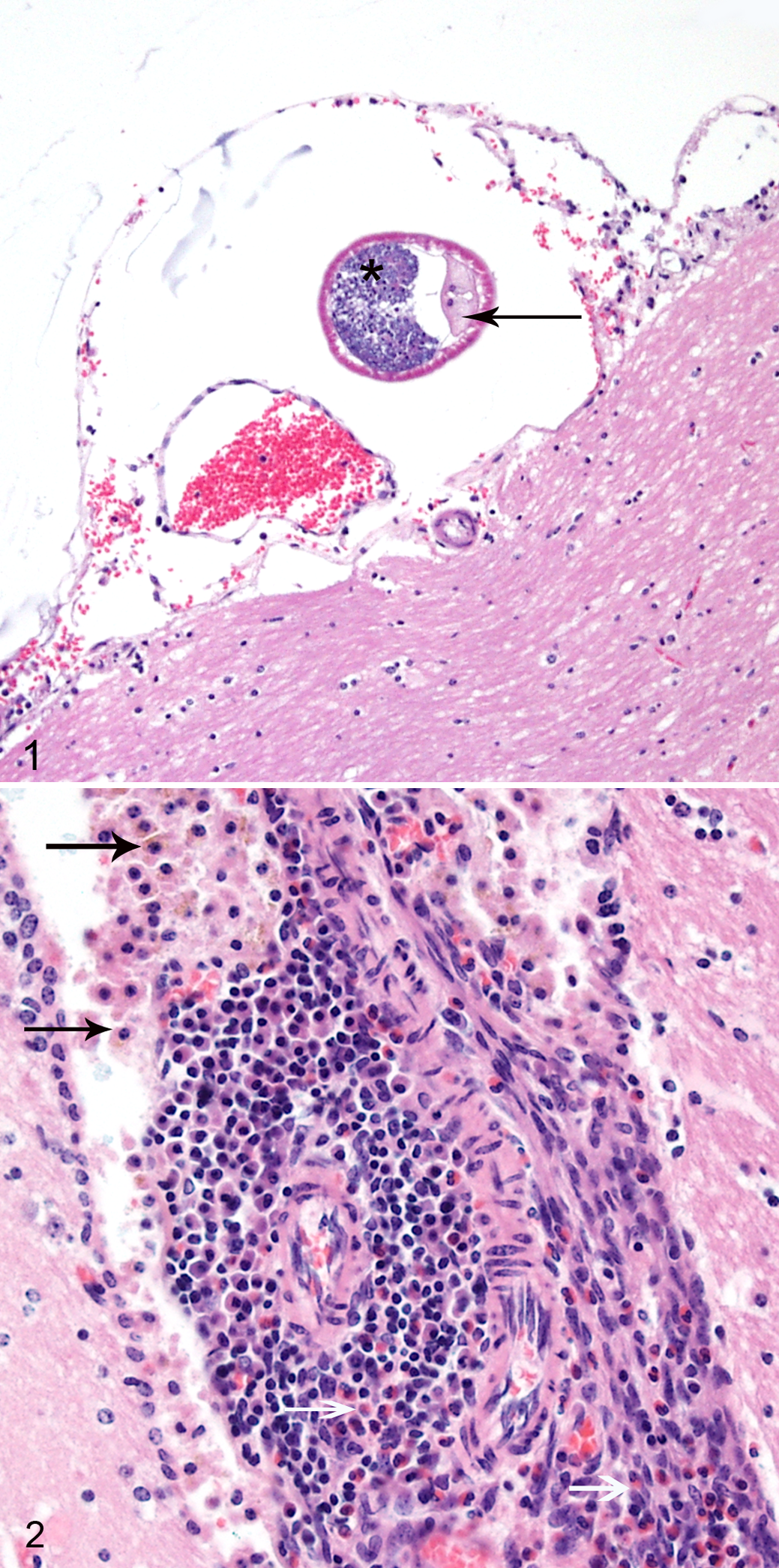
Based on histologic features, the nematode within this guinea pig’s leptomeninges was classified as a metastrongyle. The meningeal distribution of the parasite and the history of exposure to grass grazed by white-tailed deer in an area endemic for meningeal worm infection led to a tentative identification of the nematode as Parelaphostrongylus tenuis. Formalin-fixed paraffin-embedded sections of the midbrain and leptomeninges were submitted to the Center for Wildlife Health at the University of Tennessee for polymerase chain reaction (PCR) testing to confirm the nematode identification.
DNA was extracted from a thin section of the paraffin-embedded brain tissue using a DNeasy Blood and Tissue Kit (Qiagen, Inc, Valencia, CA) according to the manufacturer’s instructions. PCR was used to amplify a portion of the second internal transcribed spacer (ITS-2) region of the ribosomal RNA (rRNA) gene 5 . The Parelaphostrongylus spp primers included PTP1, 5′-(CCGTCGAATACATGTCATCC)-3′ and PTP2, 5′- (TCGTCAAGACGATGATTCCC)-3′, 4 which produce a PCR product of 228 bp. 10 PCR components and reaction cycling parameters were performed as previously described. 7 PCR products were visualized by electrophoresis in a 2% agarose gel, excised, and purified using a QIAquick Gel Extraction kit (Qiagen, Inc) according to the manufacturer’s instructions. Sequencing of the extracted DNA was performed using the PCR primer set and a 3730 Genetic Analyzer (Applied Biosystems, Foster City, CA) by the Molecular Biology Research Facility at the University of Tennessee. Nucleotide sequence of the 228-bp segment revealed a 100% identity to P. tenuis and a 99% identity to Parelaphostrongylus andersoni (GenBank accession Nos. GU122925 and EF173713).
The natural host of P. tenuis is the white-tailed deer. Adult parasites live in the subdural space. A survey of white-tailed deer in northern New York between 1986 and 1989 showed that 46% were infected with meningeal worms. 6 Larvae penetrate the venous sinuses and are transported to the lungs, where they can be coughed up and passed in mucus covering the fecal pellets. The mucus is ingested by snails and slugs, which serve as the intermediate host and may be consumed by a variety of herbivores. Once ingested, the larvae penetrate the gastrointestinal tract wall and migrate along spinal nerves to the dorsal horn of the spinal cord and then to the subdural space. Infection in white-tailed deer is usually asymptomatic, except with heavy parasite burdens or in debilitated animals. 4 However, in other species, aberrant parasite migration causes central nervous system damage, which is associated with neurologic signs and may be fatal. Clinical signs include ataxia, stiffness, muscle weakness, hypermetria, posterior paresis, paralysis, head tilt, arching neck, circling, blindness, gradual weight loss, depression, seizures, and death. Often neurologic deficits begin in the hind limbs. P. tenuis–associated neurologic disease has been reported in moose, elk, caribou, wolves, antelope, reindeer, mule deer, black-tailed deer, sika deer, sheep, goats, calves, llamas, bison, alpacas, and horses. 2,3,7 –12,14,15 Guinea pigs experimentally infected with P. tenuis develop neurologic signs and CNS lesions similar to those seen in this case. 1,13 In the sections examined, there was no evidence of penetration of the neuroparenchyma by the nematodes, suggesting that either the clinical signs were secondary to the inflammatory reaction to the migrating larvae or that there was direct damage to the brain and/or spinal cord that was not detected in our samples. In experimental infections, neurologic signs were seen in guinea pigs without evidence of CNS penetration by larvae. 1
To the authors’ knowledge, this is the first report of naturally occurring neurologic disease associated with P. tenuis infection in guinea pigs. Clinical signs and histologic findings were similar to those reported in other aberrant hosts. A younger guinea pig from the same household that has also been fed deer-grazed grass is, to date, clinically normal, possibly indicating a difference in susceptibility to infection or clinical manifestations of infection. This case suggests that guinea pigs should not be fed grass from lawns frequented by white-tailed deer in P. tenuis–endemic areas.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.