
Abstract
Group C streptococci are highly contagious pyogenic bacteria responsible for respiratory tract, lymph node, urogenital tract, and wound infections. Wild-type strains of Streptococcus equi ssp equi (S. equi) and Streptococcus equi ssp zooepidemicus (S. zoo) as well as a commercially available modified live vaccine strain of S. equi were evaluated for virulence in zebrafish. Survival times, histologic lesions, and relative gene expression were compared among groups. Based on the intramuscular route of infection, significantly shorter survival times were observed in fish infected with wild-type strain when compared to modified live vaccine and S. zoo strains. Histologically, S. zoo–infected fish demonstrated a marked increase in inflammatory infiltrates (predominantly macrophages) at the site of infection, as well as increased cellularity in the spleen and renal interstitium. In contrast, minimal cellular immune response was observed in S. equi–injected fish with local tissue necrosis and edema predominating. Based on whole comparative genomic hybridization, increased transcription of positive acute-phase proteins, coagulation factors, and antimicrobial peptides were observed in S. equi–injected fish relative to S. zoo–injected fish, while mediators of cellular inflammation, including CXC chemokines and granulin, were upregulated in S. zoo–injected fish relative to S. equi–injected fish. In a screen of 11 clinical isolates, S. equi strains with a single nucleotide deletion in the upstream region of szp, a known virulence factor of streptococci, were found to be significantly attenuated in zebrafish. These collective findings underscore the value of the zebrafish as a model of streptococcal pathogenesis.
Group C streptococci include the closely related Streptococcus equi ssp equi (S. equi) and Streptococcus equi ssp zooepidemicus (S. zoo). S. equi is the etiologic agent of equine strangles, a highly contagious lymphadenitis of horses. Strangles is an ancient disease of horses that continues to generate significant losses for breeders, the racing industry, and pleasure horse owners. S. zoo is responsible for respiratory infections in many domestic species, including horses, dogs, and pigs, among others. 22 Losses due to uterine infection as well as joint and wound infections in horses are significant.
The zebrafish (Danio rerio) is emerging as a useful animal model for the study of many disease processes, including bacterial pathogenesis. As an animal model, zebrafish have several advantages: small size, low cost, and an advanced and pliable immune system. 10,15 –18 The zebrafish has been successfully utilized in the study of other distantly related streptococci; however, a thorough evaluation of the zebrafish host response to group C streptococci has not been performed. 17,19,27 Current methods of investigating the differences in group C streptococcal pathogenesis in nonequine species are confined to mouse and less commonly used rat models. 5 –8,25 Rodent models utilizing intramuscular (IM) or intraperitoneal (IP) routes of injection have been successfully used to measure streptococcal virulence and humoral response to vaccination. 6,7,25 The intranasal route of infection with S. equi has also been investigated in mice, with 6 of 30 mice (20%) developing a mild regional lymphadenitis reminiscent of the natural disease. 5 Despite the availability of these models, an inexpensive, high-throughput model utilizing a less sentient animal species is desirable for rapid screening of large numbers of isolates or vaccine candidates. We hypothesize the following: First, the group C streptococci will produce significant disease in zebrafish at relatively low doses; second, infection with the wild-type S. equi vaccine progenitor strain (WT) will result in shorter survival times in infected fish compared to fish injected with either isolates of S. zoo or the commercially available modified live vaccine strain of S. equi (Vx); third, histologic and gene expression differences will be observed in fish injected with strains of S. equi and those injected with S. zoo; and, fourth, the model will successfully detect attenuation in 11 laboratory strains.
Materials and Methods
Bacterial Strains
The Vx (Pinnacle IN, Fort Dodge Animal Health Division of Pfizer Animal Health, Kalamazoo, MI), WT (CF32, ATCC 53185), and laboratory S. zoo strain (LBC50) were used to develop the model. Eleven clinical isolates of S. equi (A–K) identified with the Biolog ID System (Hayward, California) by the University of Illinois Veterinary Diagnostic Laboratory were also evaluated using the IM route of injection. These isolates were evaluated genotypically using pulsed field gel electrophoresis (PFGE) and DNA sequence analysis of szp and seM and phenotypically with the Biolog Phenotypic MicroArray analysis. 14
Pulsed Field Gel Electrophoresis
PFGE was performed using the restriction enzymes Sma I, Not I, and Sfi I as previously described. 14 Digestion patterns identical to those identified in the Pinnacle IN vaccine were designated Vx, while patterns not identical to the vaccine strain received a number designation. With a combination of these 3 enzymes, most isolates were identified as WT or Vx; however, the WT progenitor strain had a PFGE pattern that was indistinguishable from the Vx pattern using all 3 enzymes.
szp Polymorphisms
A single guanidine (G) deletion in the upstream region of szp has been identified in the Vx strain, which is not present in the WT progenitor. The szp gene, including this upstream region, was amplified and sequenced. Polymerase chain reaction (PCR) was performed with Phusion polymerase (New England Biolabs, Boston, Massachusetts) according to the manufacturer’s directions. Primer sequences were as follows: szp-1700F 5′ GCC AAT TGA GTC AAG ACA ATC A 3′ and szp-1700R 5′ GTG AGC AAG GCT TAG TCT CTC C 3′. The amplicons were subjected to DNA sequencing at the W. M. Keck Center for Comparative and Functional Genomics at the University of Illinois at Urbana-Champaign using the following primers: szp-SEQ-F 5′ GCT CTA GCG ATA TAG CAG GG 3′ and szp-SEQ-R 5′ GCT GAT ACA GCT GCC GCA CC 3′. Sequence alignments were performed using DNAsis software.
seM Polymorphisms
Single-nucleotide polymorphisms (SNPs) in the M protein seM gene have been identified and exploited to group-related isolates of S. equi. 1,12 A SNP at the 63rd codon was used for genotyping isolates in this experiment. PCR reactions were performed with Phusion polymerase according to the manufacturer’s directions. Primer sequences were as follows: seM-2200F 5′ AAG GGC AAC TAC TTC CTC TCC T 3′ and seM-2200R 5′ TTC CAA AAG CTT TCT GTC ACC T 3′. The amplicons were subjected to DNA sequencing at the W. M. Keck Center for Comparative and Functional Genomics at the University of Illinois at Urbana-Champaign using the following primers: seM-500F 5′ TGT TGA TCT TGA TAA AGT TTT GT 3′ and seM-500R 5′ CGG TGC AGC ATC AGT ATT AGT T 3′. Sequence alignments were performed using DNAsis software.
Biolog Phenotypic MicroArray
Isolates were grown on BUG Agar plates overnight at 37°C in 5% CO2, placed into GP inoculating fluid (Biolog), and then dispensed into a GP Biolog Microphenotypic Array 96-well plate per the manufacturer’s instructions. Substrate-dependant dye reduction was measured at 690 nm. Increased absorbance reflecting increased metabolism of psicose and methyl pyruvate was consistently observed in Vx isolates and considered to be a vaccine phenotype. Low utilization of these substrates was indicative of WT clinical isolates, including the WT progenitor of the vaccine strain. 14
Media and Culture Conditions
Bacteria were cultured overnight at 37°C and 5% CO2 in Todd Hewitt Broth with 0.5% yeast extract. Prior to inoculation into the dorsal muscles or peritoneal (celomic) cavity of test subjects, bacterial strains were collected by centrifugation for 5 minutes at 13 200 revolutions per minute, washed once, and resuspended to approximately equal concentrations in 0.01M phosphate buffered saline (PBS). Concentrations of inoculated bacteria were confirmed using serial 10-fold dilutions in sterile 0.01M PBS, followed by plating 100 μl on trypticase soy agar with 5% sheep blood (R01202, Remel, Lenexa, Kansas). Colony-forming units were manually counted following overnight incubation at 37°C and 5% CO2.
Zebrafish
Equal numbers of male and female, approximately 4-month old zebrafish were obtained commercially (EkkWill Scientific, Ruskin, Florida) and maintained live in a quarantine tank system at the University of Illinois zebrafish facility under standard conditions. 26 Fish were allowed to acclimate for 2 weeks. Prior to infection, a sample of 15 fish from the quarantine tanks were cultured for the presence of β-hemolytic streptococci, all of which tested negative. Procedures involving animals conformed to the guidelines of the Institutional Animal Care and Use Committee (University of Illinois protocol No. 05196) in compliance with state and federal laws and the standards outlined in the Guide for the Care and Use of Laboratory Animals. 11
Inoculation
Groups of 8 zebrafish were injected IM or IP with 20 μl of 104 to 106 colony-forming units (cfu) per microliter of organisms. Groups were injected with WT, Vx, or S. zoo. Experiments using the 20 μl of 105- and 106-cfu/μl dose and IM route of infection were performed in duplicate. A 0.5-cc Monoject tuberculin syringe (Kendall Healthcare, Mansfield, Massachusetts) with a 0.5-in 28-gauge needle was used to inject 20 μl of bacterial suspension. Fish were anesthetized by immersion in 168-μg/ml Tris-buffered tricaine (A5040, Sigma, St Louis, Missouri) prior to injection. For IM injection, anesthetized fish were held positioned prostrate and injected at a 45° angle relative to the spine immediately cranial and lateral to the dorsal fin. For IP injection, anesthetized fish were placed supine, and the needle was held parallel to the fish’s spine and inserted cephalad into the midline of the abdomen just caudal to the pectoral fins. For the 11 laboratory S. equi strains, the bacteria were cultured, and 20 μl of a 105 cfu/μl suspension in PBS were injected into the dorsal musculature of groups of 8 zebrafish (as described above).
Maintenance and Incubation of Zebrafish Postinoculation
Following infection, zebrafish were housed in sterile 1-liter beakers (4 fish per beaker) in 900 ml of sterile deionized water supplemented with aquarium salts (Instant Ocean, Aquarium Systems, Mentor, Ohio) at a concentration of 60 mg/l. Beakers were covered with perforated aluminum foil and housed in a 29°C incubator with a transparent door allowing ambient light. Fish were monitored every 2 hours for signs of distress (eg, circular swimming or loss of neutral buoyancy). Water changes were performed at 48 hours postinfection, with replacement of 75% to 85% of the water with fresh water, supplemented with aquarium salts as above. Euthanasia was performed using an overdose of Tris-buffered tricane (320 μg/ml).
Histopathology
Whole fish were fixed overnight in 10% neutral buffered formalin, subsequently bisected longitudinally, and then held overnight in decalcification solution (DeltaFORM, Delta Medical Products, Aurora, Illinois). Following decalcification, samples were then routinely processed in paraffin, sectioned at 3 μm, and stained with hematoxylin and eosin using standard techniques. A detailed histologic description of microscopic lesions was performed on hematoxylin and eosin–stained slides by a board certified pathologist (L.B.B.). Several parameters were scored on a 0 to 3 scale (0 = none/minimal, 3 = many/severe), including myonecrosis, leukocyte infiltration at the site of injection, circulating monocytes with intracellular cocci observed in the gill capillaries, and bacterial spread into the adjacent tissues by direct extension—specifically,
Myonecrosis: 0 = no or rare necrotic myofibers, 1 = less than 10 necrotic myofibers, 2 = greater than 10 necrotic myofibers at the site of injection and 3 = spread of muscle necrosis distant from the immediate area of injection
Leukocyte infiltration: regardless of cell type, 0 = no observed infiltrates, 1 = a mild infiltrate localized to the site of infection, 2 = a moderate and localized infiltrate, and 3 = a severe and widespread infiltrate
Circulating monocytes/macrophages: containing intracellular bacteria in the gill capillaries, 0 = none observed, 1 = 1–2 observed, 2 = 2–5 observed, and 3 = greater than 5 observed
Spread of bacteria into adjacent tissues: 0 = no bacteria observed in the dorsal musculature, 1 = small colonies of bacteria localized at the injection site, 2 = large colonies of bacteria localized at the site of injection, and 3 = bacteria spread away from the original site of injection into adjacent tissues following fascial planes
Additionally, liver, spleen, kidney, heart, and central nervous system were evaluated for the presence of cocci.
Comparative Genomic Hybridization
S. equi (WT and Vx) and S. zoo cultures were injected IM into groups of 30 or more anesthetized zebrafish (as described above). Fish were observed every 2 hours for up to 96 hours following injection. Humane euthanasia was performed, with loss of neutral buoyancy as an objective endpoint. Necropsy immediately followed euthanasia. Spleen, kidney, and injection site musculature were collected separately for mRNA extraction. Whole tissue mRNA was extracted from pooled spleen and kidney using an RNeasy Plus Mini Kit (Qiagen, Valencia, California) according to the manufacturer’s instructions with recommended modifications for tissue. Muscle tissue was homogenized via bead milling for 30 seconds using 0.5-mm zirconia-silicate beads (Cole-Parmer, Vernon Hills, Illinois) and mRNA prepared with the RNeasy Fibrous Tissue Mini Kit (Qiagen) according to the manufacturer’s recommendations. Due to insufficient mRNA, samples of kidney were not analyzed further. Isolated spleen and muscle mRNA was sent to Roche NimbleGen (Reykjavík, Iceland) for cDNA synthesis and comparative genomic hybridization using commercially available whole zebrafish genome chips. To control for potential variables introduced during fish handling and injection, gene expression differences are reported as fold changes between S. equi–injected (WT and Vx) and S. zoo–injected groups.
Statistical Analysis
Survival times were recorded and survival curves generated. The Kaplan-Meier product limit estimates of survival curves were used to compare strain differences, with the logrank test used for significance testing. Histopathologic scoring data were analyzed using the nonparametric Kruskal-Wallis test. Median survival of fish injected with laboratory strains was compared between Vx-like and WT-like isolates using a Student’s t-test. In all analyses, statistical significance was set at a level of P < .05. P values between P > .05 and P = .10 were considered trends.
Hybridization data were compared between groups using DNASTAR Array Star 2 software (DNASTAR, Madison, Wisconsin). Genes with at least an 8-fold difference between S. zoo– and S. equi–injected fish were investigated with hierarchical cluster analysis using the nearest neighbor-joining algorithm. Clusters were determined by visual inspection of clustered heat maps created in DNASTAR Array Star 2 software. Within each cluster, individual genes were identified and categorized by class of protein, and finally, the function of the predicted protein for each gene was noted. Genes that have not been annotated in the zebrafish genome were identified on the basis of homology with known protein sequences in GenBank. Hypothetical proteins with poor homology (no region with greater than 80% homology) or no homology with known proteins were excluded from further analysis. Genes covered by several microarray features were reported as individual genes.
Results
Zebrafish Model
The IM route of injection resulted in increased morbidity and mortality compared to the IP route (Fig. 1). A dose-dependent decrease in overall survival was noted over the 104- to 106-cfu/μl dose range using the IM route, and a similar trend was noted using the IP route. The IM route coupled with the 105-cfu/μl dose was determined to be optimal, as it provided the greatest differentiation of the survival curves with a portion of each group surviving. Statistical evaluation revealed a strong effect (χ2 = 17.50, df = 3, P = .00056) on the survival of infected fish compared to noninfected, mock-injected controls (Fig. 1). More important, the model successfully detected attenuation in the Vx strain, with Vx-injected fish living significantly longer than WT
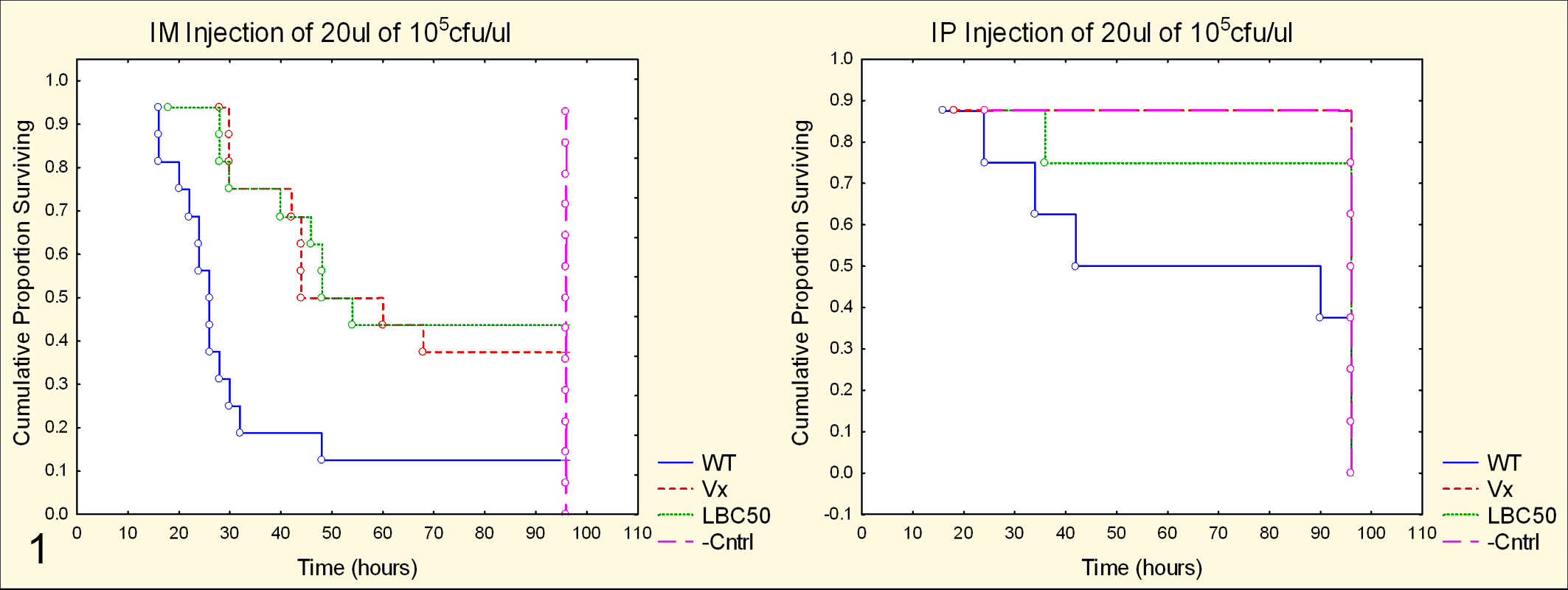
Survival curves of zebrafish injected with Streptococcus equi ssp equi (S. equi), wild type (WT) and modified live vaccine strain (Vx); S. zoo (LBC50); and phosphate buffered saline (Cntrl). At the same dose, zebrafish injected via the intramuscular (IM) route (left) demonstrated decreased survival when compared to zebrafish injected intraperitoneally (IP; right). Note the significantly increased survival of zebrafish injected IM with Vx or LBC50. PBS-injected negative control zebrafish, as well as surviving zebrafish in other groups, were euthanized at 96 hours.
Gross Findings
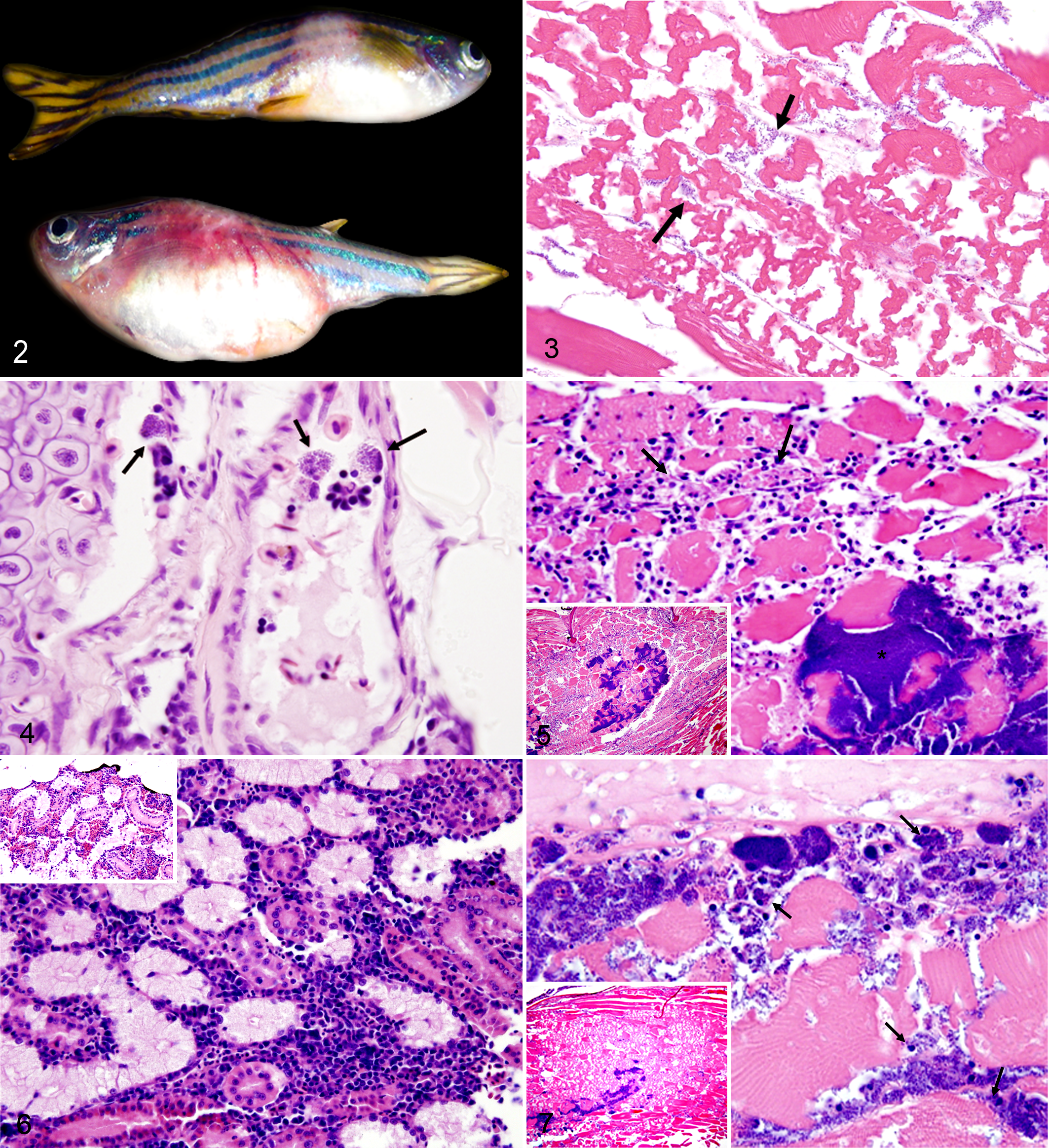
Gross lesions at injection sites varied in appearance among groups. Fish injected with the WT strain rapidly developed evidence of sepsis, characterized by diffuse hyperemia most prominently observed in the nonpigmented areas of the ventrum, tail base, head, and gills. In contrast, most fish injected with the Vx or S. zoo strain developed a focal to locally extensive area of hyperemia, progressing to pallor and resulting in 0.3- to 0.5-cm raised pale tan to white areas (Fig. 2). The tissues adjacent to these pale areas were often hyperemic.
Histology
In fish injected with WT S. equi, rapid mobilization of bacteria was observed via septic spread and direct extension far ahead of the inflammatory response (Fig. 3). Acute myonecrosis was observed and was characterized by fragmented myofibers, which lacked cross striations. The necrosis was generally not accompanied by a prominent cellular infiltrate. Also a significant increase in circulating monocytes with intracytoplasmic cocci was noted in WT-injected fish (Fig. 4).
Fish injected with S. zoo frequently demonstrated an aggressive inflammatory response to the injected bacteria with primarily macrophages surrounding large bacterial colonies at the site of injection (Fig. 5). Circulating monocytes with intracellular bacteria were infrequently observed in the S. zoo–injected fish. In fact, when S. zoo organisms were observed distant to the injection site, they were usually observed extracellularly in large, tight, cohesive colonies. There was also evidence of systemic immune stimulation in S. zoo–infected fish with hypercellularity of the renal interstitium (Fig. 6) and splenic white pulp due to increased numbers of lymphocytes and macrophages. Additionally, the spleen was present more often (χ2 = 6.98, p = .0082) in histologic sections of S. zoo–injected fish (9 of 16) than S. equi–injected (WT and Vx) fish (6 of 32).
In Vx-injected fish, the spread of bacteria via direct extension appeared limited by a mild increase of inflammatory cells (Fig. 7), and a relative decrease in monocyte trafficking compared to WT-injected fish was observed.
In either Vx or WT S. equi–injected fish, rapid infiltration and marked myonecrosis, coupled with increased intracellular survivial and evasion of immune surveillance, appeared important to the disease process. In contrast, S. zoo appeared to be highly immunogenic in the zebrafish with decreased intracellular survival and slow spread through tissues. To evaluate these trends, the nonparametric Kruskal-Wallis test was applied to histologic scores of myonecrosis, leukocytic infiltration, and circulating monocytes. While not significant (P = 0.05), trends were supported: namely, increased myonecrosis in WT-injected fish (H = 4.92, P = .085) and increased numbers of infiltrating leukocytes in S. zoo–injected fish (H = 5.58, P = .0614). There was, however, a significant difference (H = 12.67, P = .0018) in the number of circulating monocytes with intracellular bacteria between WT-injected fish and the Vx- and S. zoo–injected fish.
Comparative Genomic Hybridization
Given the histologic findings, comparative genomic hybridization using whole genome microarrays was utilized to determine the relative gene expression in the muscle (at the injection site) and spleens of zebrafish injected with S. equi (WT or Vx) compared to S. zoo. A threshold limit of an 8-fold difference in expression was set. Based on this threshold, differential gene expression was observed in the muscle and spleen of zebrafish injected with S. equi or S. zoo.
In the muscle of S. equi–injected (WT and Vx) zebrafish, relative increases in expression were observed in genes encoding positive acute-phase proteins, including protein mediators of the complement and coagulation cascades and acute inflammation (Table 1). The gene encoding intelectin-like peptide was dramatically upregulated (82.47-fold increase) compared to S. zoo–infected zebrafish. This peptide shares homology with antimicrobial lectins found in Paneth cells within intestines of mammals and is an important component of innate immunity in teleost fish. 4,13,20,21,23,24 The genes encoding interleukin-11a and major histocompatablity complex class I also demonstrated increased expression in the muscle of S. equi–infected zebrafish relative to S. zoo–infected zebrafish. In contrast, the muscle of zebrafish injected with S. zoo demonstrated increased expression of protein mediators of cellular inflammation, including CXC chemokines, inducible nitric oxide synthase, and cell-signaling peptides (Table 1) relative to S. equi–injected zebrafish. Granulin, a growth factor important for proliferation and differentiation of macrophages in teleost fish, 2,3 was increased approximately 27-fold in the muscle of S. zoo–injected zebrafish relative to S. equi–injected zebrafish.
Relative Gene Expression in the Muscle of Injected Zebrafish. a
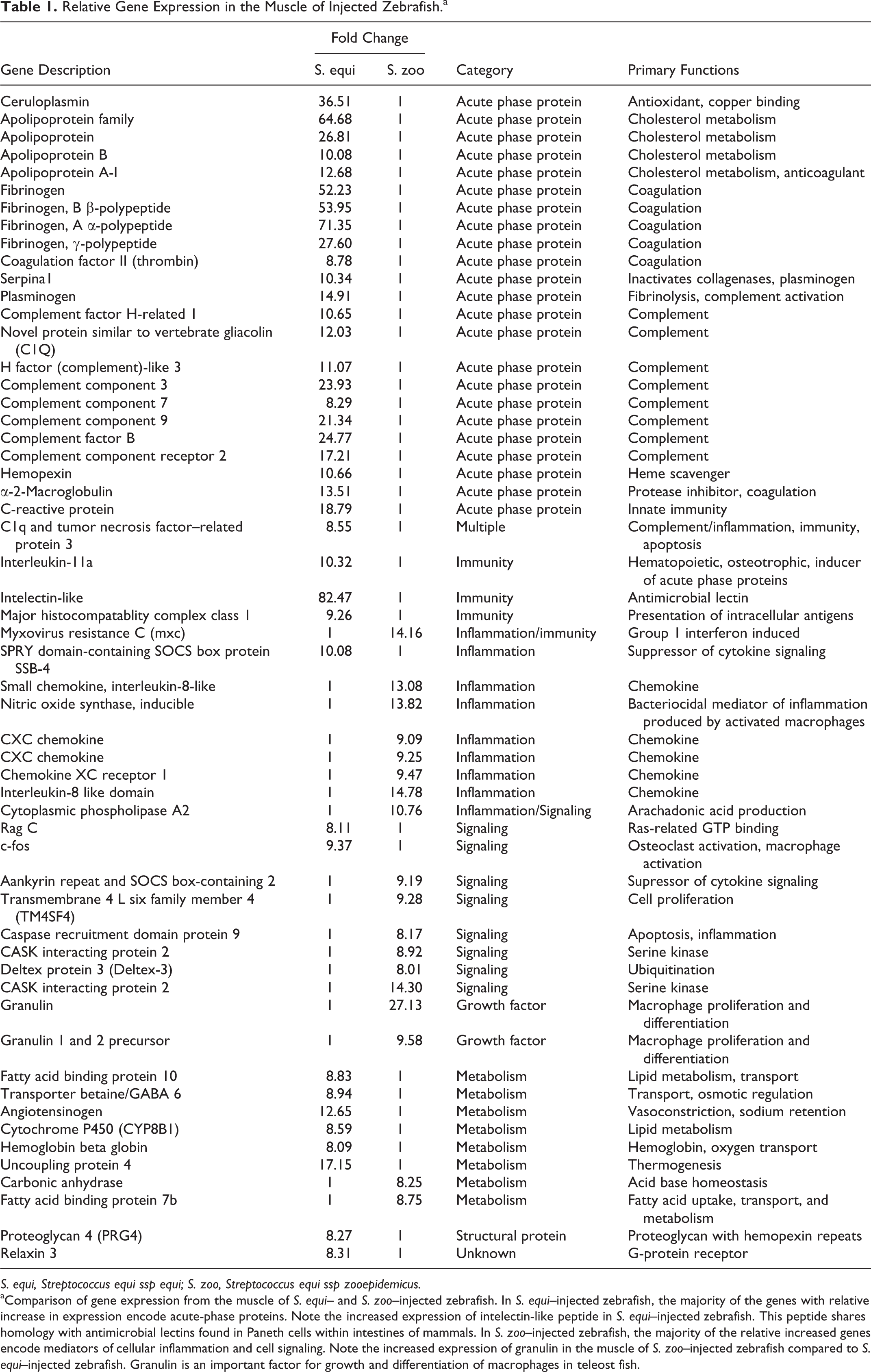
S. equi, Streptococcus equi ssp equi; S. zoo, Streptococcus equi ssp zooepidemicus.
aComparison of gene expression from the muscle of S. equi– and S. zoo–injected zebrafish. In S. equi–injected zebrafish, the majority of the genes with relative increase in expression encode acute-phase proteins. Note the increased expression of intelectin-like peptide in S. equi–injected zebrafish. This peptide shares homology with antimicrobial lectins found in Paneth cells within intestines of mammals. In S. zoo–injected zebrafish, the majority of the relative increased genes encode mediators of cellular inflammation and cell signaling. Note the increased expression of granulin in the muscle of S. zoo–injected zebrafish compared to S. equi–injected zebrafish. Granulin is an important factor for growth and differentiation of macrophages in teleost fish.
In the spleen, differences in gene expression were also observed between S. equi– and S. zoo–injected zebrafish. In S. zoo–injected zebrafish, increased expression of genes encoding intermediate filaments found in epithelium (keratins) as well as pancreatic enzymes and hormones (eg, insulin, glucagon, somatostatin) was observed relative to S. equi–injected zebrafish (Table 2). This finding suggests that pancreatic tissue may have been included with spleen samples from S. zoo–injected zebrafish, which was not the case for S. equi–injected zebrafish. In S. zoo–injected zebrafish, the spleen was pale pink and not engorged with blood, as it often was in zebrafish succumbing to S. equi injection; therefore, it was more difficult to distinguish the spleen from surrounding tissues (eg, pancreatic tissue) in the S. zoo–injected group. The gene encoding major histocompatablity complex class I demonstrated increased expression in the spleen of zebrafish infected with S. zoo, while increased expression of major histocompatability complex class II was observed in the spleen of S. equi–infected zebrafish (Table 2). Other genes with relative increased expression in the spleen of S. equi–infected zebrafish included hemoglobins, factor XIII, and several other genes involved in cellular metabolism (Table 2).
Relative Gene Expression in the Spleen of Injected Zebrafish. a
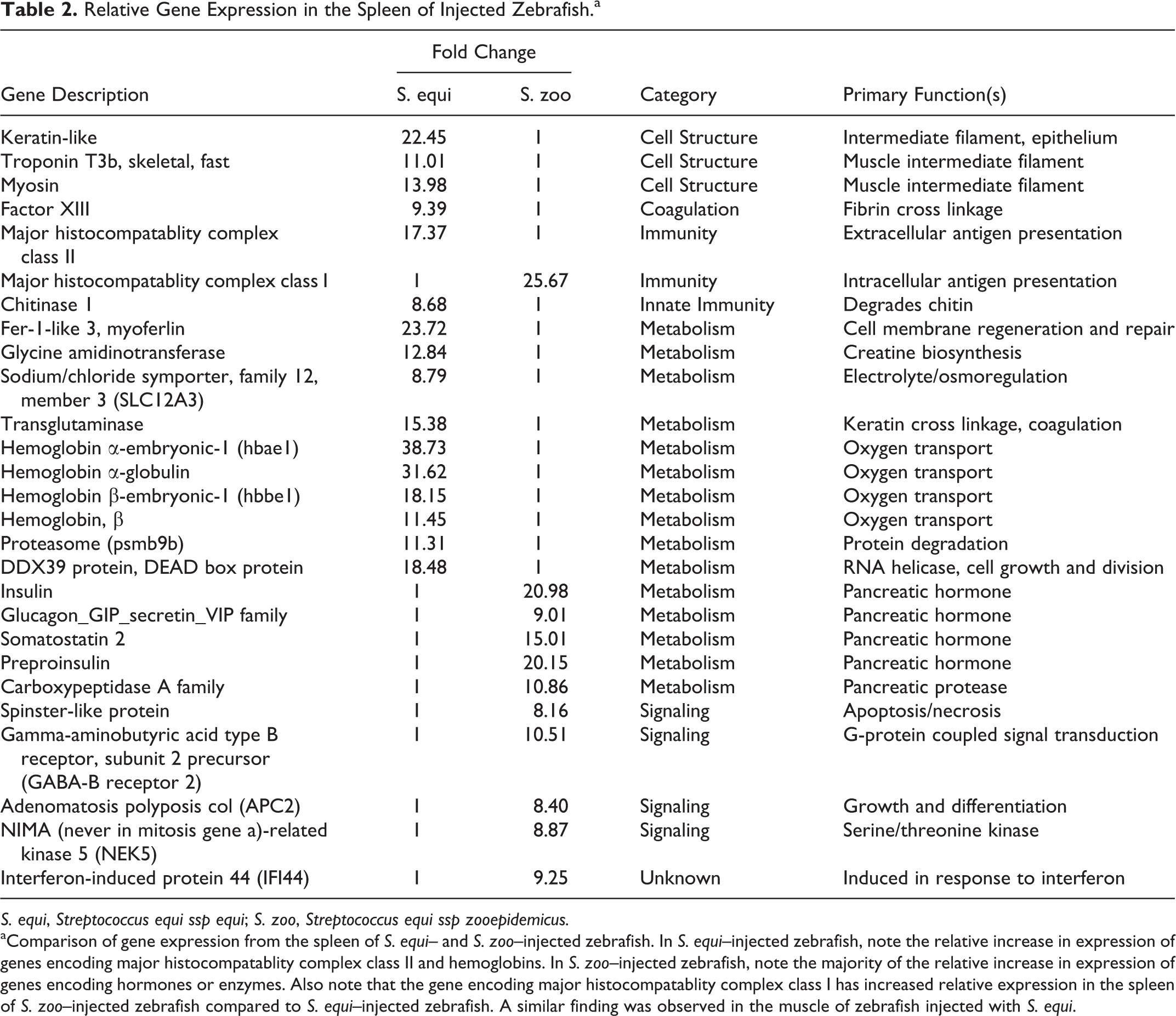
S. equi, Streptococcus equi ssp equi; S. zoo, Streptococcus equi ssp zooepidemicus.
aComparison of gene expression from the spleen of S. equi– and S. zoo–injected zebrafish. In S. equi–injected zebrafish, note the relative increase in expression of genes encoding major histocompatablity complex class II and hemoglobins. In S. zoo–injected zebrafish, note the majority of the relative increase in expression of genes encoding hormones or enzymes. Also note that the gene encoding major histocompatablity complex class I has increased relative expression in the spleen of S. zoo–injected zebrafish compared to S. equi–injected zebrafish. A similar finding was observed in the muscle of zebrafish injected with S. equi.
Comparing Virulence Among Strains
Eleven clinical isolates of S. equi from the University of Illinois Veterinary Diagnostic Laboratory were evaluated genotypically and phenotypically using PFGE, sequence analysis of seM and szp, Biolog Phenotypic MicroArray analysis, and the zebrafish model. Survival curves were created to compare groups of isolates. Results of these analyses are represented in Table 3.
Selected Isolates Tested for Virulence Based on the Zebrafish Model. a

Vx, commercially available modified live vaccine strain of Streptococcus equi ssp equi (S. equi); WT, wild-type S. equi vaccine progenitor strain.
aDetection of attenuation of 11 clinical S. equi isolates. Vx-like isolates (A–E) were identified by high substrate usage in the Biolog profile, the absence of a single guanidine residue in the upstream region of szp, and a single nucleotide polymorphism (G) at the 63rd codon of the seM gene. Isolates H–K were identified as WT-like isolates by low substrate usage in the Biolog profile and WT genotypes in the upstream region of szp.
bRelative usage of psicose, methyl pyruvate; Vx, high substrate metabolism; WT, low substrate metabolism.
cPulsed field gel electrophoresis of Sma I, Not I, and Sfi I digestion patterns; numbers denote WT patterns, while Vx denotes a vaccine-like pattern. Note that the WT progenitor strain is indistinguishable from the Vx strain.
dPresence (+) or absence (–) of G residue in the putative regulatory region at –85.
eVariation in seM sequence at 63rd codon.
fMedian survival time, hr.
As shown in the table, there was a correlation among the upstream mutation in the szp gene, Biolog phenotype, and median survival time of injected zebrafish. Strains B, C, G, and Vx are attenuated, as evidenced by the extended mean survival time. Strain A, while exhibiting a Vx-like metabolic profile, was heavily encapsulated and clearly virulent in the zebrafish model. The PFGE pattern suggested that isolate A has considerable genetic differences from Vx. Isolate G also was interesting, as the Biolog profile predicted virulence; however, the isolate demonstrated attenuation in the zebrafish model. This isolate had a unique adenine deletion 138 base pairs upstream of the szp start codon that may affect the virulence of this strain. Isolate G was not included in subsequent data analysis due to the unique genotype.
The selected isolates were separated into 2 groups for further analysis based on the presence or absence of the upstream szp mutation and the Biolog profile: isolates A–E compose the Vx-like group, and isolates F and H–K compose WT-like group. The median survival times of both groups were compared using a Student’s t-test. A significant increase in median survival time (P = .00563) was observed in fish injected with members of the Vx-like group when compared to the WT-like group.
Discussion
An injection model of Lancefield group C streptococcal pathogenesis using the zebrafish host (D. rerio) has been developed. The model successfully detected attenuation in strains of S. equi when approximately 10 6 cfu were injected into the dorsal musculature. Comparison of the WT progenitor strain to the Vx strain of S. equi yielded a statistically significant increase in median survival time among the Vx-injected group, which was comparable to the median survival time of S. zoo–injected fish. As hypothesized, these results mirror the natural state in horses in which the WT S. equi is more virulent than the commensal S. zoo and Vx. Significantly increased numbers of circulating monocytes with intracellular bacteria were observed histologically in WT S. equi–injected fish, which suggests that intracellular survival and evasion of the respiratory burst may be important in the pathogenesis of this bacterium. Histologic examination of S. equi–injected fish (WT and Vx) revealed little cellular infiltrate into the injection site muscle. In contrast, an intense cellular response was observed at the injection site of S. zoo–injected fish. While statistical analysis applied to a 3-point semiquantitive histologic grading scheme evaluating inflammatory infiltrates failed to attain significance, a strong trend was observed. The histologic findings in fish injected with WT S. equi and fish injected with an isogenic Vx were similar, despite the increased survival of Vx-injected fish. These results suggest that while closely related, S. equi and S. zoo elicit different host responses in the zebrafish.
To assess the host response, comparative genomic hybridization was performed to compare relative gene expression between zebrafish injected with S. equi (WT and Vx) or S. zoo. Differential gene expression was observed in zebrafish injected with S. equi compared to those injected with S. zoo. The most striking differences in gene expression were observed in the muscle at the site of infection. Increased transcription of acute-phase proteins was observed in S. equi–injected zebrafish, including members of the complement and coagulation cascades, as well as mediators of acute inflammation and an antimicrobial lectin. In contrast, genes involved in chemotaxic signaling to phagocytes and growth and differentiation factors for macrophages dominated in injection site muscle in fish infected with S. zoo.
While the gene expression data are consistent with and support the histologic findings, there are possible confounding factors. One important uncontrolled factor is the fact that WT S. equi–injected zebrafish succumbed to septicemia more rapidly than S. zoo–injected zebrafish. As such, it is unclear whether differences observed in gene expression are in fact due to differences in the pathogenic mechanism of the bacteria or simply represent the genes upregulated during acute inflammation (S. equi) versus subacute inflammation (S. zoo). This limitation is ameliorated to some extent due to the inclusion of Vx-infected zebrafish, which had survival times that were statistically indistinguishable from S. zoo–infected zebrafish. However, additional pathogenesis studies are needed to assess the genes expressed during the acute phase of S. zoo infection. Due to the lower-than-expected yield of mRNA, repetition of the expression arrays and confirmation of the findings with targeted real-time PCR data were not possible. However, a high threshold (8-fold change) to detect changes in expression was set to minimize false-positive results in the absence of technical repeats of the microarrays.
Finally, several laboratory strains of S. equi were screened for attenuation using the zebrafish model. These strains fell into 2 categories, based on usage of
Two of these isolates produced unexpected results in zebrafish. Isolate A, despite having a Vx-like Biolog pattern and the single adenine deletion upstream of szp, had a decreased median survival time that was similar to WT. This strain is unique among the Vx-like group in that it is heavily encapsulated. The hyaluronic acid capsule of β-hemolytic streptococci is a well-known virulence factor and is likely responsible for the decreased median survival time in zebrafish injected with this isolate. This isolate also interestingly shares many similarities with the vaccine strain. Both strains have the same Biolog profile and contain the same szp mutation and seM SNP at codon 63; however, the strains have dissimilar PFGE patterns in 2 of the 3 enzyme digests. These findings taken together suggest that this strain may represent a recombinant strain of the Vx. Isolate G also produced unexpected results in zebrafish. This isolate has Biolog and PFGE profiles that suggest virulence, but it was attenuated compared to the other WT strains. The attenuation of isolate G was corroborated in a previous study (in press) by its failure to cause disease when used as a challenge strain in unvaccinated ponies. This isolate has the single adenine deletion in the upstream region of szp. Studies on the effect of this mutation on the expression of szp are in progress. Attenuation of this strain, which was recovered from a clinical case of strangles, may have occurred during serial passage of this isolate in the laboratory. That this isolate is attenuated in zebrafish, as well as in ponies, supports the use of the zebrafish as a translational model for the study of S. equi pathogenesis. Despite the apparent usefulness of the zebrafish as an animal model, an obvious limitation is the inability to create a suppurative lymphadenitis in fish. Fish lack lymph nodes, relying instead primarily on the spleen and tissue-associated lymphoid aggregates. However, the septic form of the disease (bastard strangles) is analogous.
The streptococcal zebrafish model offers many advantages over other animal models for the study of dynamic host–pathogen interactions that influence clinical outcome. Zebrafish are inexpensive, require minimal laboratory space, and are easy to handle and maintain. Furthermore, their small size is particularly useful, as the entire host can be viewed in one longitudinal histologic section. In addition to the practical advantages, the advanced and pliable immune system of zebrafish is highly analogous to the immune system of mammals. 17 Possessing both innate and adaptive immunity, zebrafish have been shown to have analogs of most mammalian immune cells, such as antigen-presenting cells, T cells, B cells, phagocytic cells capable of producing reactive species of oxygen, and circulating granulocytes, including heterophils, similar to neutrophils, which are recruited to sites of injury and are involved in the inflammatory response. 17 Immunoglobulins, cytokines, and complement and major histocompatability complex molecules have been identified in zebrafish. 9 These factors support the continued development of the zebrafish as an accepted model of mammalian diseases.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.