
Abstract
Toll-like receptors 2 and 4 (TLR2 and TLR4) are well-characterized cell surface receptors that recognize specific pathogen-associated molecular patterns and play an important role in pathogen recognition and activation of the innate immune system. Variable expression of TLR2 and TLR4 has been described in trophoblasts from normal and diseased placentas; yet, there are limited data regarding trophoblast TLR expression in response to specific placental pathogens, and TLR expression in the guinea pig placenta has not been described. The guinea pig is an effective model for Campylobacter-induced abortion of small ruminants, and the authors have shown by immunohistochemistry that C jejuni localizes within syncytiotrophoblasts of the guinea pig subplacenta. The present study was designed to determine if the expression of either TLR2 or TLR4 would be affected in subplacental trophoblasts following infection with C jejuni. Immunohistochemistry for TLR2 and TLR4 was performed on placenta from guinea pigs that aborted following inoculation with C jejuni and from sham-inoculated controls. Quantitative assessment of TLR expression was performed, and mean immunoreactivity for TLR2 was significantly higher in subplacental trophoblasts from animals that aborted compared with uninfected controls (P = .0283), whereas TLR4 expression was not statistically different (P = .5909). These results suggest that abortion in guinea pigs following infection with C jejuni is associated with increased TLR2 expression in subplacental trophoblasts and may reveal a possible role for TLR2 in the pathogenesis of Campylobacter-induced abortion.
Campylobacter spp are a frequent cause of ovine abortion worldwide, 29 and a recent US Department of Agriculture–Animal and Plant Health Inspection Service survey reported that campylobacteriosis was the most common cause of infectious abortion in US flocks. 32 Historically, Campylobacter fetus subsp fetus has been the major isolate from cases of ovine campylobacteriosis. 18 However, two recent studies have shown a marked shift in this relationship, with Campylobacter jejuni becoming the predominant isolate. 13,28 Various molecular typing methods, including pulsed-field gel electrophoresis, have further demonstrated that a single tetracycline-resistant C jejuni clone (SA clone, for sheep abortion) has emerged as the predominant cause associated with ovine abortions in Iowa, South Dakota, Idaho, California, Oregon, and Nevada. 28 Despite the relative frequency of campylobacteriosis, the exact mechanisms of Campylobacter-induced abortion in small ruminants are largely unknown, and the focus of the present study is to gain insight into potential mechanisms underlying the pathogenesis of this disease.
Toll-like receptors (TLRs) are a well-characterized family of pattern recognition receptors that play a key role in innate immune function, and they have been extensively reviewed. 3,19,30,31 In brief, 13 members of the TLR family have been identified in mice 19 —10 of which (TLRs 1–10) have also been identified in cattle, swine, sheep, and humans—and TLR genes have a high degree of sequence similarity between species. 10 TLRs can be grouped on the basis of the characteristics of their ligands and on the location of cellular expression. TLRs 1, 2, 4, 5, and 6 recognize specific pathogen-associated molecular patterns, such as triacyl lipopeptide (TLR1/TLR2), peptidoglycan (TLR2), lipopolysaccharide (TLR4), flagellin (TLR5), and diacyl lipopeptide (TLR6/TLR2), and they are expressed on the cell surface, whereas TLRs 3, 7, 8, and 9 recognize pathogen-associated nucleic acid patterns and are expressed in intracellular compartments, such as endosomes. 19 TLR2 recognizes the most diverse group of ligands, and this is made possible, at least in part, by the formation of heterodimers with TLR1 and TLR6. 34 Members of the TLR family are expressed by a variety of cell types, including those directly involved in host immunity, such as macrophages, neutrophils, and dendritic cells, as well as nonimmune cells such as fibroblasts, endothelial cells, and various epithelial cells. 19
The maternofetal interface has long been considered an immunological paradox because the endometrium must allow for implantation of the allogenic placenta while maintaining adequate immune surveillance against a diverse spectrum of potential pathogens. 24 The role of pattern recognition receptors in this process—particularly, TLRs—has been an active area of research and the subject of a recent review. 1 Immunohistochemistry (IHC) has been used to characterize the expression of TLR2 and TLR4 in the various trophoblast layers from normal and diseased human placentas; 15,20,22,25 however, there are limited data regarding trophoblast TLR expression associated with specific placental pathogens, and TLR expression in the guinea pig placenta has not been described. We have shown by IHC that C jejuni localizes within junctional-zone trophoblasts of the guinea pig subplacenta, 6 and we hypothesize that expression of one or more TLRs will be increased in subplacental trophoblasts following infection with C jejuni IA 3902 (a clinical sheep abortion isolate belonging to the SA clone).
Previous studies have revealed that TLR2 and TLR4, but not TLR5, are capable of binding pathogen-associated molecular patterns of Campylobacter spp. Specifically, the flagellin of C jejuni does not effectively bind TLR5; 4,33 C jejuni and C coli induce interleukin 8 (IL-8) secretion by polarized human intestinal epithelial cells in a TLR-mediated manner; 35 C rectus infection increases TLR4 expression in murine placentas; 5 C jejuni surface polysaccharides induce IL-6 production in human intestinal epithelial cells via TLR2; 14 and lysed C jejuni effectively activates chicken and human TLR2 and TLR4. 12 Accordingly, in the study reported here, we used IHC to evaluate the expression of TLR2 and TLR4 in guinea pig subplacental trophoblasts following infection with C jejuni IA 3902 to delineate the involvement of these TLRs in response to this specific placental pathogen.
Materials and Methods
Sample Collection
Blocks of formalin-fixed, paraffin-embedded guinea pig placenta were obtained from animals used in previous experiments 6 in which animals were inoculated orally with C jejuni IA 3902 at approximately 4 weeks of gestation, with abortions occurring between 4 and 14 days postinoculation. Aborted placentas (44 total) were obtained from 12 orally inoculated guinea pigs. Pooled placental tissues from each dam were culture positive for C jejuni, indicating that at least one of the placental samples from each orally challenged guinea pig was infected. Control placentas (28 total) were obtained from 9 sham-inoculated animals at 21 days postinoculation.
Histologic Evaluation
Hematoxylin and eosin–stained sections of each placenta were evaluated to determine the presence or absence of suppurative placentitis as defined by the presence of infiltrating neutrophils within the junctional zone between the subplacenta and decidua.
Immunohistochemistry
Formalin-fixed, paraffin-embedded guinea pig placental samples were prepared for IHC as previously described. 6 For antigen retrieval, sections were treated with 0.1% protease in a Tris buffer (pH 7.6) at 37°C for 10 minutes, followed by three rinses in phosphate buffered saline solution before placement in an automated cell staining system (BioGenex, San Ramon, CA). Sections were then incubated in 10% neutral goat serum at 22°C for 20 minutes to inhibit nonspecific binding. Primary antibodies consisted of commercially available affinity-purified rabbit antihuman TLR2 and affinity-purified rabbit antihuman TLR4 (Rockland, Gilbertsville, PA) and were used at dilutions of 1:500 and 1:300, respectively. Rabbit antihuman TLR2 and TLR4 primary antibodies have been shown to be cross-reactive in guinea pig tissues. 17 Slides were incubated at 22°C for 60 minutes (TLR2) and 120 minutes (TLR4), followed by rinsing in a bath of phosphate buffered saline solution for 5 minutes. The secondary antibody (BioGenex), horse radish peroxidase–streptavidin conjugate (Invitrogen, Carlsbad, CA), chromogen (Vector, Burlingame, CA), and counterstaining steps were performed as previously described. 6 Positive control samples consisted of guinea pig spleen and lung tissues for evaluation of receptor expression on phagocytes. Negative controls were prepared from the same blocks and consisted of sections in which the primary antibody was omitted and sections in which normal (preimmune) rabbit serum was applied in place of the primary antibody.
IHC for C jejuni was also performed on all aborted placentas as previously described. 6
Image Analysis
IHC-stained sections were examined with an Olympus BX60 light microscope (Olympus, Tokyo, Japan) and RGB color digital images were recorded with an Olympus DP70 camera (Olympus) with 2040- × 1536-bit resolution and 24-bit depth. Three magnification images (40×) were obtained for each subplacenta and included one image taken from each lateral aspect of the junctional zone and one from the area just below the central excavation. The recorded microscopic fields were oriented such that subplacental trophoblasts filled as much of the field as possible while excluding any inflammatory exudate present in the junctional zone.
To quantify the level of TLR protein expression in subplacental trophoblasts, all digital images were analyzed with ImageJ image analysis software (National Institutes of Health, Bethesda, MD) and a color deconvolution (stain separation) plug-in implementing methods previously described. 27 Using the plug-in, the red color (positive immunohistochemical staining) was isolated in each image and then converted to a black-and-white image and set to a common threshold. Thresholds of 175 and 200 were selected for TLR2 and TLR4, respectively. The percentage area of each image exhibiting the selected threshold level of staining was then quantified. The mean percentage of the stained area was then determined for the three images from each placenta, and the average staining of all placentas from each individual guinea pig was calculated and compared.
Statistical Analysis
SAS 9.2 was used to perform independent-sample t tests for a difference in mean TLR protein expression between groups. Statistical significance was determined as P ≤ .05.
Results
Histologic Findings in Affected Placentas
Placentas from aborting guinea pigs were typically characterized by one or more of the following changes: congestion of the placental labyrinth, multifocal necrosis of the placental disc, and multifocal to coalescing necrosis of subplacental trophoblasts (Fig. 1 ) that was often associated with neutrophilic infiltration. The placental junctional zone in at least one placenta from all 12 aborting guinea pigs was infiltrated with neutrophils; however, not every placenta from each aborting animal was equally affected. This was most notable in guinea pigs with higher fetal numbers, as expected, and placentas from individual fetuses occasionally lacked histologic lesions. In total, 88.6% of placentas from aborting guinea pigs (39 of 44) had evidence of suppurative placentitis. IHC for C jejuni revealed that 65.9% of placentas from these aborting animals (29 of 44) had direct evidence of bacterial infection within subplacental trophoblasts and phagocytes in the placental junctional zone, and 100% of IHC-positive placentas (29 of 29) had suppurative placentitis.
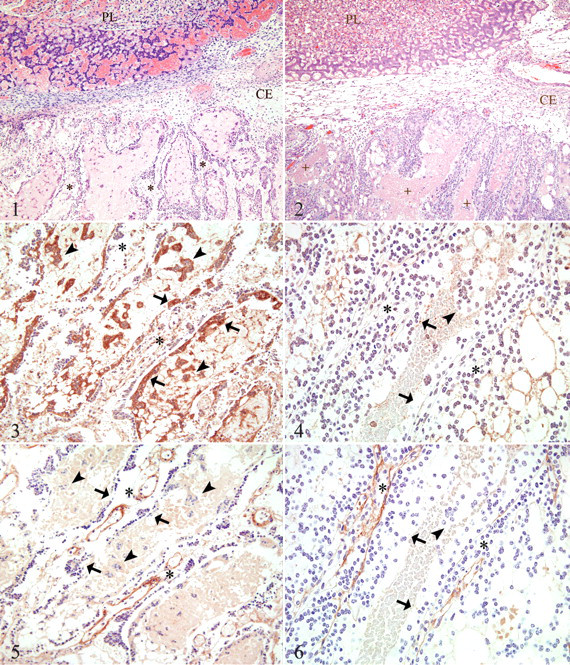
Placentas from sham-inoculated controls lacked significant lesions (Fig. 2 ). Multifocally, there were small areas in which subplacental trophoblasts (1) appeared hypereosinophilic with pyknotic nuclei and (2) often surrounded accumulations of scant to moderate amounts of eosinophilic cellular and karyorrhectic debris (necrosis) admixed with basophilic granular material (mineral); however, these areas were not associated with neutrophilic infiltration and were generally limited to the lateral aspects of the junctional zone.
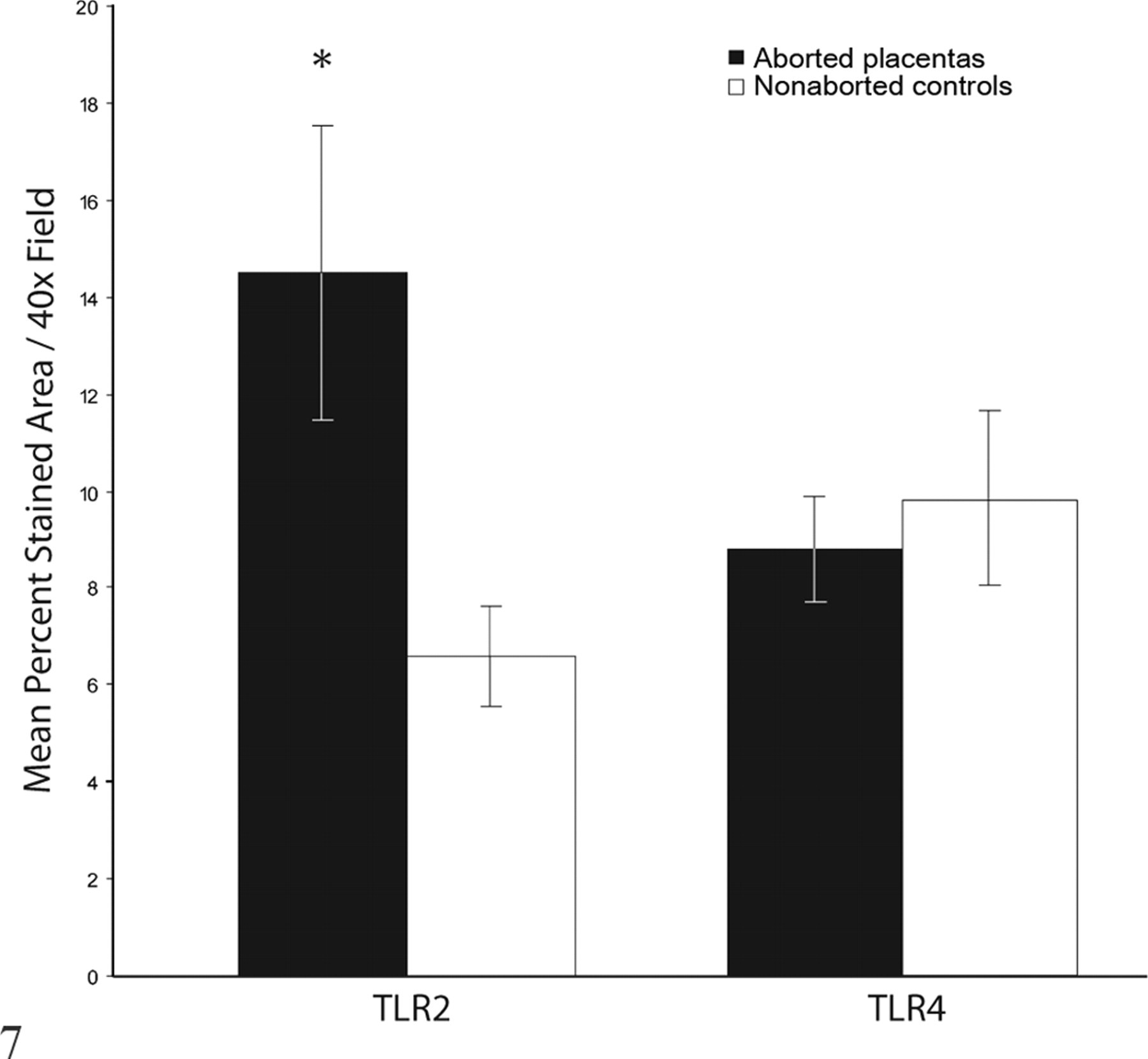
Percentage area of subplacental photomicrographs corresponding to positive staining for Toll-like receptors 2 and 4 (TLR2 and TLR4) per 40× field (mean ± SEM). Values represent mean immunoreactivity for all placentas from 12 aborting guinea pigs (44 placentas) and 9 nonaborted controls (28 placentas). Asterisk indicates that values differ significantly (P = .0283).
Expression of TLR2
IHC of aborted placentas, as compared with controls, revealed a moderate increase in TLR2 immunoreactivity within trophoblasts of the main placenta; however, strong positive immunoreactivity was a consistent feature of subplacental syncytiotrophoblast cells and syncytial islands within lacunae, whereas the cytotrophoblast layer lacked significant staining (Fig. 3 ). In sham-inoculated control tissues, a low level of diffuse cytoplasmic immunoreactivity for TLR2 was evident in trophoblasts of the interlobium of the main placenta, whereas the syncytiotrophoblast and cytotrophoblast cells of the subplacenta were generally unstained (Fig. 4 ). Positive immunoreactivity was also observed in neutrophils and macrophages within control tissues, whereas negative controls were nonreactive (not shown).
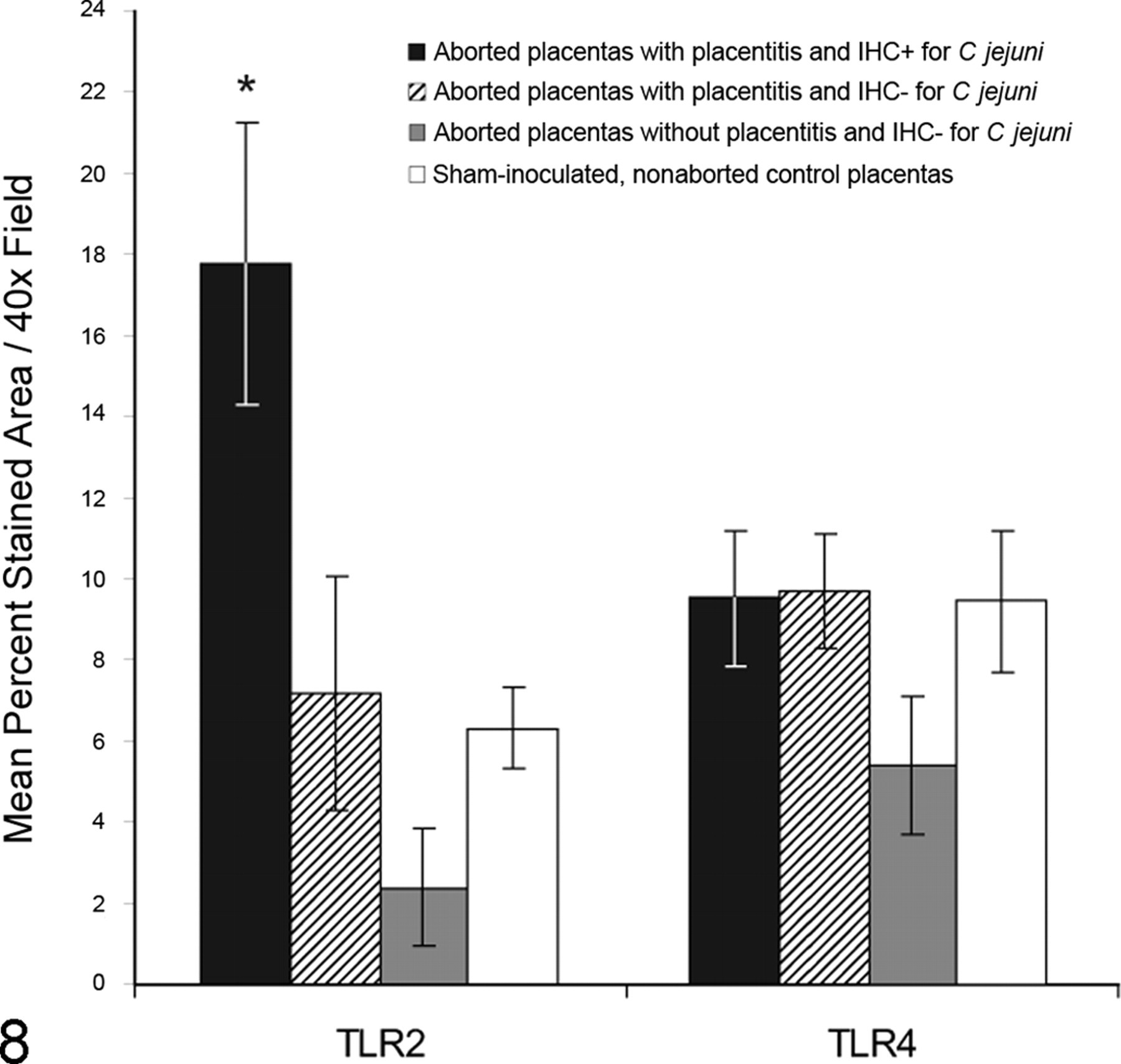
Percentage area of subplacental photomicrographs corresponding to positive staining for Toll-like receptors 2 and 4 (TLR2 and TLR4) per 40× field (mean ± SEM). Values represent mean immunoreactivity for all aborted placentas immunohistochemistry positive (IHC+) for Campylobacter jejuni (9 guinea pigs, 30 placentas), aborted placentas with placentitis but immunohistochemistry negative (IHC–) for C jejuni (5 guinea pigs, 9 placentas), aborted placentas without placentitis (3 guinea pigs, 5 placentas), and nonaborted controls (9 guinea pigs, 28 placentas). Asterisk indicates that value differs significantly from controls (P = .0108).
Expression of TLR4
IHC of aborted placentas revealed moderate TLR4 immunoreactivity within trophoblasts of the main placenta and in the mesenchyme and surrounding vascular spaces of subplacental lamellae. A low level of positive immunoreactivity was an inconsistent feature of subplacental syncytiotrophoblast cells, whereas the adjacent cytotrophoblast layer lacked significant staining (Fig. 5 ). IHC of sham-inoculated control tissues revealed scant cytoplasmic immunoreactivity for TLR4 in trophoblasts of the interlobium, moderate immunoreactivity within the mesenchyme and surrounding vascular spaces in subplacental lamellae, and minimal to no immunoreactivity within subplacental syncytiotrophoblast and cytotrophoblast cells (Fig. 6 ). Strong positive immunoreactivity was also observed in neutrophils and macrophages within control tissues, whereas negative controls were nonreactive (not shown).
Image Analysis of Subplacental Immunoreactivity
TLR2 immunoreactivity was significantly higher in subplacental trophoblasts from aborted placentas compared with nonaborted, sham-inoculated controls (P = .0283). Figure 7 summarizes the mean percentage area corresponding to positive immunohistochemical staining for images obtained from each group. This difference was even more significant when we compared mean TLR2 immunoreactivity between controls and only those placentas with suppurative placentitis and positive IHC for C jejuni (P = .0108), whereas mean immunoreactivity of (1) placentas with evidence of suppurative placentitis but negative IHC for C jejuni and (2) those aborted placentas that lacked placentitis did not differ significantly from controls (P = .7350 and P = .0724, respectively). Figure 8 summarizes these results.
TLR4 immunoreactivity did not differ significantly between aborted placentas and controls (P = .5909). Figure 7 summarizes the mean percentage area corresponding to positive immunohistochemical staining for images obtained from each group. Additionally, mean TLR4 immunoreactivity did not differ significantly between control placentas and (1) only those placentas from aborting animals that had suppurative placentitis and positive IHC for C jejuni (P = .9830), (2) those with only suppurative placentitis (P = .9321), and (3) only those from aborting guinea pigs that lacked placentitis (P = .2380). Figure 8 summarizes these results.
Discussion
Campylobacteriosis is a well-recognized cause of small ruminant abortion worldwide; however, the molecular mechanisms involved in this process are not known. We recently showed using IHC that, following oral inoculation, C jejuni IA 3902 localizes within syncytiotrophoblast cells of the guinea pig subplacenta, 6 and in the present study, 65.9% of placentas from infected guinea pigs (29 of 44) had evidence of C jejuni antigen by IHC, consistent with the 71.4% noted in our previous work. To further explore potential mechanisms associated with Campylobacter-induced abortion, we performed IHC for TLR2 and TLR4 on these placental tissues.
The guinea pig placenta consists of a disclike, labyrinthine chorioallantoic main placenta with separate subplacenta and yolk sac placenta. 16 The subplacenta is unique to hystricognath rodents, and its histological, histochemical, and ultrastructural properties have been thoroughly described elsewhere. 11,23,26 In brief, the subplacenta is separated from the main placenta by a thin layer of fetal mesenchyme, 9 and the distal segments of the uteroplacental arteries enter the main placenta through the periphery of this junction. 11 It is in this location that we consistently observe C jejuni via IHC, which seems logical because this is the site of placental vascular invasion and would be the first location reached during episodes of bacteremia. The subplacenta is composed of lamellae of mesenchyme that support a folded layer of cytotrophoblast cells that give rise to a syncytiotrophoblast layer along the decidual surface. 11 As gestation progresses, syncytiotrophoblasts form extensive lacunae that contain abundant eosinophilic material and syncytial islands. 11 The interface between the subplacenta and decidua is known as the junctional zone, 11 and our current and previous immunohistochemical findings indicate that, in pregnant guinea pigs, C jejuni IA 3902 localizes within subplacental trophoblasts at the periphery of this junctional zone near the site of placental vascular invasion, whereas the main placenta is spared. For this reason, we focused our efforts in the present study on TLR expression within subplacental trophoblasts.
In the study reported here, we observed markedly increased immunoreactivity for TLR2 protein in the syncytiotrophoblast layer of the subplacenta from guinea pigs aborting following infection with C jejuni versus sham-inoculated controls. This occurred without significant change in the underlying cytotrophoblast cells, which is significant because the syncytiotrophoblast constitutes the frontal barrier of the placenta at the maternofetal interface. Because the guinea pig generally has multiple offspring in each litter, during any bacteremic event, there may likely be an uneven distribution of organisms seeding fetoplacental units. As such, we further subdivided the placentas from aborting guinea pigs on the basis of positive IHC for C jejuni, placentitis without IHC for C jejuni, and the absence of placentitis. Previous studies have shown that IHC is less sensitive than culture in detecting Campylobacter spp within placental tissues; 6,7 thus, it is possible that all inflamed placentas were infected with C jejuni but that campylobacter numbers in IHC-negative inflamed placentas were substantially lower than IHC-positive placentas. Within the subdivided groups, placentas with placentitis and positive IHC for C jejuni had significantly greater levels of TLR2 immunoreactivity when compared with controls, whereas the placentas from aborting animals that were inflamed but IHC negative or that lacked placentitis did not differ from controls. These findings are intriguing and suggest an association between the presence of C jejuni and increased expression of TLR2. Although these results do not give information with regard to the mechanism of increased TLR2 expression, the presence of increased TLR2 protein suggests a potential role in the disease pathogenesis. A recent study in human intestinal epithelial cells showed that C jejuni surface polysaccharides induce IL-6 production via TLR2, 14 and studies in first-trimester human trophoblasts have revealed that TLR2 ligation induces increased production of IL-6 and IL-8 as well as increased trophoblast apoptosis. 2 The combined presence of suppurative placentitis and increased TLR2 expression in the infected tissues of this report is consistent with this type of proinflammatory response; however, further studies are required (1) to determine if the increased expression of TLR2 is a direct effect of the organism or a downstream effect of placental inflammation and (2) to elucidate the exact cytokine response that occurs in guinea pig trophoblasts in response to C jejuni.
In contrast to our findings with TLR2, TLR4 protein expression in subplacental trophoblasts did not differ significantly between controls and placentas from aborting animals overall or from any of the subdivided groups. These results suggest that altered TLR4 expression is not associated with Campylobacter-induced abortion in guinea pigs. Although these results appear to conflict with the findings of Arce et al, 5 whose study revealed increased TLR4 in murine placentas exposed to C rectus, 5 our study is significantly different from theirs. First, unlike caviomorphs, mice lack a subplacenta, 8 and TLR4 IHC in the study by Arce et al assessed expression in trophoblasts from the placental labyrinth. Second, abortion was not the endpoint of their murine study; thus, the level of TLR4 expression at the point of pregnancy termination was not assessed. Additionally, the murine study was designed to assess the placental response to chronic oral exposure to a periodontal pathogen (C rectus), whereas the present study assessed the placental response to acute infection with a highly abortifacient strain (C jejuni). Finally, Arce et al evaluated TLR4 only, so it remains to be seen what, if any, TLR2 response might occur in murine placentas following infection with C rectus. It is also conceivable that both receptors may be elevated at some point throughout the disease process, given that time-dependent interdependence between TLR4 and TLR2 has been described. 21
Limitations of this study include its retrospective nature and the lack of a specific time-matched control placenta for each aborted placenta. The presence of noninflamed placentas within the pool of aborted tissues did, however, serve as an internal control. Furthermore, the immunoreactivity of these tissues was similar to sham-inoculated control tissues; as such, the constitutive expression of TLR2 and TLR4 in the guinea pig subplacenta did not significantly vary throughout the period studied. In addition, the relevance of these findings to ovine campylobacteriosis is uncertain; however, a recent study of obesity-induced placental inflammation in ewes revealed elevated mRNA expression for TLR2 and TLR4 with a concurrent increase in proinflammatory cytokines, 36 and these results support a role for TLRs in the response of ovine placenta to inflammatory stimuli.
In summary, whereas TLR4 immunoreactivity was essentially unchanged, we observed increased TLR2 immunoreactivity in subplacental syncytiotrophoblast cells from aborting guinea pigs following oral inoculation with C jejuni, and these changes were most prominent in syncytiotrophoblasts from placentas with direct evidence of C jejuni by IHC. Identification of these changes in subplacental syncytiotrophoblast cells is significant because these cells form the frontal barrier at the maternofetal interface, which therefore suggests a role for TLRs in bacterial sensing and the subsequent inflammatory response generated during septic abortion due to C jejuni. To our knowledge, this report is the first of TLR expression in the guinea pig placenta and the first to describe trophoblast TLR expression associated with a specific well-documented placental pathogen. These findings warrant further investigation to determine the exact role of TLR2 expression in the pathogenesis of C jejuni–associated abortion.
Footnotes
Acknowledgements
We would like to thank Deborah Moore for her assistance in developing the immunohistochemistry procedures used in this work.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
This study was supported by the Iowa Livestock Health Advisory Council (grant No. 109-05-66) and the Iowa State University CVM Summer Scholars Research Program.