
Abstract
For decades, the advancement of cancer research has relied on in vivo models for examining key processes in cancer pathogenesis, including neoplastic transformation, progression, and response to therapy. These studies, which have traditionally relied on rodent models, have engendered a vast body of scientific literature. Recently, experimental cancer researchers have embraced many new and alternative model systems, including the zebrafish (Danio rerio). The general benefits of the zebrafish model for laboratory investigation, such as cost, size, fecundity, and generation time, were quickly superseded by the discovery that zebrafish are amenable to a wide range of investigative techniques, many of which are difficult or impossible to perform in mammalian models. These advantages, coupled with the finding that many aspects of carcinogenesis are conserved in zebrafish as compared with humans, have firmly established a unique niche for the zebrafish model in comparative cancer research. This article introduces methods for generating cancer models in zebrafish and reviews a range of models that have been developed for specific cancer types.
The zebrafish (Danio rerio) emerged as a model for scientific research in the 1930s, with many early studies devoted to the investigation of developmental processes in embryos. 53 The use of zebrafish in research has subsequently expanded into a broad range of disciplines. As a research species, zebrafish offer certain advantages over traditional vertebrate models, including rapid generation time, high fecundity, external embryonic development, a capacity for high stocking density in relatively small areas, and lower maintenance costs. 64 In recent years, the zebrafish has come into its own as a relevant comparative model for a diverse spectrum of human diseases.
Although zebrafish are relative newcomers to the field of comparative cancer research, these small animals have emerged as a powerful tool for investigators. As vertebrate organisms, they share many features of embryonic development and adult anatomy that are critical for comparative animal models. 64,71 In the world of cancer research, this has important implications ranging from conserved signaling pathways and transcriptional regulatory mechanisms, to organogenesis and tissue differentiation, to cellular transformation and tumor initiation. Zebrafish cancer models have been produced by a variety of methods, leading to a broad spectrum of cancer susceptibilities in animals of multiple age groups. Flexibility in genetic manipulation, adaptability to current tools for cancer analysis, and capacity to define cellular and molecular interactions in vivo provide the zebrafish with its own very unique set of advantages for the investigation of cancer biology.
Mechanisms for Developing Cancer Models in Zebrafish
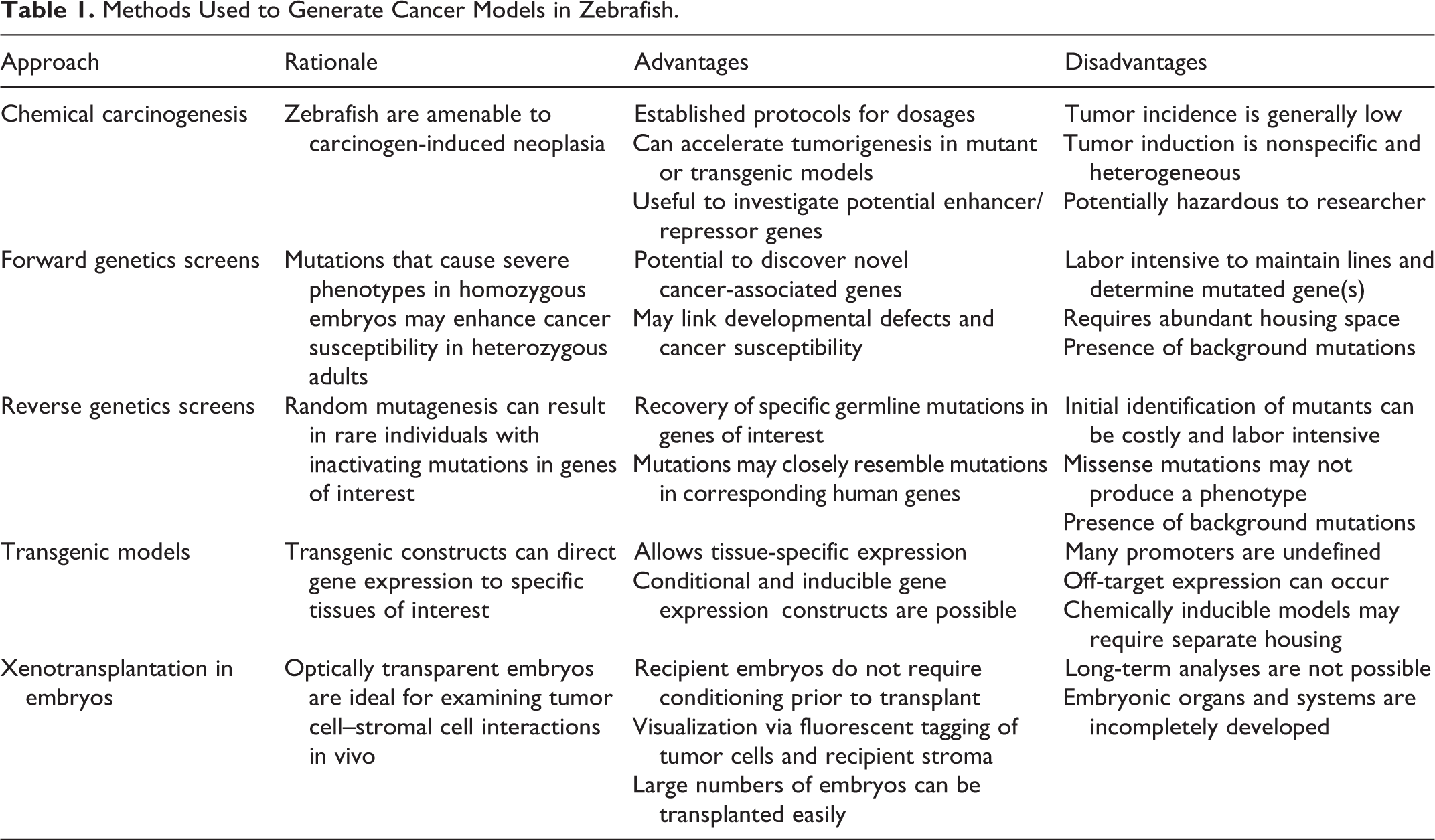
Zebrafish cancer models have been developed by a number of mechanisms (Table 1). Although initial studies relied on chemical carcinogens for tumor induction, 105 cancer modeling in zebrafish has expanded to include numerous mutant and transgenic lines. In this setting, chemical carcinogens are often useful for identifying tumor-prone zebrafish lines from forward genetic screens 97,98 or in combination with genetic manipulation in defining a tumor susceptibility phenotype. 39 Although the use of forward genetic screens to develop cancer models is not typical in traditional animal models, multiple zebrafish cancer models have been successfully derived by this method. 4,5,32,73,97,98 In some cases, this approach has identified novel genes with a previously unknown or poorly defined link to cancer susceptibility. 4,97,98
Methods Used to Generate Cancer Models in Zebrafish.
Multiple chemical carcinogens have been studied in fish species to determine their efficacy in fish, to model human carcinogen exposure, and to assess environmental toxins. 7,11,56,104,105 The most common routes of carcinogen exposure in zebrafish are aqueous and dietary. 11,39,51,56,73,104,105 Additional methods used in fish species include embryo injection and topical application. 11,104 In comparison, exposure methods used in rodent models include introduction in feed or water, gastric lavage, inhalation, injection, or topical application. 43,44 Although the routes of exposure may differ in carcinogenicity studies with fish versus mammalian species, the liver is a primary target tissue for many carcinogens in both fish and rodents. 11,43,44,104
Another approach for generating cancer models in animals is through genetic manipulation. As embryonic stem cell technology is yet to be achieved in zebrafish, the generation of zebrafish lines with mutations in genes of interest has most often relied on reverse genetic screens of mutagenized zebrafish. This approach typically involves chemical or retroviral mutagenesis strategies to generate a large population of zebrafish, with each individual carrying a unique collection of randomly distributed mutations. 3,74 The offspring of each uniquely mutagenized individual can be screened for mutations in genes of interest and are used to establish a mutant line. 3,103 Mutant zebrafish lines that have been established by this method harbor mutations in important cancer susceptibility genes, including zebrafish orthologues of the human tumor suppressor genes TP53, APC, PTEN, and BRCA2, among others. 9,24,39,85,93,99
Creation of transgenic zebrafish lines is also possible, since 1-cell stage zebrafish embryos can be injected with plasmid or bacterial artificial chromosome (BAC) gene constructs with relative ease. 18,115 A number of technologies have been developed to increase the rate of early and efficient transgene integration. 107,108 Transgenic lines have typically relied on zebrafish promoters driving expression of human or mouse genes, 59,60,87,88 although in some cases, the zebrafish orthologue has been used succesfully. 78 The adaptation of conditional or inducible strategies for gene expression in zebrafish has greatly expanded the possibilities for genetic manipulation in this species. 37,57,63
Xenotransplantation of human or mouse cancer cells into zebrafish embryos has also proven to be a useful tool in cancer research (reviewed by Amatruda and Patton 2 ). As zebrafish embryos do not have a fully developed immune system, conditioning of recipients is not required in these studies. 80 Because of the optical transparency of zebrafish embryos and the availability of multiple zebrafish lines that express fluorescent proteins in normal tissues, xenotransplantation experiments can allow detailed in vivo examination of cell-cell and cell-stromal interactions within the context of neoplastic cell survival, migration, and invasion. 80 Xenotransplanted zebrafish embryos also offer a high-throughput system for evaluating novel drug therapies in vivo. 54 Although further discussion of this methodology is beyond the scope of this article, the zebrafish model clearly offers a unique and diverse spectrum of approaches for the study of cancer.
Before reviewing zebrafish models for human cancer, one should be aware of nomenclature conventions for zebrafish gene names and symbols, proteins, and genotypes. Guidelines for zebrafish nomenclature are available from the Zebrafish Information Network (ZFIN), an online database for zebrafish researchers. 106 In brief, zebrafish gene names and gene symbols are lowercase and italicized and are named after the mammalian orthologue whenever possible. The protein symbol matches the gene symbol, but is not italicized, and begins with an uppercase letter. For mutant zebrafish lines, the mutant allele is identified by a unique line designation code that indicates the institution of origin. Transgenic lines are generally identified by “Tg,” followed by the promoter and gene construct, and a unique line designation code that indicates the institution of origin. More detailed information on zebrafish nomenclature can be found on the ZFIN website (http://zfin.org/).
Models of Cutaneous Neoplasia
A basic understanding of zebrafish skin anatomy facilitates the discussion of zebrafish models of cutaneous neoplasia. The integument of fish includes a nonkeratinized stratified squamous epidermis and underlying dermis. 71,72 Teleost species (including zebrafish) have overlapping elasmoid scales that are covered by the epidermis in the trunk region. 72 Within the epidermis are keratinocytes, goblet cells, and alarm cells. 71 Pigmented cell populations, including melanocytes, iridiphores, and xanthophores, are predominantly located in the hypodermis between the dermal stratum compactum and underlying muscle layer. 40 As in mammals, pigmented cells in fish are of neural crest origin and are designated during embryogenesis. 13
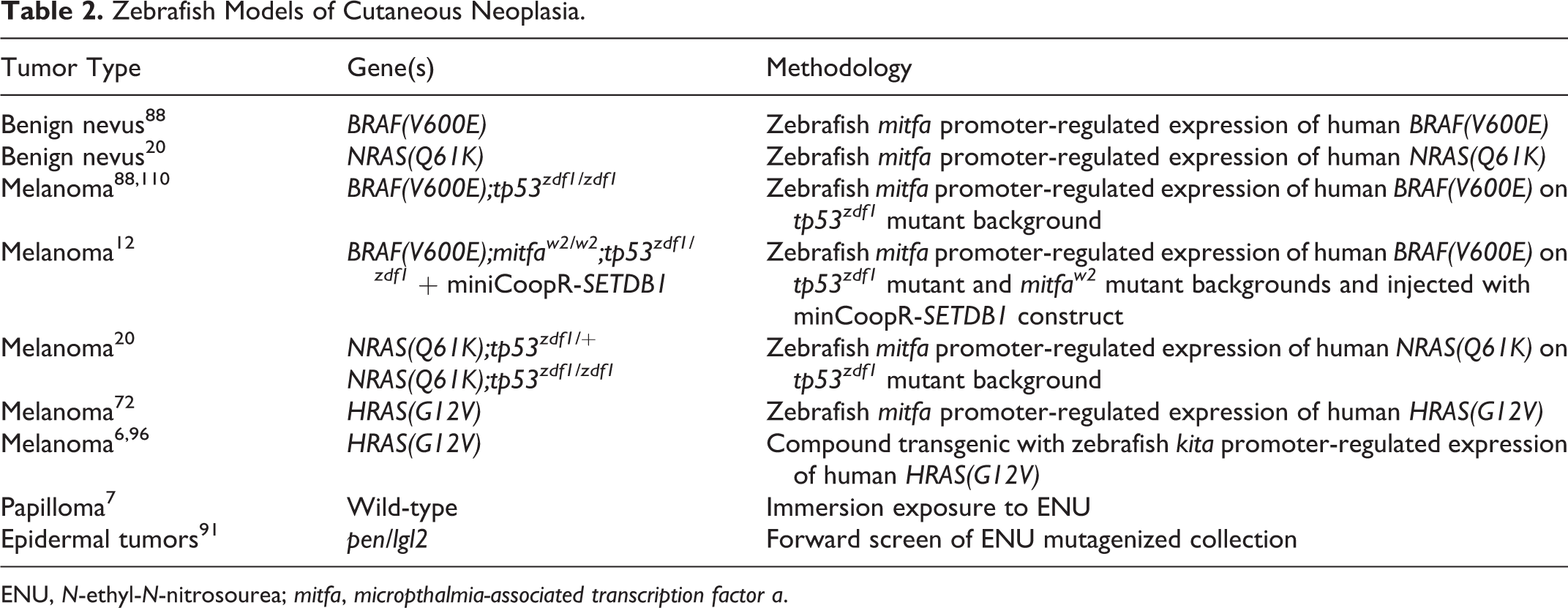
The majority of zebrafish models for cutaneous neoplasia have focused on melanocyte-derived tumors. Multiple models for benign nevus and cutaneous melanoma exist and have been derived by targeted expression of human oncogenes in melanocytes (Table 2 and Fig. 1). Most of these models use the melanocyte-specific promoter mitfa 66 to drive gene expression. MITF, or microphthalmia-associated transcription factor, is a critical mediator of melanocyte differentiation and survival in vertebrates and may also support survival of melanoma cells. 111 Similarly, the zebrafish orthologue of mammalian KIT is required for melanocyte migration and survival, 86 and the kita promoter also drives oncogene expression in zebrafish melanocytes. 96
Zebrafish Models of Cutaneous Neoplasia.
ENU, N-ethyl-N-nitrosourea; mitfa, micropthalmia-associated transcription factor a.
Zebrafish models for melanoma demonstrated that expression of a mutant allele of human BRAF (BRAFV600E) or NRAS (NRASQ61K) in zebrafish melanocytes could drive formation of benign nevi in adults, but progression to melanoma required concomitant p53 mutation. 20,88 In comparison, melanoma could be induced in zebrafish without p53 mutation through targeted expression of a mutant allele of human HRAS (HRASG12V). 6,72,96 These studies converge on activation of the Ras-Raf-MEK-ERK signaling pathway, the most significant initiating event in human melanoma (reviewed by Gray-Schopfer et al 36 ). As would be expected, there is robust activation of Ras-Raf-MEK-ERK signaling in these transgenic zebrafish models. 72,88 Phosphoinositide-3-OH kinase (PI(3)K) pathway activation, also shown to be an important factor in human melanoma, 36 is variable in zebrafish models. 72,96 Interestingly, the role for p53 in melanoma progression is not consistent in zebrafish models, as robust p53 pathway activation was observed in melanomas from kita-GFP-HRASG12V transgenic zebrafish. 96 In human melanoma, p53 mutations are relatively rare, 19 but p53 pathway inactivation is both common and critical. 50
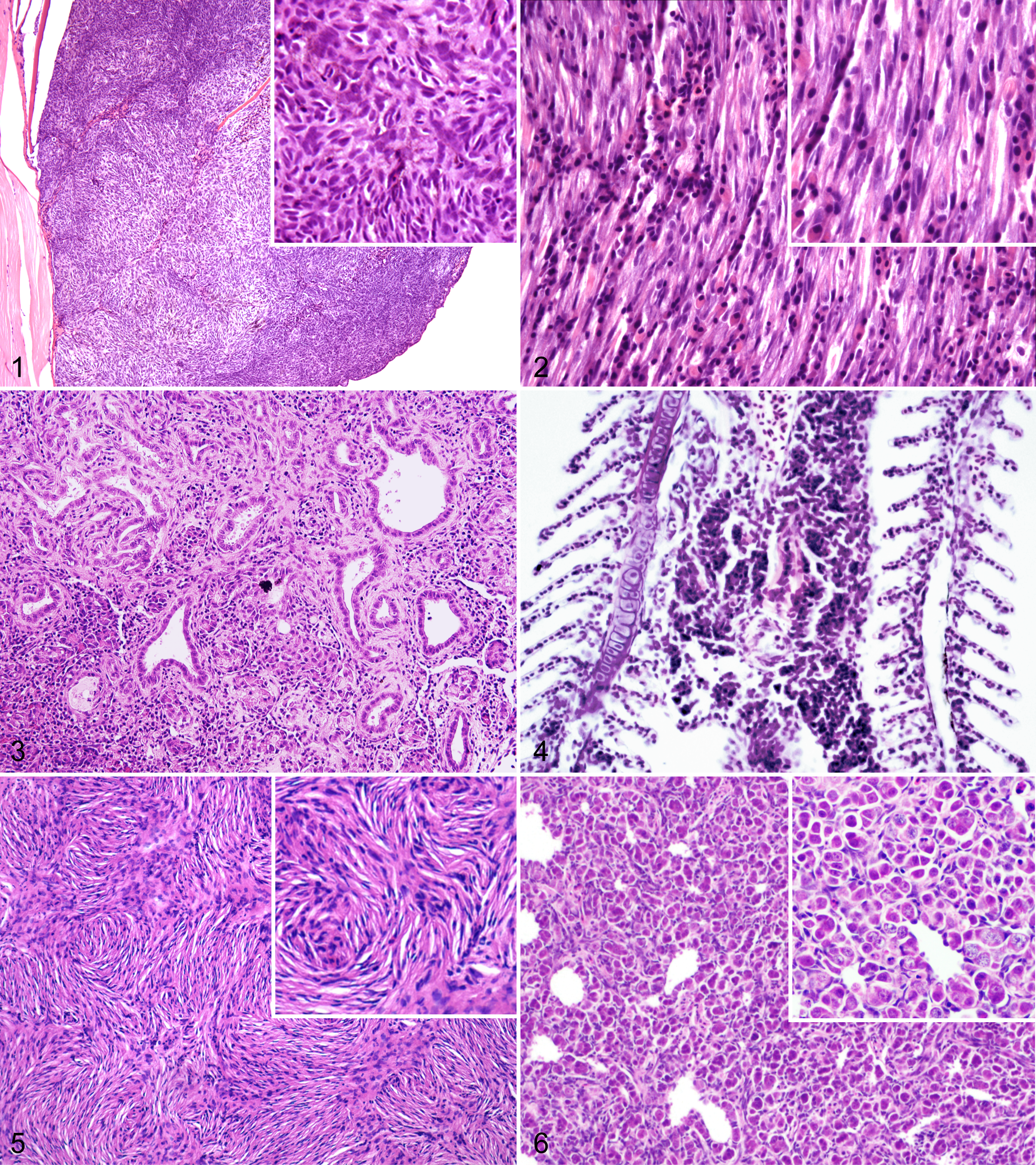
Abnormal stripe patterning and irregular areas of cutaneous pigmentation (hyperpigmentation) are commonly observed in zebrafish models of melanoma. 20,72,88,96 Areas of aberrant pigmentation were observed in zebrafish embryos as early as 3 days postfertilization (dpf) in some models. 96 Benign nevi arise in adult zebrafish as flat, discrete to coalescing patches that are heavily pigmented, randomly distributed over the head and body, and confined to the epidermis. 20,88 In comparison, zebrafish harboring melanomas present with variably nodular masses that exhibit invasive behavior. 20,72,88,96 Although the majority of zebrafish melanomas appear heavily pigmented grossly, neoplastic cells show variable degrees of pigmentation by histology. 20,72,88,96 Zebrafish melanomas are transplantable into sublethally irradiated recipients, 20,88 although benign nevi are not. 88 Additional characterization demonstrated that zebrafish melanoma cells are highly aneuploid, 88 are highly proliferative, 72,96 and exhibit immunoreactivity for tyrosinase, Melan-A, S100, and HMB-45, 96 markers that are useful in the diagnosis of human melanoma. 16
Zebrafish models of melanoma represent a powerful tool for multiple downstream experiments that delve more deeply into melanoma pathogenesis. Comparisons of microarray data from zebrafish and human melanoma samples have demonstrated similar patterns of gene expression and enrichment profiles, suggesting a similar biology among these tumors. 6,20 In a recent report, multiple candidate genes that are located in a recurrently amplified region of chromosome 1 in human melanoma were tested in a zebrafish melanoma model. 12 This study showed that overexpression of the histone methyltransferase SETDB1 significantly accelerated zebrafish melanoma formation and induced transcriptional dysregulation of multiple genes. 12 A zebrafish melanoma model has also revealed the importance of neural crest developmental pathways in melanoma formation. 110 Through a chemical screen for small-molecule inhibitors of neural crest development in zebrafish embryos, White et al 110 found that an inhibitor of neural crest progenitor cells could abrogate melanoma cell proliferation in vitro and in vivo.
Nonmelanoma models for cutaneous tumors in zebrafish are few (Table 2), although the convenience of this animal model for studying tumors of the skin is evident. In one study, adult zebrafish that were exposed to the mutagen N-ethyl-N-nitrosourea (ENU) developed sessile or pedunculated epidermal papillomas at 100% incidence. 7 However, no epidermal tumors developed during a subsequent ENU carcinogenicity study in juvenile zebrafish, although tumors of other tissues were observed. 104 Studies with other carcinogens in zebrafish have not described the induction of cutaneous neoplasia at any significant level (summarized by Spitsbergen and Kent 104 ). The pen/lgl2 mutant zebrafish line develops aberrantly proliferative epidermal cells during embryogenesis, which form small epidermal masses when transplanted into wild-type embryos (Table 2). 91 As pen/lgl2 mutants exhibit early embryonic lethality, 91,102 this line has limited use as a cancer model. Successful development of nonmelanoma models for cutaneous neoplasia in zebrafish may require a combination of techniques, such as genetic manipulation coupled with carcinogen exposure.
Models of Muscular, Adipocytic, and Vascular Neoplasia
Muscular and vascular tumors are described at low frequency in several zebrafish models with a diverse spectrum of cancer susceptibility. 48,98,105 However, there are only a small number of zebrafish models with a specific propensity for muscular, adipocytic, or vascular neoplasia (Table 3). Rhabdomyosarcoma and liposarcoma can be induced in transgenic zebrafish by targeting the Ras-Raf-MEK-ERK or PI(3)K-Akt pathways, respectively, whereas hemangiosarcoma in zebrafish is associated with haploinsufficiency for pten. 15,38,59,61
Zebrafish Models of Muscular, Adipocytic, and Vascular Neoplasia.

ENU, N-ethyl-N-nitrosourea.
The recombination activating gene-2 (RAG2) shows specific expression in mammalian lymphoid cells, although promoter activity has been observed in nonlymphoid cell populations. 81,117 In zebrafish, the rag2 promoter is expressed in both lymphoid and nonlymphoid cell populations, including mesenchymal progenitor cells. 47,59,60 Ectopic expression of the zebrafish rag2 promoter in these mesenchymal progenitor cell compartments led to development of models for rhabdomyosarcoma and liposarcoma. 38,59
Embryonal rhabdomyosarcoma developed in zebrafish that were injected at the 1-cell stage with a human KRASG12D transgene driven by the rag2 promoter (Fig. 2). 59 About 50% of mosaic transgenic zebrafish developed tumors by 80 dpf, with some animals developing masses as early as 10 dpf. 59 Histologic examination revealed undifferentiated myocytes and multinucleated cells with striations that expressed markers of myocyte differentiation, including desmin, myoD, and myogenin. 59 Serial transplantation of fluorescently labeled zebrafish rhabdomyosarcoma cells allowed isolation of a cancer stem cell population with an expression signature similar to activated satellite cells. 59 In another model, rhabdomyosarcomas were observed in 76% of compound transgenic zebrafish that expressed heat shock–inducible human KRASG12D under the zebrafish β-actin promoter. 61 Similar to the prior model, these tumors were composed of small undifferentiated myocytes and striated myocytes, and they expressed desmin, myoD, and myogenin. 61
In a similar approach, the zebrafish rag2 promoter driving myristoylated, constitutively active mouse Akt2 was used to develop a transgenic model for well-differentiated liposarcoma. 38 Although tumor incidence was low in a wild-type background, 29% of transgenic zebrafish with concomitant homozygous tp53 mutation developed liposarcomas. 38 Liposarcomas were composed of well-differentiated lipocytes intermixed with atypical stromal cells and lipoblasts with pleomorphic nuclei and were locally invasive. 38
Although rhabdomyosarcoma and liposarcoma models were developed by transgenic approaches, a model for hemangiosarcoma in zebrafish was recently described in zebrafish with germline mutations in the zebrafish orthologue of the tumor suppressor gene PTEN. 15 Zebrafish have 2 copies of pten (ptena and ptenb), and 2 mutant lines carrying nonsense mutations in each gene were identified from an ENU-mutagenized library. 24 Interestingly, embryonic development is normal with homozygous mutation in either gene alone; however, combined homozygous mutations in ptena and ptenb result in embryonic lethality. 24 Adult zebrafish with haploinsufficiency for the pten genes are predisposed to hemangiosarcoma, particularly in the retrobulbar region. 15 These locally invasive tumors exhibited variably sized, blood-filled spaces lined by CD31-positive neoplastic cells and were presumed to arise from the retrobulbar vascular network. 15
Zebrafish models for rhabdomyosarcoma and liposarcoma have provided interesting insights into the biology of these diseases in humans. On the basis of the finding that Ras activation resulted in a high incidence of rhabdomyosarcoma in zebrafish, Langenau et al 59 compared human and zebrafish rhabdomyosarcoma by microarray analysis. These studies revealed conserved gene signatures for rhabdomyosarcoma in both species and showed that Ras pathway activation is a common feature of human rhabdomyosarcoma. 59 Similarly, the discovery that Akt activation drives liposarcoma development in zebrafish led to the finding that the Akt pathway is activated in about a third of clinical liposarcoma specimens and that inhibition of the PI(3)K-Akt-mTOR pathway in human liposarcoma cell lines impairs viability. 38 These findings demonstrate that zebrafish can be a useful and relevant comparative animal model for human mesenchymal tumors.
Models of Intestinal, Hepatic, and Exocrine Pancreatic Neoplasia
Although the basic structures and functions of the intestine, liver, and exocrine pancreas are similar between zebrafish and mammals, some anatomical features in zebrafish are of note. As zebrafish are agastric, the esophagus leads directly into the intestine. 71 The intestine is a simple tubular structure that lacks distinguishable small and large intestinal segments, although differences can be observed in the density of goblet cells and morphology of the mucosal epithelium along its length. 71 The liver is composed of 3 lobes and a gallbladder. 71 Unlike mammals, hepatic cords, lobules, and portal triads are not clearly defined in the zebrafish liver, and Kupffer cells are absent. 71 The pancreas in zebrafish is anatomically distinct from the liver but does not form a discrete organ. 71 Clusters of exocrine pancreatic tissue are present along the intestinal tract; similar to mammals, pancreatic acini are composed of secretory cells containing bright eosinophilic granules. 71
Zebrafish that are heterozygous for a germline mutation in the tumor suppressor gene adenomatous polyposis coli (apc) 45 develop intestinal, hepatic, and pancreatic tumors (Table 4). 39 This tumor susceptibility phenotype is similar to the familial adenomatous polyposis syndrome in humans with hereditary APC mutations (reviewed by Sieber et al 100 ), indicating a conserved role for this gene in tumors of these tissues. Although apc heterozygotes developed adenomatous intestinal polyps and hepatomas without sensitization, exposure of juvenile apc-mutant zebrafish to the carcinogen 7,12-dimethylbenz[a]anthracene (DMBA) increased the incidence of intestinal and hepatic tumors and also induced pancreatic acinar cell adenomas and biliary duct neoplasia. 39 Tumors from apc-mutant zebrafish showed activation of Wnt signaling, like their human counterparts, but did not exhibit loss of heterozygosity for the wild-type apc allele; 39 however, tumors from human patients with APC mutations typically develop biallelic APC inactivation through the acquisition of somatic mutations in the wild-type allele. 100 Additional studies in apc-mutant zebrafish have defined a role for retinoic acid signaling and cyclooxygenase-2 (COX-2) expression in the pathogenesis of APC-associated neoplasia 22 and have been used to model a multistep progression from adenoma to carcinoma in tumors that arise in association with APC mutation. 90
Zebrafish Models of Intestinal, Pancreatic, and Hepatic Neoplasia.

DBP, dibenzo[a, l]pyrene; DMBA, 7,12-dimethylbenz[a]anthracene; ENU, N-ethyl-N-nitrosourea; fabp10a, fatty acid binding protein 10a, liver basic; xmrk, mutated Xiphophorus EGFRb.
Intestinal hyperplasia, dysplasia, and neoplasia have been described at variable frequency in several other transgenic and mutant zebrafish lines, 61,73,98 as well as in zebrafish exposed to carcinogens. 105 Importantly, malignant intestinal neoplasia can develop in zebrafish infected with the nematode parasite Pseudocapillaria tomentosa (Table 4). 51 Nematode-associated intestinal carcinomas typically arise at the esophageal-intestinal junction, often contain infiltrating inflammatory cells, and may be directly associated with adult nematodes. 51 In addition to providing an interesting model for parasite- and inflammation-associated intestinal neoplasia, it is important to consider P. tomentosa infestation as a potential confounding factor in zebrafish cancer models. This parasite has been shown to promote intestinal tumorigenesis in a zebrafish carcinogenicity study. 51 Although commonly found in some Florida wild-type zebrafish lines, 104 P. tomentosa appears to occur infrequently in today’s research colonies.
Zebrafish models for hepatic neoplasia have employed both genetic manipulation and carcinogen exposure. Carcinogens are relatively reliable for the induction of hepatic neoplasia and have been useful in defining conserved gene expression signatures in human and zebrafish hepatic tumors. 56,105 However, the lack of specificity for hepatic tumorigenesis in carcinogenesis studies and the desire for a stable and heritable cancer susceptibility phenotype have spurred the development of transgenic zebrafish models for hepatic cancer.
Several transgenic models for hepatocellular carcinoma (HCC) have been described, each using the liver-specific promoter fabp10 to drive oncogene expression (Table 4). 63,78,79 Although targeted expression of the human oncogene KRAS(G12V) resulted in variable incidence of HCC in juvenile and adult zebrafish, 78 development of a mifepristone-inducible KRAS(G12V) transgene targeted to the liver resulted in reliable induction of HCC with 100% penetrance. 79 Similarly, expression of a mutated form of EGFRb 35 in the liver controlled by a tetracycline-inducible element led to a 100% incidence of HCC in juvenile and adult zebrafish. 63 The histologic features of HCC in these models included expansion of the liver by sheets of pleomorphic hepatocytes with prominent nucleoli, with evidence of intravascular and regional tissue invasion. 63,79 These inducible models, which show complete tumor regression upon mifepristone or tetracycline withdrawal, represent useful tools for small-molecule screens, as well as for evaluating potential enhancer/repressor genes. 63,79
A transgenic approach has also been applied for developing 2 zebrafish models for exocrine pancreatic carcinogenesis (Table 4). In both models, an eGFP-KRAS(G12V) fusion gene controlled by regulatory elements for the zebrafish pancreatic progenitor cell marker ptfa1 has been used successfully to induce pancreatic carcinoma in zebrafish. 68,87 In ptfa1a: eGFP-KRASG12V zebrafish, progenitor cells expressing the transgene undergo minimal differentiation during embryogenesis and subsequently instigate several types of malignant pancreatic neoplasia in adult zebrafish. 87 These locally invasive tumors are of both acinar and ductular origin, and they exhibit varying degrees of acinar cell differentiation, stromal expansion, and mucin production. 87 Similar to human and murine pancreatic adenocarcinomas, Hedgehog pathway activation occurred in neoplastic cells. 87 The second model is a compound transgenic zebrafish line generated with the Gal4-UAS system that targets expression of the eGFP-KRAS(G12V) fusion gene to the exocrine pancreas, leading to development of acinar and ductular pancreatic carcinoma (Figure 3). 68 This model will allow the simultaneous expression of additional candidate oncogenes in the zebrafish pancreas, facilitating the study of complex genetic interactions in pancreatic carcinogenesis. 68 The histologic and genetic similarities between human and zebrafish exocrine pancreatic tumors make this an appealing model for comparative studies.
Models of Hematopoietic and Lymphoid Neoplasia
A relatively large body of work is devoted to zebrafish models for hematopoietic and lymphoid neoplasia (Table 5), including the first report of a line of transgenic tumor-bearing zebrafish. 60 Several key anatomic and histologic differences between zebrafish and mammals are relevant for the study of these diseases. Unlike mammals, zebrafish bones do not have a medullary cavity containing hematopoietic tissue; instead, hematopoiesis occurs in the splenic stroma and renal interstitium. 71 Zebrafish lack lymph nodes but do possess a thymus with cortical and medullary organization that supports lymphocytic maturation. 71 Peripheral blood components include erythrocytes and thrombocytes (both nucleated in teleost species), 46,71 neutrophilic and eosinophilic granulocytes, 8,17,65 monocytes, 75 and B and T lymphocytes. 71,75 Lymphocytes are the predominant circulating leukocyte in zebrafish, representing 71% to 92% of the population. 75
Zebrafish Models of Hematopoietic, Lymphoid, and Small Round Blue Cell Neoplasia.

AML, acute myeloid leukemia; B-ALL, B-cell acute lymphocytic leukemia; ENU, N-ethyl-N-nitrosourea; spi1, spleen focus forming virus (SFFV) proviral integration oncogene spi1; T-ALL, T-cell acute lymphocytic leukemia; T-LBL, T-cell lymphoblastic lymphoma.
The majority of transgenic models for T-cell lymphoma/leukemia in zebrafish make use of the rag2 promoter, although as stated previously, this promoter can be expressed in nonlymphoid populations. Most of these models have induced neoplastic transformation in lymphoid cells via expression of the mouse or human MYC oncogene (Table 5 and Fig. 4), as overexpression of MYC or MYCN is a common feature of most human T-cell acute lymphocytic leukemia (T-ALL) subgroups. 29 Targeted expression of the MYC oncogene in zebrafish lymphocytes resulted in expansion of a malignant population of T lymphocytes that heavily infiltrated the thymus, kidney, and perithymic soft tissues. 60 Malignant T cells were oligoclonal, aneuploid, or hyperdiploid and were transplantable to sublethally irradiated recipients. 58,60 Additional approaches for modeling T-cell lymphoblastic lymphoma (T-LBL) and T-ALL, including the identification of several mutant lines through a forward genetic screen, have been successful (Table 5). 14,32
More complex transgenic approaches have modeled the progression from T-LBL to T-ALL in zebrafish. Use of conditional promoters in compound transgenic zebrafish have shown that zebrafish develop T-LBL, characterized by thymic enlargement and local infiltration, which progresses to T-ALL, characterized by widely disseminated disease and an abundance of malignant cells in the kidney marrow. 27,28 These types of complex genetic manipulations have permitted investigation into the mechanisms of malignant progression in zebrafish and human lymphoid neoplasia. Overexpression of the antiapoptotic gene bcl2 blocked progression from T-LBL to T-ALL in zebrafish, and malignant cells were defective in intravasation and underwent autophagy. 28 Further analyses of both zebrafish and human samples in this study defined molecular differences between T-LBL and T-ALL, with important implications for cell transformation, survival, and intravasation in vivo. 28 In another study, creation of an inducible model for T-ALL in zebrafish demonstrated the importance of the Akt pathway in accelerating tumorigenesis in combination with MYC. 37 A line of syngeneic zebrafish has been described that can be used for high-throughput, rapid screening of leukemic adult zebrafish, permitting the analysis of tumor-initiating cell populations in transplanted immunocompetent recipients. 101 Zebrafish models for lymphoid malignancies thus represent a sophisticated in vivo tool for examining the mechanisms underlying transformation and disease progression.
Zebrafish are also useful in studying acute myeloid leukemia (AML), although there are fewer models for this disease (Table 5). Unlike models for T-cell malignancies, zebrafish models for AML have generally relied on the expression of human fusion genes in myeloid cells. Several of these models have demonstrated that expression of these fusion genes induces disturbances in the myeloid compartment, including redirection of progenitor cell fate from erythroid to myeloid lineage and expansion of progenitor and myeloid compartments in the marrow. 10,83,116 Transgenic zebrafish expressing the human fusion gene NUP98-HOXA9 under a heat shock–inducible promoter developed malignant myeloid cell infiltrates in multiple organs. 31 Since zebrafish embryos develop externally, the controlled expression of NUP98-HOXA9 at certain time points during embryogenesis could be used to define specific effects on myeloid differentiation in early hematopoiesis. 31 These and other models hold great promise for the study of myeloid malignancies in vivo.
Few other models for lymphoid or other small round cell tumors exist in zebrafish (Table 5). A single zebrafish model for B-cell acute lymphocytic leukemia (B-ALL) has been described, in which the human TEL-AML1 fusion was expressed under ubiquitous promoters. 94 Transgenic zebrafish developed lymphoblastic leukemia at low incidence, and neoplastic cells appeared to be arrested at a pre–B-cell differentiation stage. 94 In addition, a zebrafish model transgenic for the human EWS-FLI1 fusion was recently found to develop small round blue cell tumors at low incidence. 62
Models of Neural and Neuroendocrine Neoplasia
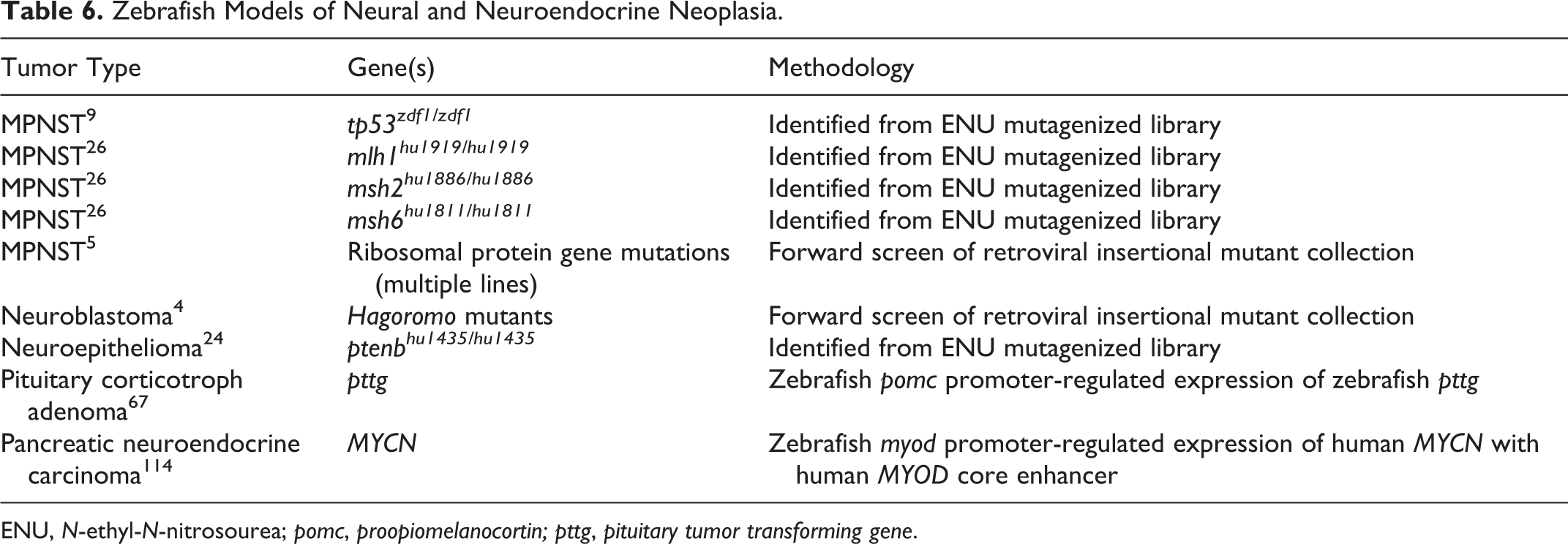
Tumors of the peripheral nervous system are not uncommon in zebrafish cancer models (Table 6). A number of mutant zebrafish lines develop malignant spindle cell tumors that are histologically reminiscent of malignant peripheral nerve sheath tumors (MPNSTs). 5,9,26,85,92 Curiously, these cancers arise with greatest frequency in zebrafish lines that harbor mutations in tumor suppressor genes (Table 7), suggesting that the cancer spectrum associated with these types of mutations may be more limited in zebrafish compared with humans. MPNSTs in these mutant lines typically arose in zebrafish older than 1 year. 5,9,85,92
Zebrafish Models of Neural and Neuroendocrine Neoplasia.
ENU, N-ethyl-N-nitrosourea; pomc, proopiomelanocortin; pttg, pituitary tumor transforming gene.
Tumor Spectrum in Humans and Zebrafish with Mutations in Tumor Suppressor Genes.

MPNST, malignant peripheral nerve sheath tumor.
aThe most commonly reported cancers are listed.
Histologic features of these tumors are similar between mutant models. Neoplastic cells are typically spindloid to variably epithelioid with oval to tapered nuclei, and they are arranged in fascicles, stacks, and whorls (Fig. 5). 5,9,26,85 These highly invasive tumors can arise in any of 3 distinct sites: the retrobulbar region, the coelom, and the flank or tail. 9,26,85,99 Interestingly, MPNSTs in zebrafish are highly aneuploid and show nonrandom regions of chromosomal amplifications and losses, even when tumors are derived from different mutant zebrafish lines. 118
Classification of these tumors as MPNSTs has been somewhat controversial, as these tumors reportedly do not express S100 or PGP9.5, 85 markers that have been used for identification of human MPNSTs. 30,41 However, the report that zebrafish MPNSTs do express glial fibrillary acidic protein (gfap) by RNA in situ hybridization 9 supports a neural origin for these tumors, as human MPNST can be immunoreactive for GFAP. 30 The difficulty in classifying these malignant spindle cell tumors, as well as other tumors that arise in zebrafish cancer models, highlights the need for zebrafish-specific antibody panels that will complement histologic analyses.
In a forward genetic study, 4 mutant zebrafish lines (designated as Hagoromo mutants) derived from a retroviral insertional mutant library were susceptible to neuroblastoma-like tumors of the head. 4 Heterozygous Hagoromo mutants developed expansile masses composed of small, ovoid cells in densely packed sheets with infrequent rosette formation; cells were positive for the pan-neural marker HuC. 4 Juvenile Hagoromo mutants exhibited aberrant persistence of putative neural progenitor cells in cranial ganglia that were proposed to be a preneoplastic population. 4 In all 4 lines, retroviral integration resulted in activation of an upstream gene, fibroblast growth factor 8 (fgf8). 4 The role for fgf8 in neoplastic transformation or progression is unclear but may promote progenitor cell proliferation or survival. 4
There are very few models for tumors of neuroendocrine origin in zebrafish (Table 6). A single zebrafish model for pituitary corticotroph adenoma has been described. 67 In this model, overexpression of the pituitary tumor transforming gene (pttg/securin) in cells of the adenohypophyseal proopiomelanocortin (POMC) lineage leads to formation of pituitary corticotroph adenomas in adult transgenic zebrafish. 67 Interestingly, adult transgenic zebrafish developed multiple signs of endocrine dysfunction, including hypercortisolemia, hyperplasia of renal steroidogenic cells, relative insulin resistance, hepatic lipidosis, and cardiomyopathy. 67 Tg(pomc-pttg) zebrafish embryos were successfully used to identify small-molecule inhibitors of pituitary POMC cell expansion, which were subsequently found to inhibit growth of neoplastic corticotrophs in vitro and in vivo. 67
A model for pancreatic neuroendocrine carcinoma in zebrafish has also been described, although a stable transgenic line has not yet been established (Table 6). Zebrafish that overexpressed MYCN in the pancreas developed invasive, insulin-expressing carcinomas, with neurosecretory granules identified in neoplastic cells by transmission electron microscopy, presumably reflecting malignant transformation of pancreatic β-cells. 114
Much remains to be explored in the development of transgenic or mutant zebrafish models for tumors of neural and neuroendocrine origin. The development of reporter lines defining functional promoter regions for the neural-specific genes nestin and gfap 55 will be useful tools for targeting gene expression to cells of the central nervous system. Although certain endocrine tissues are organized differently in the zebrafish, 71 the success of the above models in recapitulating major features associated with the equivalent diseases in humans suggests that development of novel models for neuroendocrine neoplasia in zebrafish will be fruitful.
Models of Gonadal Neoplasia
There are a small number of zebrafish models for reproductive tumorigenesis, all of which show gonadal tumor susceptibility in males (Table 8). Gonadal tumors in male zebrafish have also been observed at variable frequency in several studies reporting a spectrum of tumor susceptibility. 73,98,105 In contrast, ovarian tumors are rarely reported in transgenic, mutant, or carcinogen-exposed female zebrafish. 73,77
Zebrafish Models of Gonadal Neoplasia.
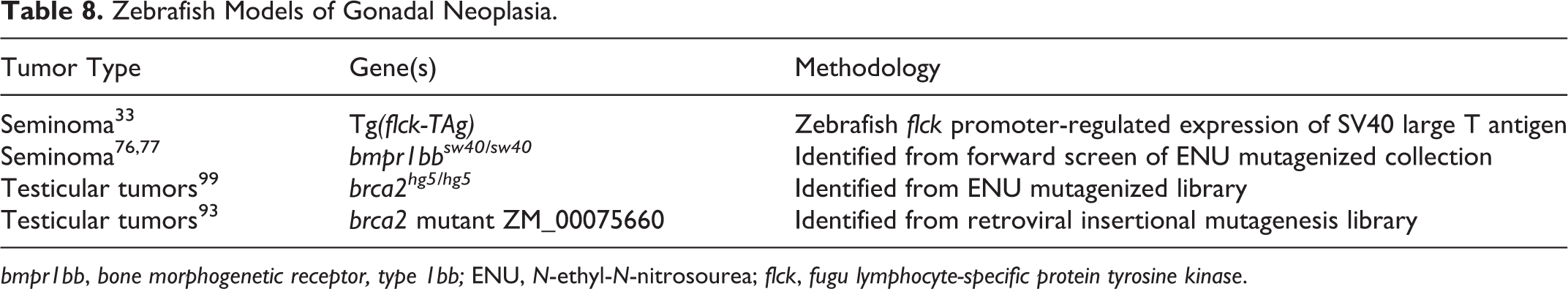
bmpr1bb, bone morphogenetic receptor, type 1bb; ENU, N-ethyl-N-nitrosourea; flck, fugu lymphocyte-specific protein tyrosine kinase.
Specific susceptibility to testicular germ cell tumors (seminomas) has been described in a transgenic zebrafish model 33 and in a mutant zebrafish line. 77 In tg(flck-TAg) zebrafish, off-target expression of the SV40 large T antigen by the fugu lymphocyte-specific protein tyrosine kinase (flck) promoter in the testis led to seminoma development in older adult males. 33 In comparison, zebrafish with a nonsense mutation in the type 1B bone morphogenetic protein receptor alk6b developed seminomas at a median age of 7 months. 76,77 Both tg(flck-TAg) and alk6b-mutant zebrafish showed variable degrees of differentiation in seminomas, ranging from tumors with clear spermatocytic differentiation to tumors composed of homogeneous sheets of undifferentiated neoplastic spermatogonia. 33,77 Interestingly, alk6b-mutant female zebrafish did not develop ovarian tumors spontaneously, although germ cell differentiation was markedly impaired. 76,77 Rare ovarian germ cell tumors occurred in alk6b-mutant female zebrafish after exposure to DMBA. 77 Undifferentiated human germ cell tumors showed loss of bone morphogenetic protein (BMP) signaling, suggesting that BMP signaling may influence germ cell differentiation and transformation in human and zebrafish gonads. 76
Testicular tumors have also been reported in 2 different zebrafish lines with mutations in brca2 (Table 8). In humans, carriers for pathologic BRCA2 mutations are at increased risk for tumors of reproductive organs (breast and ovary); 52,113 however, the specific association between BRCA2 mutation and reproductive tumorigenesis is not understood. In both brca2-mutant zebrafish lines, homozygous brca2 mutation led to an all-male phenotype characterized by arrested spermatogenesis. 93,99 Although the absence of brca2-homozygous female zebrafish impeded evaluation of ovarian cancer susceptibility, male brca2 homozygotes were susceptible to testicular neoplasia of both somatic and germ cell origin (Figure 6). 93,99 Zebrafish with combined mutations in brca2 and tp53 developed MPNSTs and other sarcomas in multiple tissues, and female zebrafish with these mutations showed an increased incidence of tumors arising in the ovary. 93,99 These studies provided new insight into the role for BRCA2 in reproductive development and demonstrate a conserved role for BRCA2 in reproductive tumorigenesis in humans and zebrafish.
Zebrafish Cancer Models: Future Directions
There are currently numerous undefined or underexplored areas for cancer research in zebrafish models, some of which have been noted in this review. Models for cancers of the brain, bone, and ovary remain to be developed; many other types of cancer models, including nonmelanoma cutaneous tumors, certain mesenchymal tumors, and endocrine tumors, are thus far quite limited. Although the absence of some tissue types in fish species, such as breast and prostate, limits the usefulness of zebrafish for investigating cancers of these organs, the zebrafish model has nonetheless become a powerful tool in comparative biomedical research. New methods for creating gene modifications or disruptions, such as zinc finger nucleases and transcription activator-like effector nucleases (TALENS), are being applied in zebrafish to generate targeted, heritable genetic mutations. 21,42,70,95 Recent technological advances for analyzing tumor development in zebrafish in vivo, including magnetic resonance imaging, 49 ultrasound biomicroscopy, 34 and high-throughput fluorescent macroscopic imaging, 101 reflect the development and adaptation of increasingly sophisticated systems tailored to zebrafish research.
The successful development of numerous zebrafish models for human cancer clearly demonstrates that mechanisms for neoplastic transformation and tumor progression are conserved across relatively wide evolutionary distances. Although it is significant that many features of cancer development in zebrafish faithfully recapitulate important aspects of their human counterparts, the usefulness of zebrafish as a cancer model extends far beyond mere phenotypic or genotypic replication of disease. Multiple studies have shown that judicious and creative use of zebrafish cancer models can lead to important insights into disease pathogenesis and therapy. Such approaches may range from small-molecule chemical screens to high-throughput investigations of potential enhancer/repressor genes to transplantation or xenotransplantation experiments. A key strength of the zebrafish model in cancer research is the ability to seamlessly transition between embryonic and adult animal studies, providing coordinated exploration of critical developmental, transformative, and pathological events in vivo. These attributes ensure that zebrafish models for human cancer are here to stay.
Footnotes
Acknowledgements
The author thanks Dr Dennis Hickstein, Dr Robert West, and Ms Lisa Embree for critical review of the manuscript and Dr Leonard Zon, Dr Charles Kaufman, Dr Colleen Albacker, Dr Narie Storer, Dr Steven Leach, Dr Shu Liu, Dr David Langenau, and Dr Jessica Blackburn for contributions of histologic images.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Intramural Research Program of the US National Institutes of Health, National Cancer Institute, Center for Cancer Research.