
Abstract
The pregnant guinea pig is an effective model for studying abortifacient Campylobacter spp, and previous experiments have demonstrated that C. jejuni IA3902 has a marked predilection for the subplacenta while sparing the placental disc in this species. In the study described here, the growth and chemotaxis of IA3902 and a reference strain (NCTC 11168) are compared in the presence of subplacental and placental factors, as well as bile and plasma, from pregnant and nonpregnant guinea pigs. Both strains grew better in subplacental versus placental disc tissue extracts at 24 hours; however, only IA3902 maintained this enhancement at 48 hours. Histochemistry and lectin histochemistry were used to localize mucin, iron, and
Campylobacteriosis is a significant cause of ovine abortion worldwide, with abortion rates ranging from 5% to 50% in affected flocks. 31 In a 2001 survey by the US Department of Agriculture, campylobacteriosis ranked first among infectious causes of abortion in domestic sheep. 34 Recent studies have revealed that Campylobacter jejuni has become the predominant species associated with ovine abortions in the United States, 8,30 and further analysis using pulsed-field gel electrophoresis has demonstrated that within these C. jejuni isolates, a single tetracycline-resistant clone (named clone SA for “sheep abortion”) has emerged as the predominant cause of ovine abortion in Iowa, South Dakota, Idaho, California, Oregon, and Nevada. 30 The identification of this single clone recovered from abortions in different regions and during different lambing seasons is significant in that Campylobacter spp traditionally exhibit marked genetic and antigenic heterogeneity. 8,9,22,35
The chemotactic behavior of C. jejuni has been described in response to specific chemical elements, with strong chemoattraction observed toward mucin,
The marked virulence of clone SA has been demonstrated using oral inoculation of C. jejuni IA3902, a field isolate of clone SA, in pregnant guinea pigs, which induced abortion at a rate of 60%, whereas oral inoculation with C. jejuni NCTC 11168 or a sheep fecal isolate did not induce abortion in any inoculated animals. 4 In this previous study, 4 C. jejuni IA3902 was recovered from the blood, bile, feces, uterus, and placenta of aborting animals, while NCTC 11168 and the sheep fecal isolate were recovered from only the feces. This finding strongly indicates that IA3902 is highly invasive and causes systemic infection in pregnant animals. Based on immunohistochemistry, localization of C. jejuni organisms has been described primarily within subplacental trophoblasts, the lateral aspects of the subplacental/decidual junctional zone, and surrounding maternal arteries in the decidua while the main placenta was spared, 4 suggesting that C. jejuni IA3902 is differentially localized in the placental unit. Despite these advances in understanding the pathogenesis of clone SA, the factors influencing its pathogenicity and differential localization in infected tissues are still unknown. The aims of this current study were fourfold: (1) to compare the growth and chemotaxis of IA3902 versus NCTC 11168 in tissue extracts from the guinea pig subplacenta and placenta, (2) to characterize the spatial distribution of putative growth-promoting factors in various regions of the guinea pig placental unit, (3) to evaluate the effects of these potential factors on the growth and chemotaxis of IA3902 and NCTC 11168 in vitro, and (4) to compare the effects of pregnant versus nonpregnant guinea pig plasma and bile on the growth and chemotaxis of both strains. These studies were undertaken in an effort to shed light on potential mechanisms that may underlay the fetoplacental tropism and spatial localization of C. jejuni IA3902 during septic abortion in guinea pigs.
Materials and Methods
Bacterial Strains and Growth Conditions
C. jejuni IA3902 is a clinical isolate belonging to clone SA, which is commonly isolated from sheep abortions in the United States. 30 C. jejuni NCTC 11168 is a human isolate associated with gastroenteritis and not known to be associated with abortion. 12 The isolates were grown routinely on Mueller-Hinton (MH) agar plates at 42°C under microaerobic conditions (5% O2, 10% CO2, 85% N2). Only second passages on MH agar (grown for about 20 hours; hereafter, fresh overnight culture) of frozen stock cultures were used to provide consistency for inoculation and avoid damage to the bacterial cells due to multiple laboratory passages. For inoculation, fresh cultures were adjusted to OD600 of 0.1, which gives approximately 1 × 10 7 colony-forming units per milliliter (CFU/ml) as determined by viable CFU plate counting. Minimal essential medium (MEM; MEM α, 41061-029, Invitrogen, US) without phenol red was used to determine the effect of various tissue extracts, biological fluids, or chemicals on the growth of C. jejuni strains as described below.
Humane Animal Care and Use
All procedures were approved by the Institutional Animal Care and Use Committee at Iowa State University, Ames, Iowa.
Animals
Twenty female Hartley guinea pigs were obtained from a commercial source (Elm Hill Labs, Chelmsford, Massachusetts). Upon arrival, 10 of the animals were pregnant at approximately 35 days of gestation (midterm), and 10 were nonpregnant. Pregnancy was confirmed by abdominal palpation. Guinea pigs were group housed, provided wood chip bedding, fed a commercial pelleted guinea pig diet ad libitum, and allowed to acclimate for 5 days.
Euthanasia and Necropsy Sampling
All animals were euthanized 5 days after arrival via intraperitoneal injection of sodium pentobarbital (approximately 150 mg/kg) and necropsied immediately. At necropsy, the entire uterus was removed intact using sterile surgical technique. In a laminar flow hood, the uterus was incised with a sterile surgical blade, and each placental disc (along with its associated subplacenta) was carefully removed as a unit. Each placental unit was then cut in half longitudinally and one part placed into 10% neutral buffered formalin while the other half was prepared for use in the in vitro growth and taxis assays. In preparation for these assays, careful aseptic technique was used to separate the subplacenta from the placental disc using sterile curved surgical scissors. The separated subplacenta and placental disc were placed into separate sterile specimen bags (Whirl-pak, Nasco, US) and then pooled for pairs of guinea pigs to yield a total of 5 samples for analysis. For both pregnant and nonpregnant animals, heart blood and bile were collected using sterile needles and syringes; blood samples were placed in anticoagulant tubes; and plasma was separated for analysis. Due to small sample volumes, plasma samples were also pooled for each pair of guinea pigs (5 samples total), and bile samples were pooled for groups of 2 or 3 guinea pigs (4 samples total). Placental samples taken for histopathology remained in neutral buffered formalin for 24 hours and were then embedded in paraffin and processed routinely for histochemical staining.
Histology and Histochemistry
Serial sections from each placenta were cut to 5 μm and stained with Perl’s iron stain, Alcian blue pH 2.5, and the periodic acid–Schiff (PAS) reaction, with and without diastase pretreatment. Sections were evaluated to determine the presence or absence of material with staining characteristics consistent with iron, acid mucin, and neutral mucin within the different aspects of the placental unit (placental disc, visceral yolk sac placenta, parietal yolk sac placenta, subplacenta, and decidual junctional zone). Each area of each placenta was then quantified subjectively per the amount of positive staining in a random 40× field: negative, if no staining was observed; low, if < 10% of the field stained positively; moderate, if 10% to 25% of staining was observed; and abundant, if > 25% of the field stained positive. Staining was also evaluated for distribution within each tissue type and classified as intracellular or extracellular and multifocal or diffuse.
Lectin Histochemistry
Serial sections from each placenta were cut to 3 μm, placed on aminoalkylsilane-coated glass slides, and placed in a 56°C oven for 2 hours. Sections were routinely deparaffinized in xylene and rehydrated in graded alcohol and water baths. To inhibit endogenous peroxidase, sections were immersed (2 immersions, 10 minutes each) in 3% hydrogen peroxide in water. For antigen unmasking, sections were treated with Tris-EDTA (pH 9.0) in a stream bath for 20 minutes, allowed to cool to room temperature, and then rinsed 3 times in phosphate buffered saline (PBS) prior to placement in an automated cell staining system (BioGenex, US). Lectins employed consisted of commercially available biotinylated Ulex europaeus agglutinin I (UEA-I; Vector, US) and biotinylated Lotus tetragonolobus lectin (LTA; Vector) applied to sections at 20 μg/ml and incubated at 22°C for 30 minutes, followed by rinsing in a bath of PBS solution for 5 minutes. Lectin binding was visualized using a commercial kit (Vectastain Elite ABC, Vector) and chromogen (NovaRED, Vector) per the manufacturer’s instructions; the sections were then counterstained with hematoxylin and mounted routinely. Specificity of lectin binding was confirmed by preabsorbing lectins in 250mM of
Growth Assays
Subplacental and placental tissues were placed in PBS (10% w/v), crushed aseptically using bare hands, and centrifuged at 1000 g for 5 minutes to remove large tissue fragments, and the resultant supernatants were collected for use in the growth assays. Plasma samples were heat treated at 56°C for 30 minutes before use to inactivate complement, and bile samples were used directly. For all biological samples, 100 μl from an overnight culture of C. jejuni (grown on MH agar under microaerophilic conditions at 42°C) that had been adjusted to an OD600 of 0.1 (approximately 1 × 10 7 viable CFU/ml) in PBS was inoculated into separate culture tubes containing 2 ml of the biological sample to be tested, yielding a theoretical initial CFU count of 5 × 10 5 CFU/ml. At 24 and 48 hours, 100 μl of each inoculated sample was then plated onto MH agar plates at different dilutions and grown under microaerophilic conditions at 42°C to determine the number of CFUs for each sample.
Commercially available mucin (porcine gastric),
Chemotaxis Assays
A fresh overnight culture of each C. jejuni strain (IA3902 and NCTC 11168) on MH agar plates was harvested in MH broth, pelleted by centrifugation at 5000 g for 5 minutes, and then resuspended in PBS, with the bacterial concentration adjusted spectrophotometrically to approximately 4 × 10
9
CFU/ml, with temperate PBS soft agar (0.4%) as described by others.
15,36
Twelve milliliters of bacterial suspension was poured into a 90-mm sterile Petri dish, and 6-mm sterile filter discs (Whatman) saturated with approximately 50 μl of test solution were placed on the solidified bacterial suspension. Solutions tested included subplacental and placental extracts (10% and 50% solutions as prepared in the growth assays), undiluted plasma, undiluted heat inactivated plasma, undiluted nonpregnant bile, pregnant bile (25%, 50%, and 100%), iron (0.01M in double-distilled water), mucin (0.1% and 1.0% solutions autoclaved at 70°C for 30 minutes),
Statistical Analysis
A commercial statistical software package (JMP 8.0.2, SAS Institute Inc., US) was used to perform all analyses. Independent samples t tests were utilized to detect differences in bacterial growth by strain in the tissue homogenate assays and by pregnancy status in the bile assays. One-way analysis of variance was used to detect differences in bacterial growth between multiple substrates, with P values reported after Tukey adjustment for multiple comparisons. Fisher exact test for binomial variables was employed when comparing the presence or absence of positive Campylobacter culture by pregnancy status in the plasma assays.
Results
Histology and Histochemistry
To characterize the spatial distribution of putative growth-promoting factors within the guinea pig placental unit, histochemical staining for mucin and iron was employed. In sections stained with the PAS reaction following diastase pretreatment, abundant PAS-positive material compatible with neutral mucin was present within subplacental lacunae (Fig. 1a), while the associated lamellae did not stain. Moderate to abundant PAS-positive, diastase-resistant extracellular material was also identified throughout the junctional zone with frequent marked perivascular accumulation. A low level of diffuse apical cytoplasmic staining was noted in the visceral yolk sac endoderm, while the parietal yolk sac and placental disc did not stain (Table 1).
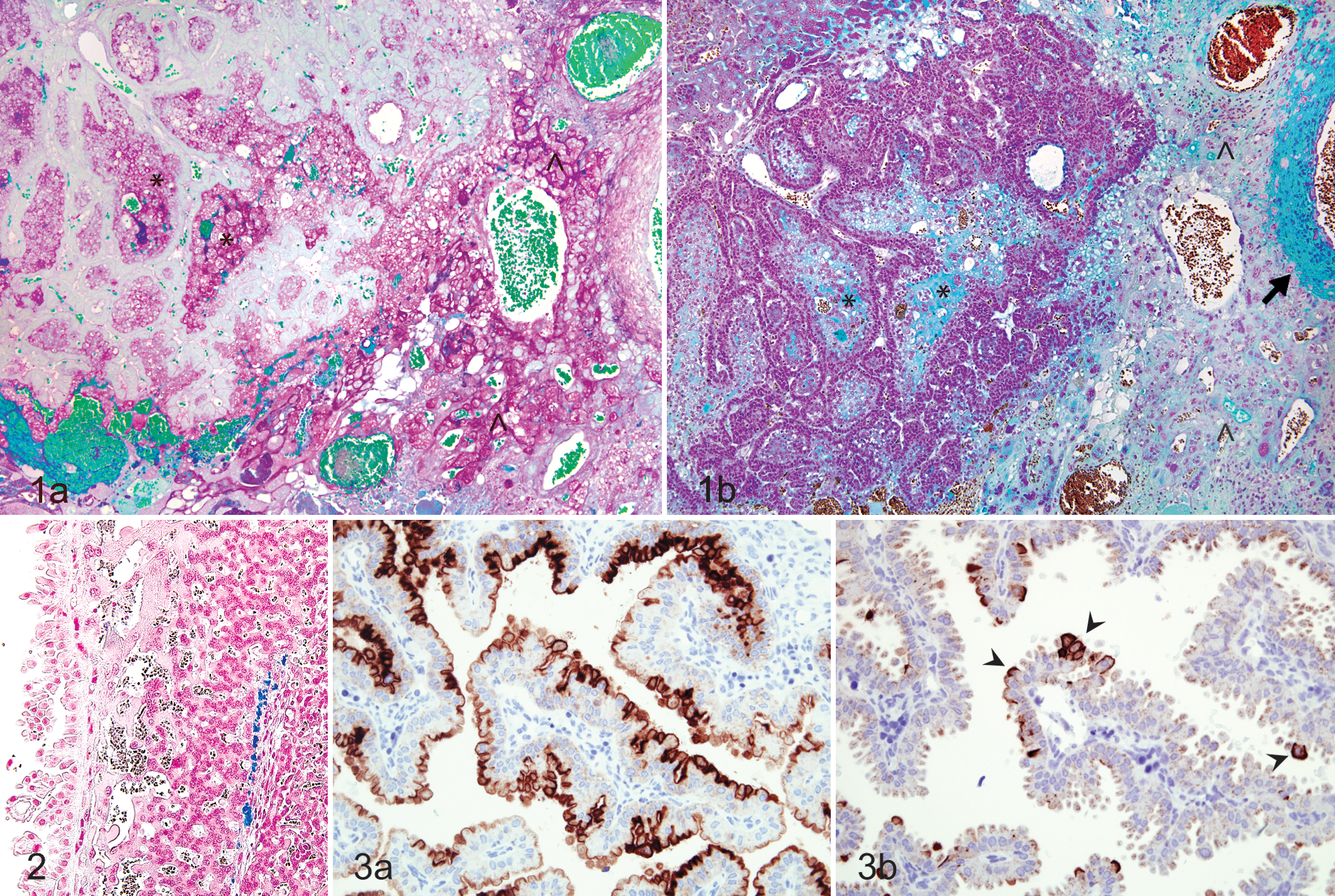
Subplacenta; guinea pig. (a) Subplacental lacunae (*) and junctional zone (^) containing abundant periodic acid–Schiff-positive material. (b) Subplacental lacunae (*) containing moderate amounts of acid mucin and moderate to abundant amounts within the junctional zone (^) and surrounding maternal blood vessels (arrow). Alcian blue, pH 2.5.
Histochemistry and Lectin Histochemistry Findings in Guinea Pig Placenta a
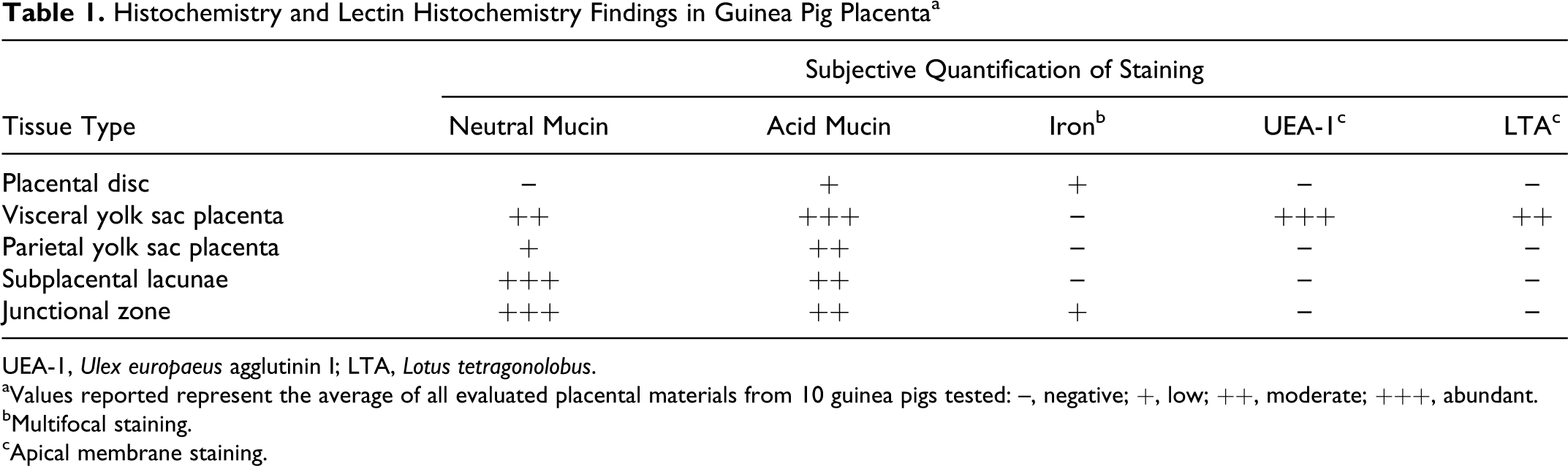
UEA-1, Ulex europaeus agglutinin I; LTA, Lotus tetragonolobus.
aValues reported represent the average of all evaluated placental materials from 10 guinea pigs tested: –, negative; +, low; ++, moderate; +++, abundant.
bMultifocal staining.
cApical membrane staining.
Staining with Alcian blue pH 2.5 revealed moderate amounts of globular extracellular material within subplacental lacunae (Fig. 1b), consistent with acid mucin. Similar material was present in moderate amounts within the junctional zone, most often along the lateral aspect and frequently surrounding the maternal arteries. There was a low level of positive staining in the apical cytoplasm of cells lining vascular channels in the placental labyrinth and along the apical membrane of the parietal yolk sac endoderm. Cells of the visceral yolk sac endoderm were characterized by diffuse, strongly positive apical cytoplasmic and membrane staining (Table 1).
Perl’s staining revealed low to moderate amounts of scattered intracellular and extracellular iron in the placental disc along the junction of the interlobium and the labyrinth (Fig. 2) and surrounding large maternal blood lacunae. Lesser amounts of scattered intracellular and extracellular iron were noted along the basal and, less often, lateral margins of the junctional zone. No iron was noted in the subplacenta or either layers of the yolk sac in the sections evaluated (Table 1).
Lectin Histochemistry
To characterize the spatial distribution of
Growth Assays
None of the biological or chemical preparations yielded microbial growth when plated on MH agar incubated at 37°C aerobically, indicating that the aseptic techniques employed during sample preparation had prevented microbial contamination that would otherwise have confounded interpretation of the results.
At 24 hours, the majority of the pooled placental tissues tested had higher CFU counts in subplacental tissue extracts versus placental disc extracts for both strains of C. jejuni (IA3902 and NCTC 11168), and all samples had considerably higher CFU counts relative to the estimated 0 time value of 5 × 10 5 CFU/ml, suggesting that these tissue extracts support the growth of C. jejuni. At this time point, CFU counts in subplacental tissue extracts ranged from 2.5 × 107 to 3.25 × 108 CFU/ml (mean, 1.9 ± 0.5 × 108 CFU/ml) for IA3902 and from 1.38 × 108 to 3.5 × 108 CFU/ml (mean, 2.61 ± 0.41 × 108 CFU/ml) for NCTC 11168, while CFU counts in placental disc extracts ranged from 1.55 × 107 to 1.98 × 108 CFU/ml (mean, 1.19 ± 0.31 × 108 CFU/ml) for IA3902 and from 2.85 × 107 to 3.59 × 108 CFU/ml (mean, 2.37 ± 0.57 × 108 CFU/ml) for NCTC 11168. The growth multiple for each strain in each pool of tissues was calculated (CFUs in subplacental extracts / CFUs in placental disc extracts), and the mean growth multiples were compared (Fig. 4). Mean growth multiples were > 1 for both strains at 24 hours, indicating greater growth in subplacental extracts at this time point.
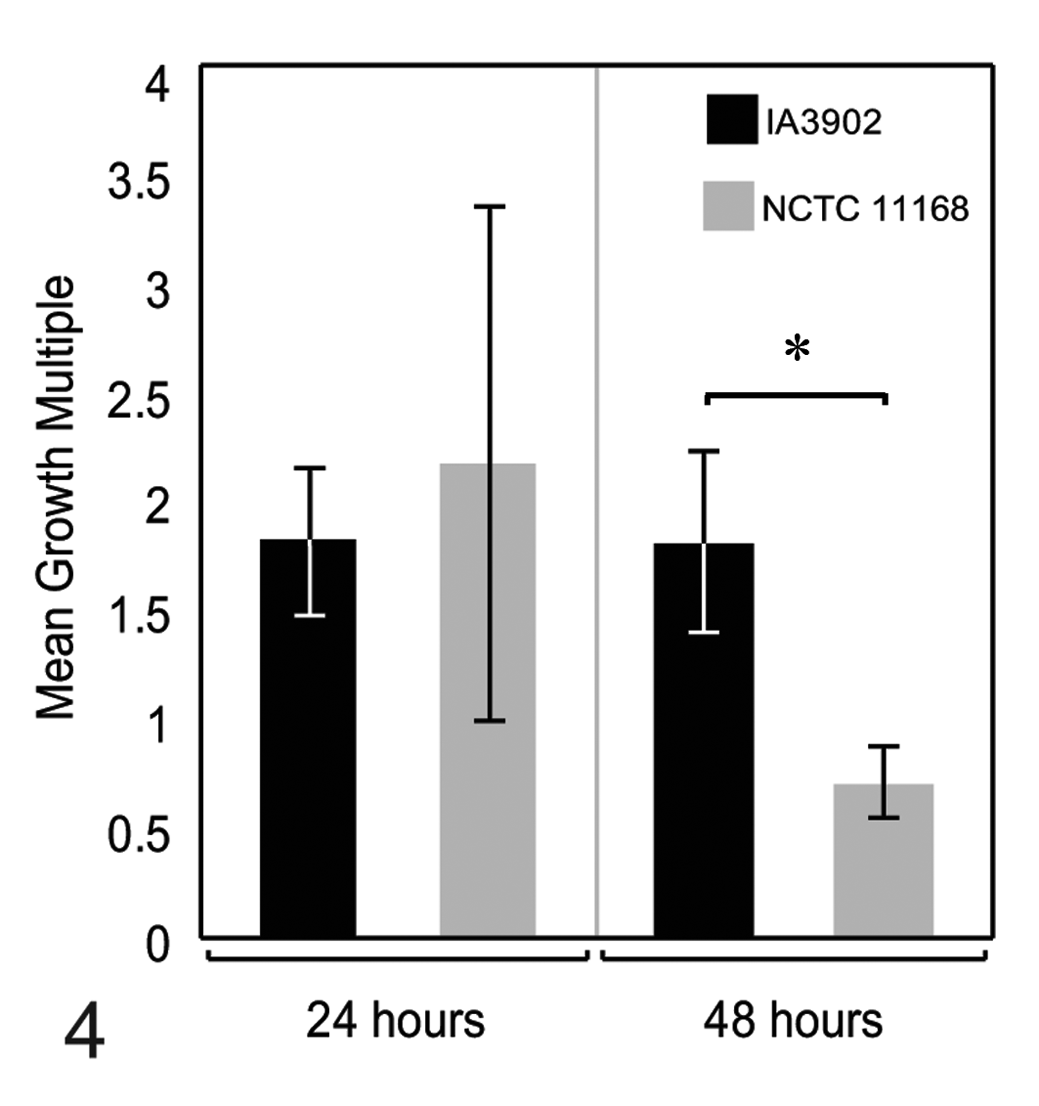
Mean growth multiples (colony-forming units in subplacental extracts / colony-forming units in placental disc extracts) at 24 and 48 hours for Campylobacter jejuni strains IA3902 and NCTC 11168. At 24 hours, the growth multiple for both strains was > 1, indicating greater growth in subplacental extracts versus placental disc extracts. At 48 hours, values differed significantly (* P = .0397), and the growth multiple for NCTC 11168 was < 1, indicating greater growth in placental disc extracts versus subplacental extracts at this time point.
At 48 hours, CFU counts in subplacental tissue extracts ranged from 3.2 × 106 to 2.6 × 107 CFU/ml (mean, 1.2 ± 0.38 × 107 CFU/ml) for IA3902 and from 5.5 × 106 to 4.23 × 107 CFU/ml (mean, 2.58 ± 0.68 × 107 CFU/ml) for NCTC 11168, while CFU counts in placental disc extracts ranged from 1.4 × 106 to 1.25 × 107 CFU/ml (mean, 7.5 ± 1.8 × 106 CFU/ml) for IA3902 and from 2.32 × 107 to 4.84 × 107 CFU/ml (mean, 3.51 ± 0.51 × 107 CFU/ml) for NCTC 11168. The growth multiple for each strain in each pool of tissues was calculated (CFUs in subplacental extracts / CFUs in placental disc extracts), and the mean growth multiples were compared (Fig. 4). The mean growth multiple for IA3902 was > 1, indicating continued greater growth in subplacental extracts at this time point; however, the mean growth multiple for NCTC 11168 was < 1, suggesting greater growth in placental disc extracts for this strain at this time point.
The incidence of bacterial survival was higher in nonpregnant plasma versus pregnant plasma for both strains at 24 hours; however, statistical significance was not attained in the analysis by strain due to low sample numbers associated with sample pooling. CFU counts were generally low overall at this time point and considerably lower than the estimated 0 time value of 5 × 105 CFU/ml, suggesting an antimicrobial effect of the plasma. CFU counts ranged from 0 to 2.48 × 104 CFU/ml (mean, 3.95 ± 2.69 × 103 CFU/ml) for nonpregnant plasma, and the single positive sample in pregnant plasma yielded 3.75 × 103 CFU/ml. Analyzing the incidence of bacterial survival at 24 hours without subdividing by strain revealed a significantly increased incidence in nonpregnant versus pregnant plasma (P = .0076) (Fig. 5). At 48 hours, the incidence of bacterial survival did not differ by pregnancy status for either strain, as organisms were not recovered from the majority of samples at this time point; in addition, CFU counts for those samples where C. jejuni was recovered were extremely low, ranging from 10 to 180 CFU/ml (mean, 56 ± 34.1 CFU/ml).
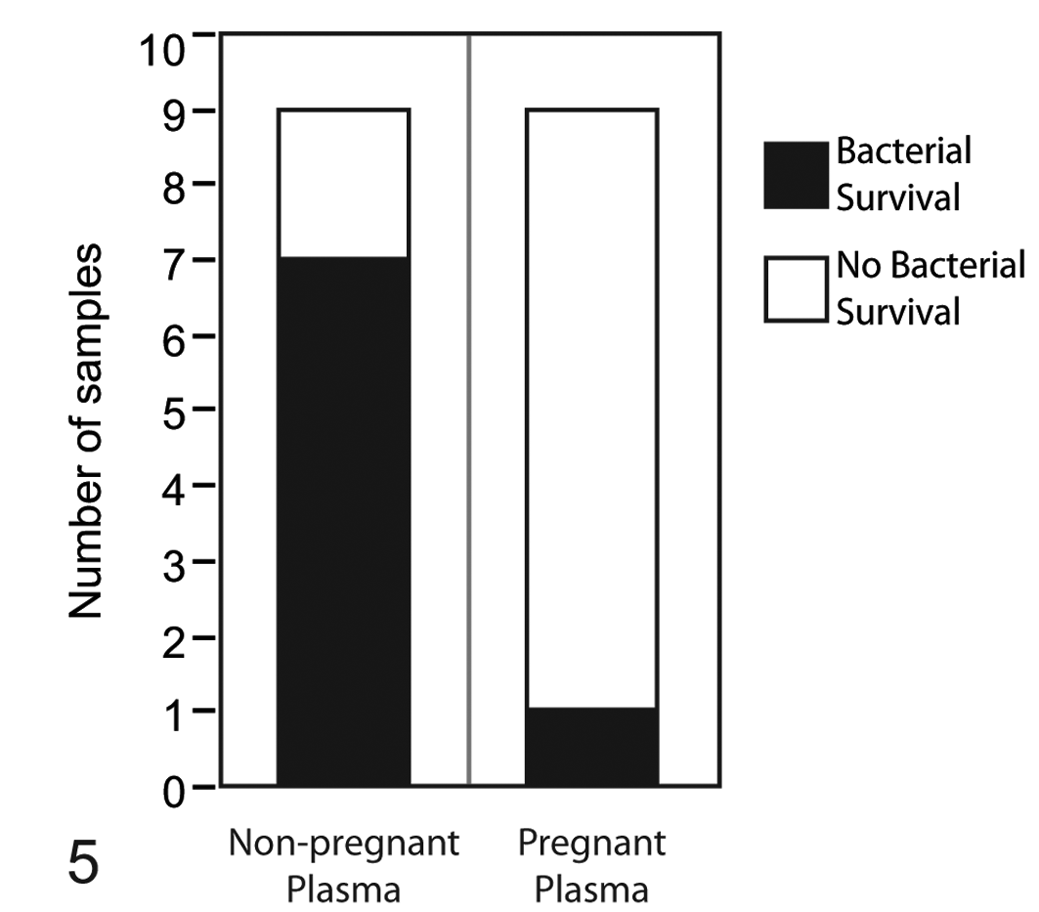
Incidence of bacterial survival at 24 hours in pregnant versus nonpregnant plasma. Shaded areas represent the number of samples from which bacteria were recovered (incidence of bacterial survival) for both Campylobacter jejuni IA3902 and NCTC 11168 combined. Incidence of survival in nonpregnant plasma was significantly greater (P = .0076).
Due to the small volume of the bile samples, only IA3902 was tested in all samples, and CFU counts did not differ significantly by pregnancy status at either 24 or 48 hours (P = .5350 and P = .3844, respectively). Mean CFU counts per milliliter for pregnant versus nonpregnant bile at 24 hours were 7.8 ± 2.7 × 104 and 5.5 ± 2.3 × 104, respectively. Three samples of bile were available for testing NCTC 11168—2 pregnant and 1 nonpregnant—and similar CFU counts were observed as with IA3902; however, statistical analysis was not performed given the low number of samples.
Growth of C. jejuni in the presence of mucin, iron, and
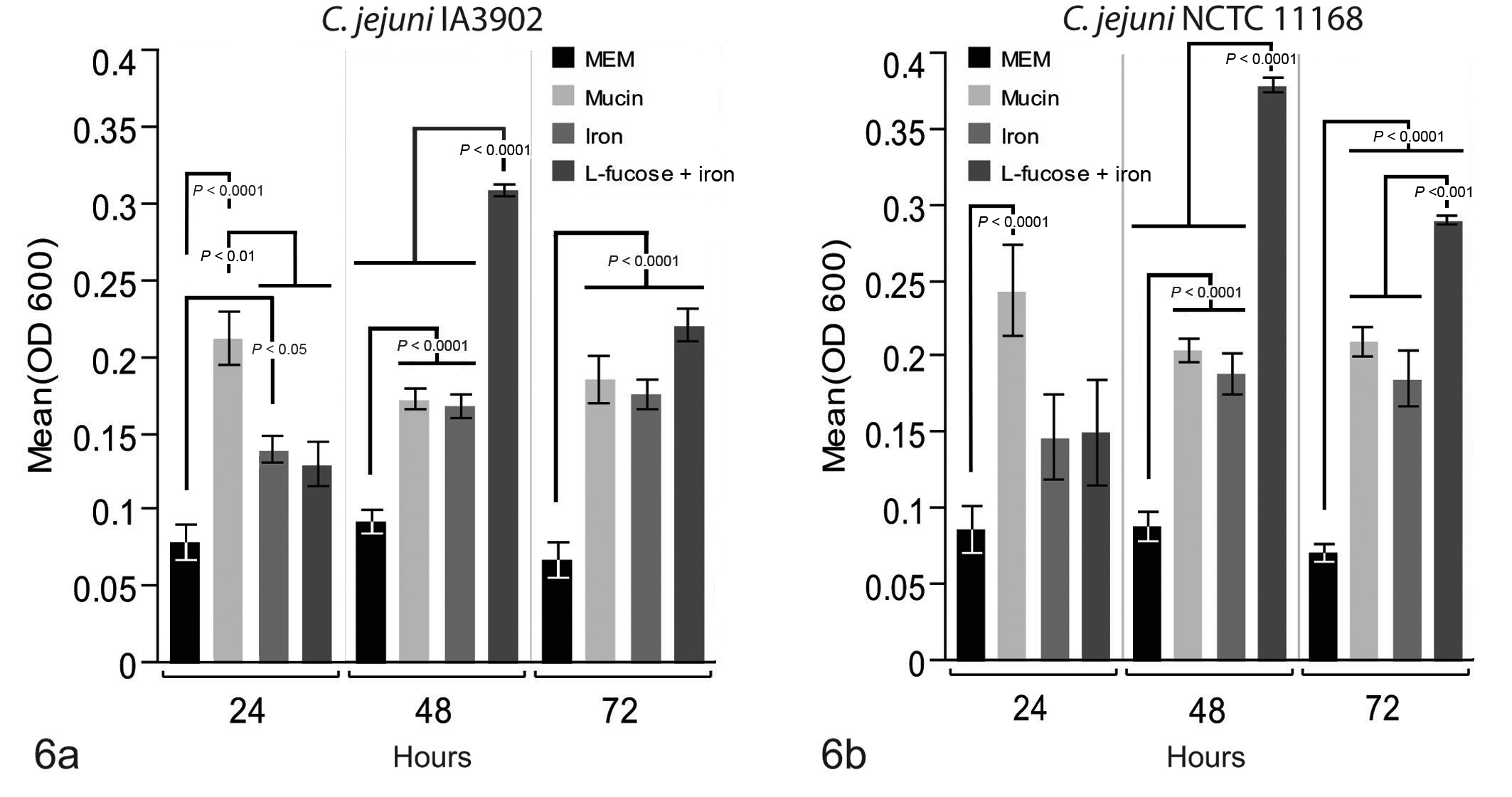
Mean growth (OD600) of Campylobacter jejuni in the presence of individual placental factors at 24, 48, and 72 hours. (a) Growth of IA3902 was significantly higher in the presence of mucin than all other factors at 24 hours, while growth in iron was also greater than minimal essential medium (MEM) control. At 48 hours, growth of IA3902 was significantly higher in the presence of
The growth of C. jejuni in the presence of estradiol + iron, progesterone + iron, and iron alone was compared to that in MEM + DMSO control (Fig. 7). At 24 hours, growth in the presence of all 3 substrates was significantly greater than that in MEM + DMSO control for both IA3902 and NCTC 11168; however, there was no difference between growth in iron alone and iron with the addition of either hormone for either of the C. jejuni strains. At 48 and 72 hours, the results for both C. jejuni strains were similar to the 24-hour time point, with growth in all 3 test substrates significantly higher than that in MEM + DMSO control and with no significant difference observed between any of the iron or hormone combinations for either IA3902 or NCTC 11168.
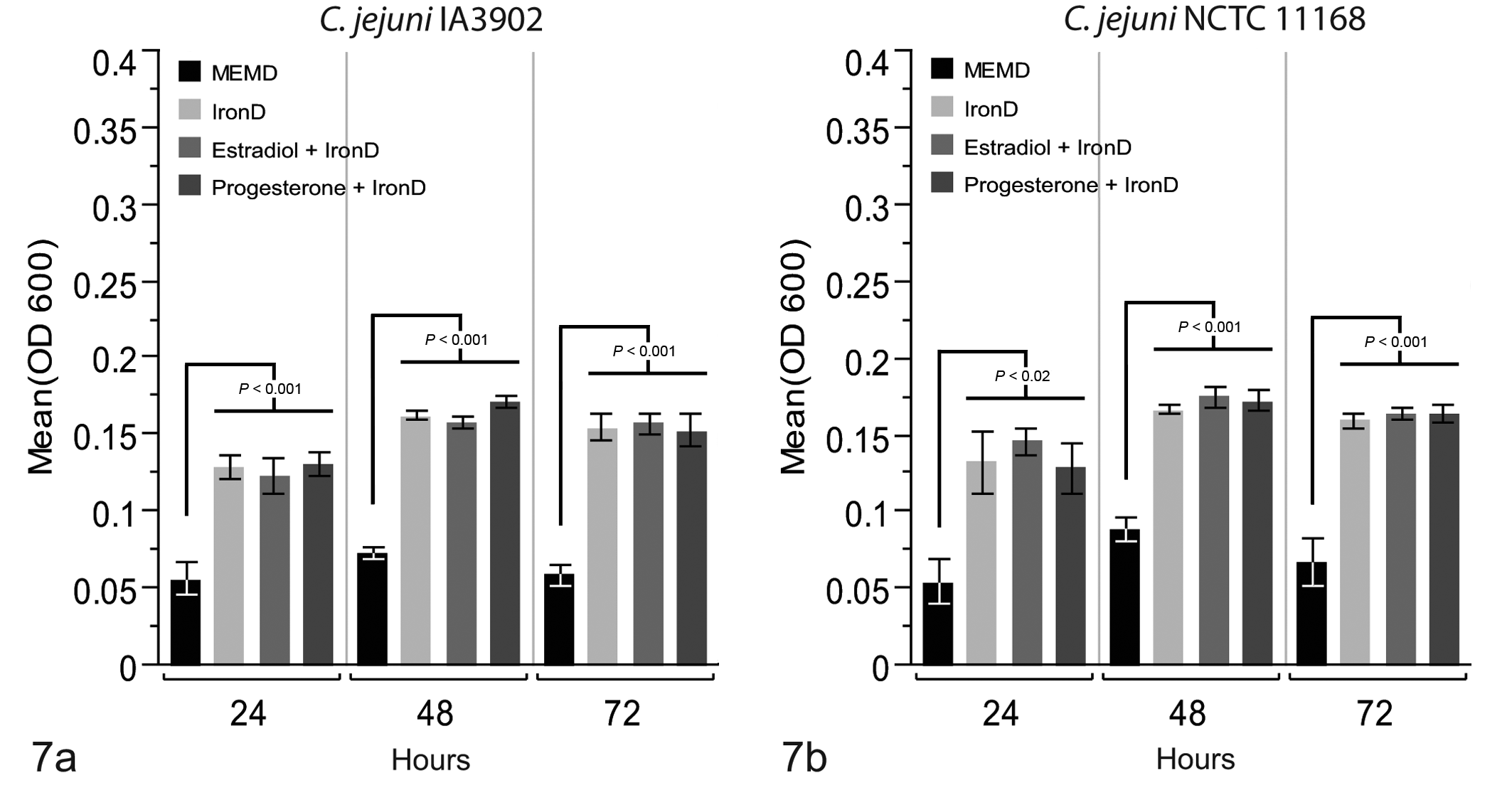
Mean growth (OD600) of Campylobacter jejuni in the presence of individual plasma factors at 24, 48, and 72 hours. (a) At all time points, growth of IA3902 in the presence of all 3 iron-containing treatments was significantly greater than that in minimal essential medium (MEM) + DMSO control; however, no differences were observed with the addition of either progesterone or estradiol. (b) At all time points, growth of NCTC 11168 in the presence of all 3 iron-containing treatments was significantly greater than that in MEM + DMSO control; however, no differences were observed with the addition of either progesterone or estradiol.
Chemotaxis Assays
The results of the filter disc chemotaxis assays are summarized in Table 2. No significant differences were noted between strains in response to any of the tested biological or commercial chemical samples. The only tested chemical with positive chemoattraction was
Chemotactic Responses of Campylobacter jejuni by Strain a
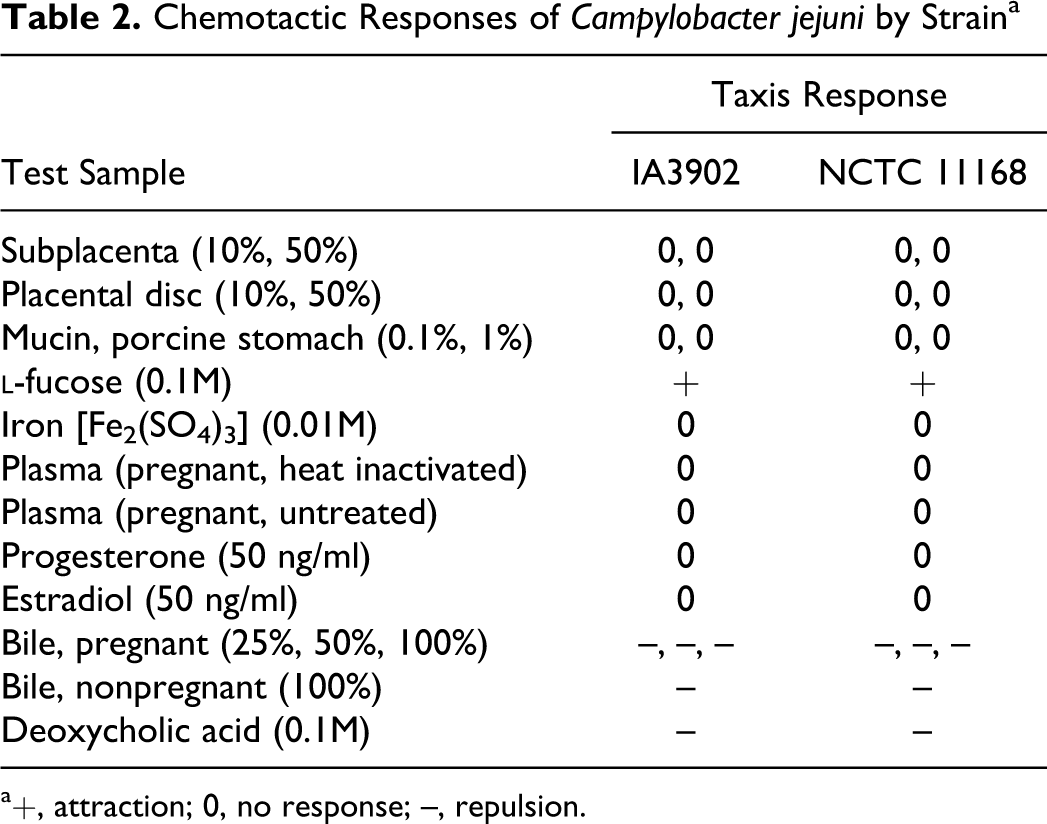
a+, attraction; 0, no response; –, repulsion.
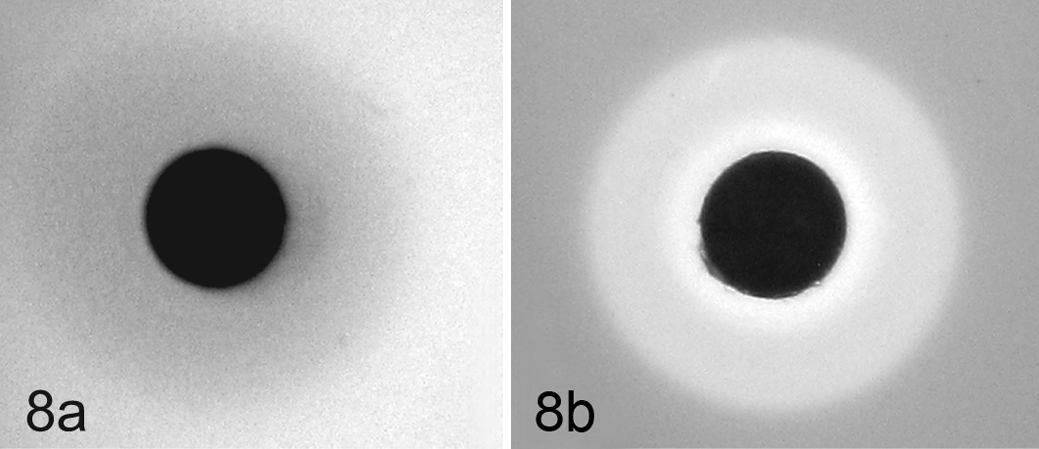
Chemotaxis assays with Campylobacter jejuni suspended in phosphate buffered saline soft agar. Filter discs saturated with test chemicals were placed onto solidified bacterial suspension and migrations zones measured after 4 hours of incubation at 42°C. (a) Strain NCTC 11168 showing chemoattraction toward a filter disc saturated with 0.1M
Discussion
The pathogenesis of ovine campylobacteriosis is complex and includes intestinal invasion, evasion of the host immune response, hematogenous dissemination, and fetoplacental colonization, which ultimately culminates in abortion of the fetus with recovery of Campylobacter in large numbers from expelled placentas. 16 Affected flocks often experience waves of abortions, involving 23.2% of the flock on average. 31 The epidemiology of this disease is further complicated by the high prevalence rate of C. jejuni in domestic flocks, 25 high numbers of healthy sheep with carriage in the bile, 10 and the marked heterogeneity of strains encountered. 22,25 A single clone of C. jejuni was recently recognized as the predominant cause of Campylobacter-associated abortions in the United States, 30 and the pregnant guinea pig was demonstrated as an effective model for studying this highly abortifacient clone. 4 To our knowledge, factors that may drive the fetoplacental tropism of C. jejuni have not been described in any species.
The guinea pig placenta consists of a discoid, labyrinthine main placenta with a separate yolk sac placenta and subplacenta. 19 The subplacenta, a feature unique to histrichognath rodents, is composed of extensive lamellae of cytotrophoblasts lined by a layer of syncytiotrophoblast with large extensively vacuolated lacunae. 6 The subplacenta forms the frontal barrier at the fetomaternal interface and penetrates the decidua, forming the junctional zone. 6 The subplacenta is separated from the main placenta by a layer of fetal mesenchyme, 5 the lateral aspects of which are penetrated by the maternal arteries as they enter the placental disc. 6 Previous immunohistochemical analysis has shown that C. jejuni localizes within this junctional zone and within subplacental trophoblasts following oral infection, 4 a seemingly appropriate location, as this is the site of maternal vascular invasion. In mid- to late gestation, the placenta becomes a major source of progesterone production in guinea pigs, with hormone synthesis occurring in the spongy zone syncytiotrophoblast of the placental disc. 33 The yolk sac placenta is inverted in the guinea pig, with its visceral endoderm facing the uterine lumen and directly contacting the endometrium. 5,17 The functions of this inverted yolk sac include receptor-mediated endocytosis, passage of maternal antibodies, and absorption, degradation, and transfer of maternal serum proteins. 17,20
While there are significant differences between placentation in the guinea pig and the cotyledonary epitheliochorial placentation of sheep, the immunohistochemical localization of C. jejuni within trophoblasts, leukocytes, and perivascular tissues of guinea pigs 4 is consistent with the results of another study 14 of placentas from aborting sheep infected with C. jejuni. Taken together, these findings suggest that pathogenic strains of C. jejuni are capable of infecting trophoblasts regardless of placentation type, and they provide further support for the use of pregnant guinea pigs as a method for investigating pathogenic mechanisms of Campylobacter-associated septic abortion and for the marked fetoplacental tropism observed with strain IA3902.
In the study reported here, the effects of whole tissue extracts from 2 separate regions of the guinea pig placenta were compared, as well as various subcomponents within each region, on the growth and chemotaxis of 2 strains of C. jejuni: IA3902 and NCTC 11168. These 2 strains were demonstrated to have markedly different abortive capacity following oral inoculation. 4 Both strains responded similarly to all in vitro tests, with the exception of the 48-hour growth assay in whole tissue extracts. While both strains grew to higher numbers in the subplacental extracts at 24 hours, only IA3902 maintained this preferential growth at 48 hours, suggesting a potential difference between the 2 strains in their ability to grow within various regions of the guinea pig placenta. This is significant in that previous immunohistochemical studies have shown that IA3902 colonizes the subplacenta and junctional zone while the main placenta is spared.
Previously published studies on the growth and chemotaxis of Campylobacter spp provided potential target compounds to evaluate within the placenta, including mucin,
The results of the 24-hour growth assays revealed that mucin significantly enhanced growth of both strains of C. jejuni versus controls and that for IA3902, this growth was also significantly greater than that in the presence of iron or
Bacterial survival in plasma and bile from pregnant and nonpregnant guinea pigs was compared to determine if a pregnancy-related factor present in these fluids, such as the hormones progesterone and estradiol, might enhance the growth of either strain of C. jejuni. For both strains, the incidence of bacterial survival was higher in plasma from nonpregnant versus pregnant guinea pigs at 24 hours, and neither strain showed enhanced growth in the presence of either progesterone or estradiol in the individual factor assays, suggesting that these hormones do not stimulate the growth of either bacterial strain. This is in contrast to a previous report of enhanced growth in the presence of both hormones with C. rectus 38 and in agreement with previous findings of no growth enhancement in the presence of progesterone with C. fetus subsp venerealis. 37 Both strains survived well in the presence of bile regardless of pregnancy status, again suggesting that soluble factors of pregnancy in the guinea pig and their metabolites do not directly enhance the growth of either C. jejuni IA3902 or NCTC 11168. These findings are of particular note with regard to progesterone, as pregnant guinea pigs have markedly elevated midgestational plasma progesterone levels, with a roughly 56-fold increase versus nonpregnant controls. 2
C. jejuni responds to its environment, undergoing chemotaxis toward a variety of specific chemical substrates
15
and ultimately toward areas that will enhance its growth.
36
The potential chemotactic effects of progesterone were of interest, as guinea pigs and sheep both have increasing placental synthesis of progesterone as pregnancy advances,
27,33
which might serve to explain the late gestational incidence of ovine campylobacteriosis and the marked tropism of this organism for the placenta; however, chemotaxis was not observed for either strain toward progesterone, pregnant plasma, or extracts from the placental disc, suggesting that neither strain responds directly to the presence of progesterone. Marked chemotaxis was observed for both strains toward
Previous chemotaxis studies involving C. jejuni have revealed chemoattraction toward several types of mucin, 15,23,36 which was not confirmed in the present study using swine gastric mucin by either the saturated filter disc or hard-agar plug procedure. While chemotaxis of C. jejuni toward hog gastric mucin has been demonstrated using a hard-agar plug procedure, 15 the strains tested in that study did not include either IA3902 or NCTC 11168, thereby raising the possibility of strain-related differences in chemotaxis. Additionally, 2 recent studies have raised concern about the potential for false-positive results in chemical-in-plug chemotaxis assays, as apparent chemotaxis has been observed in motility-deficient mutant strains of C. jejuni 18 and other bacteria. 21
Gall bladder colonization and fecal shedding may be important in the maintenance of Campylobacter spp in an animal population. In previous studies, IA3902 was isolated from the bile of pregnant and nonpregnant guinea pigs, and C. jejuni was recovered from the bile of healthy sheep at slaughter. 1,3,10 Interestingly, in the present study, bile, regardless of pregnancy status, was strongly chemorepellent for IA3902 and NCTC 11168, and the diameter of the zones of chemorepulsion was concentration dependent where larger diameters correlated with more concentrated bile. This is in contrast to a previously published study where diluted bovine and chicken bile was described as chemoattractive for C. jejuni strains of chicken fecal origin; 15 however, in the same study, the residual bile components after removal of the mucin fraction were strongly chemorepellent for all tested strains. Taken together, these findings suggest that the observed chemorepulsive effect of bile in the present study is due to differences in the composition (eg, mucin) of guinea pig bile relative to that of bovine or chicken origin or due to strain-specific responses of C. jejuni.
In summary, this study demonstrates the presence of specific elements within the guinea pig placenta that enhance growth (iron, mucin,
Footnotes
Acknowledgement
We thank Deborah Moore for her assistance in developing the lectin histochemistry procedures used in this work.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This work was supported by the Iowa Livestock Health Advisory Council (109-05-66), the National Research Initiative Competitive Grants Program from the US Department of Agriculture, National Institute of Food and Agriculture (2010-65110-20419), and the Fort Dodge Animal Health Fellowship.