
Abstract
West Nile virus (WNV) infection was diagnosed in 38 psittacine birds based on histology, immunohistochemistry, and reverse transcriptase polymerase chain reaction (RT-PCR). Rosellas (Platycercus spp, n = 13), conures (Enicognathus, Aratinga, and Nandayus spp, n = 6), and lorikeets (Trichoglossus spp, n = 6) represented the most commonly affected species. Clinical signs ranged from lethargy, ruffled feathers, anorexia, and weight loss in most birds to sudden death in others. Except for mild to moderate enlargement of liver and spleen, there were no significant gross lesions at necropsy. Histopathologic findings included lymphoplasmacytic and histiocytic hepatitis, interstitial nephritis, myocarditis, splenitis, enteritis, pancreatitis, and occasionally, encephalitis. Viral antigen was detected by immunohistochemistry in 34 of 35 hearts (97.1%), 29 of 32 pancreata (90.6%), 33 of 37 kidneys (89.2%), 31 of 35 intestines (88.6%), 27 of 33 gizzards (81.8%), 8 of 10 ovaries (80%), 27 of 34 spleens (79.4%), 30 of 38 livers (78.9%), 23 of 32 lungs (71.9%), 21 of 31 proventriculi (67.7%), 14 of 21 adrenals (66.7%), 10 of 16 testes (62.5%), 17 of 30 brains (56.7%), 15 of 27 skins (55.5%), 3 of 6 oviducts (50%), 15 of 34 skeletal muscles (44.1%), 11 of 27 crop or esophagus (40.7%), and 1 of 6 thymuses (16.7%). Kidney was positive for WNV by RT-PCR in all the cases tested. In conclusion, Psittaciformes are susceptible to West Nile virus infection, and WNV infections are often associated with nonspecific clinical signs and widespread viral distribution in this order of birds.
Keywords
West Nile virus (WNV) is an arthropod-borne virus (arbovirus) in the genus Flavivirus and family Flaviviridae that first arrived in the eastern United States in 1999, 23 then spread westward to California in 2003. 30 The virus causes death in a variety of bird species, meningoencephalitis in human and horses, and clinical disease and mortality in other mammalian and reptilian species.5,25,27 WNV is primarily transmitted by mosquitoes, 25 but avian hosts have been incriminated as the vertebrate reservoir. 4 Among avian species, birds such as crows, ravens, jays, magpies, owls, and some raptors are highly susceptible to WNV,10,15,18,26,36 with American crows (Corvus brachyrhynchis) and other corvid species serving as amplifying hosts. 20 The virus has been identified in more than 326 bird species with marked differences demonstrated in clinical and pathologic manifestations, with the disease ranging from subclinical to peracute and lethal and with the pathologic changes varying greatly in their anatomical location, intensity, and chronicity.15,16,21,31–33,36,37
Surveillance reports indicate an increasing number of species affected within the order Psittaciformes, including budgerigars (Melopsittacus undulatus), cockatiels (Nymphicus hollandicus), cockatoos (Cacatua spp), macaws (Ara spp), parrots (Amazona, Rhynchopsitta, Poichephalus, and Psittacus spp), Pacific parrotlets (Forpus coelestis), canary-winged parakeet (Brotogeris versicolurus), rosellas (Platycercus spp), lories and lorikeets (Eos, Lorius, Pseudeos, and Trichoglossus spp), and blue-crowned conure (Thectocercus acuticaudata).7,9,14,24,34 Even though WNV infection in birds has occurred in the Unites States since 1999, not much is known about the infection in psittacines, and the tissue distribution of viral antigen has not been described. Here we report the clinical signs, pathology, and use of reverse transcriptase polymerase chain reaction (RT-PCR) and immunohistochemistry (IHC) for the diagnosis of WNV in psittacines that were submitted to the California Animal Health and Food Safety Laboratory System (CAHFS) in 2004 and 2005.
Materials and Methods
Case Histories
All the psittacines (N = 38) were routine submissions to the CAHFS laboratories at the University of California, Davis. The diagnostic protocol for most birds consisted of detailed clinical history, necropsy, histopathology, and IHC for WNV antigen detection. RT-PCR for WNV and virus isolation in embryonated chicken eggs were also performed for WNV confirmation in a few cases. Epidemiologic information was also collected, such as the location of the aviary (indoor or outdoor), the number and species of birds in the aviary, the presence of mosquitoes in and around the aviary, and the death of crows and other free-flying birds in the vicinity.
Postmortem Examination and Collection of Tissue Specimens
Complete necropsies were performed in 37 of 38 birds. An additional bird was included in the study in which we received only formalin-fixed tissues. Tissue samples collected during necropsy were fixed in 10% neutral buffered formalin, routinely processed, paraffin embedded, sectioned at 5 μm thickness, and stained with hematoxylin and eosin for histopathologic examination.
Immunohistochemistry
Immunohistochemistry was used to identify birds infected with WNV and to determine the tissue and cellular tropism of the virus. All cases (38 of 38) were analyzed by IHC using an immunoperoxidase method (Envision System, DAKO Corporation, Carpinteria, CA). Four-μm-thick paraffin sections were mounted on ProbeOn slides (Fisher Scientific, Pittsburgh, PA) and allowed to dry overnight at room temperature before being baked at 60°C for 30 minutes. After deparaffinization and hydratation, the slides were rinsed in phosphate buffered solution and then treated with proteinase K solution, followed by peroxidase blocking in 10% hydrogen peroxide and application of 3% normal goat serum as a blocking agent. The tissue sections were incubated for 1 hour with the following primary antibodies at a concentration of 1:2000: a rabbit polyclonal anti-WNV antibody (BioReliance, Rockville, MD) and a rabbit monoclonal anti-WNV antibody (ATCC, Manassas, VA) for cases within 2004 and 2005, respectively. Following incubation with peroxidase-conjugated secondary antibody (Envision System reagent) at room temperature for 30 minutes, color was developed with Nova Red Chromogen (ImmPACT NovaRED Peroxidase Substrate, Vector Lab, Burlingame, CA), after which the sections were counterstained with Mayer’s hematoxylin. The amount of antigen expression in different organs, in terms of the number of stained cells and the intensity of staining, was graded subjectively as negative, minimal, moderate, and abundant on the basis of the estimated number of positive cells. Negative controls were run either for each batch of slides or for individual cases.
Biotechnology
Kidneys from 21 birds were tested for WNV by RT-PCR according the method described.8,19
Virology and Electron Microscopy
Homogenized suspensions of tissue pool (heart, kidney, and liver) from 2 cases (Nos. 4, 6) were inoculated into the allantoic cavity of 10-day-old specific-pathogen-free embryonated chicken eggs and tested for WNV according to the method previously described. 8 Negative staining of allantoic fluid samples was examined by transmission electron microscopy. 12
Results
Clinical Signs
A total of 38 psittacine birds submitted between 2004 and 2005 were identified with WNV infection primarily based on IHC (Table 1): 13 rosellas, 6 conures, 6 lorikeets, 2 cockatoos, 2 caiques, 3 scarlet-chested parakeets, 1 Indian ringneck parakeet, 1 derbyan parakeet, 1 red rump parakeet, 1 Princess of Wales parakeet, 1 turquoise parrot, and 1 Australian king parrot. Males (20 of 38) and females (16 of 38) were affected, and most birds were mature (1 to 15 years) except for five 2-month-olds (Nos. 20, 27, 33–35) and one 4-month-old (No. 19). WNV infections were identified in psittacine birds from southern (15 of 38), northern (14 of 38), and central (9 of 38) California. Most outbreaks occurred in outdoor aviaries (23 of 38), and a majority of the owners reported large numbers of mosquitoes and dead crows in and around the vicinity during the time of the outbreaks. The most common clinical signs reported were loss of weight (15 of 38), anorexia (7 of 38), lethargy (5 of 38), listlessness (7 of 38), depression (4 of 38), and weakness (2 of 38). Sudden death without previous clinical signs was reported in 11 cases (Nos. 18–24, 29–31, 37). The clinical presentation in 14 of 38 birds was neurologic with varying degrees of severity. Specific neurologic symptoms consisted of rolling over (2 of 38), legs stretched backward (2 of 38), stumbling (1 of 38), disorientation (1 of 38), and flapping around (1 of 38).
Immunohistochemical Analysis of West Nile Virus Antigen Distribution in Psittacine Birds a
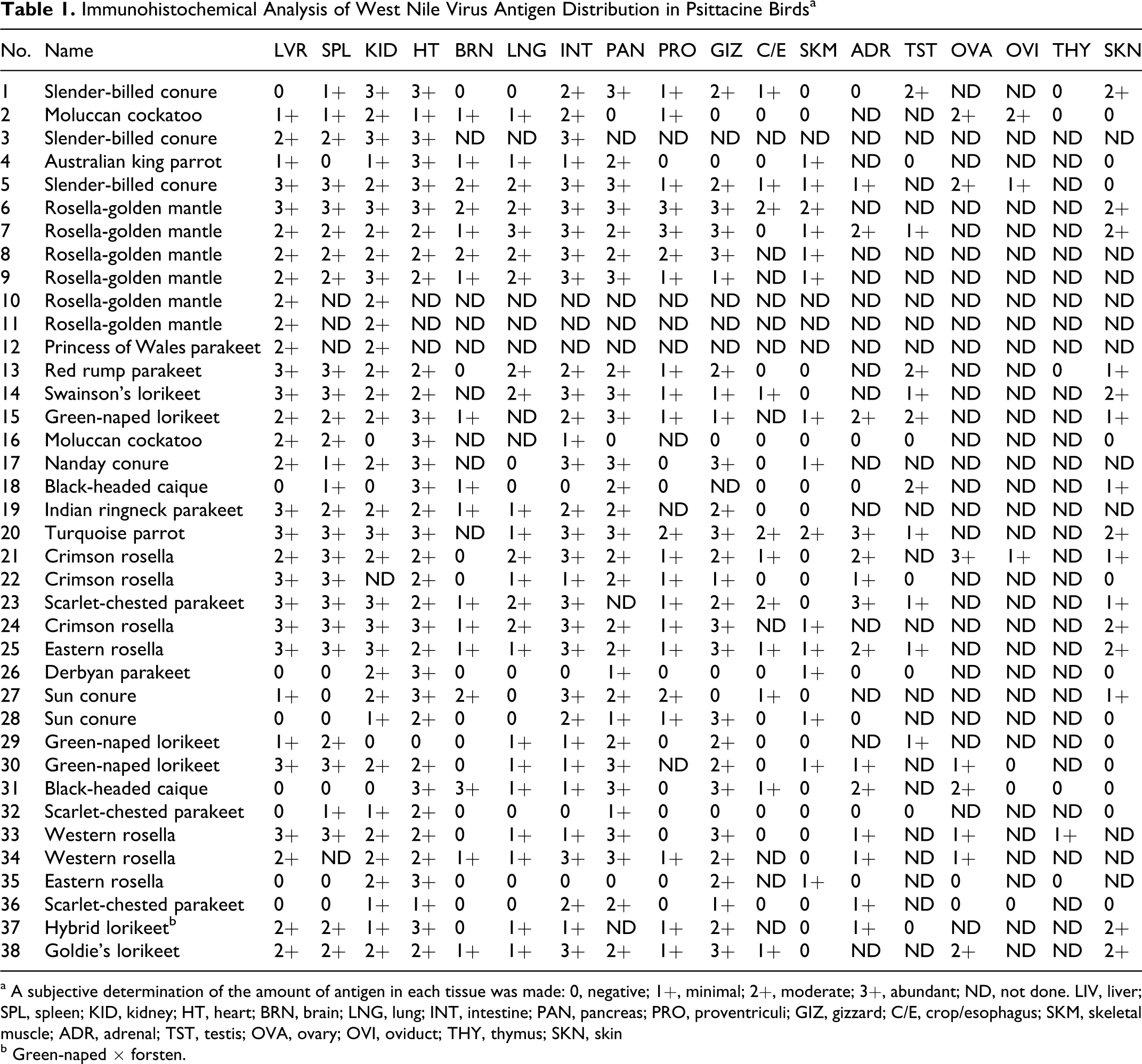
a A subjective determination of the amount of antigen in each tissue was made: 0, negative; 1+, minimal; 2+, moderate; 3+, abundant; ND, not done. LIV, liver; SPL, spleen; KID, kidney; HT, heart; BRN, brain; LNG, lung; INT, intestine; PAN, pancreas; PRO, proventriculi; GIZ, gizzard; C/E, crop/esophagus; SKM, skeletal muscle; ADR, adrenal; TST, testis; OVA, ovary; OVI, oviduct; THY, thymus; SKN, skin
b Green-naped × forsten.
Gross Findings
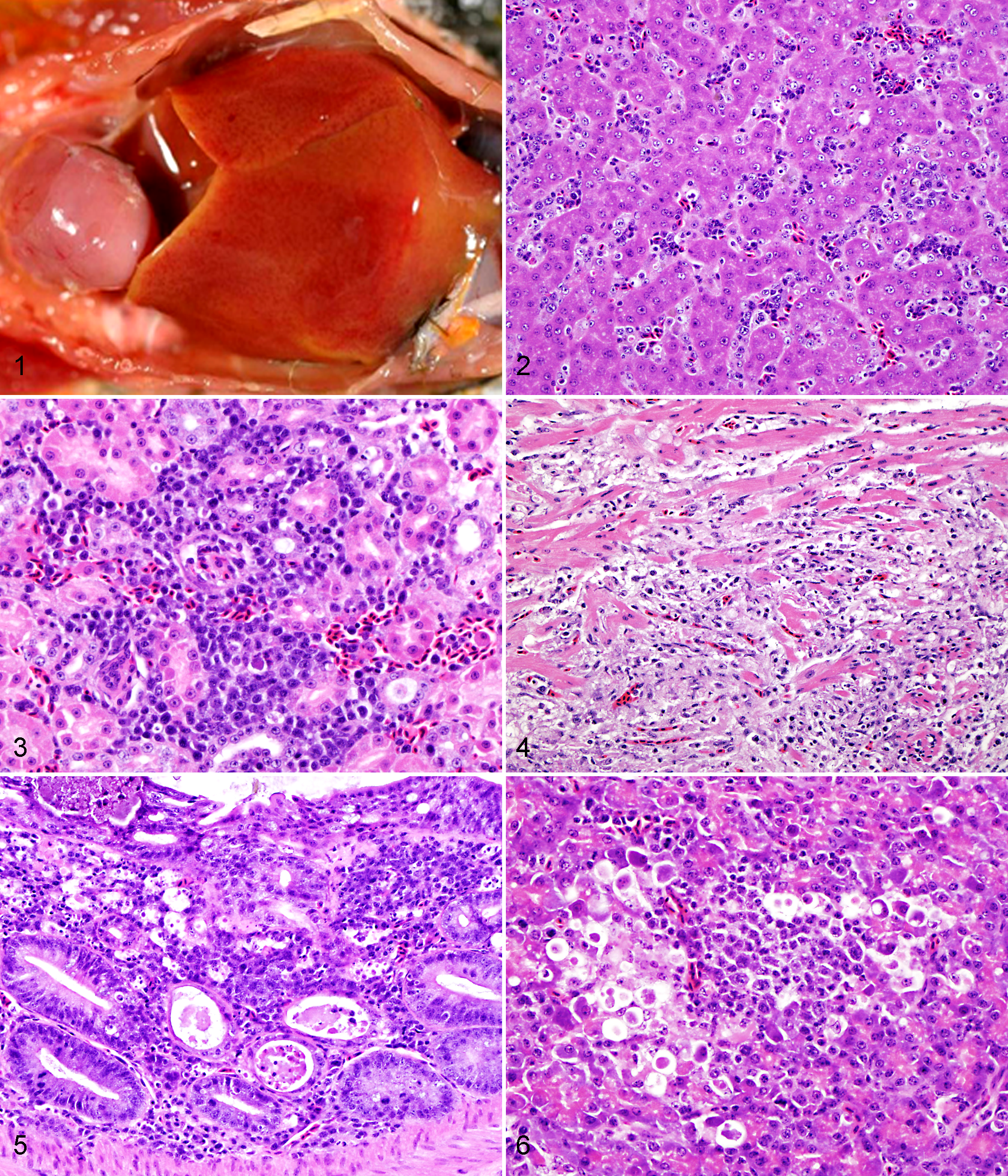
The body condition of most birds ranged from slightly to moderately thin (16 of 38). The most common gross findings were mild to moderate enlargement of the liver (25 of 38) (Fig. 1) and spleen (23 of 38). In some birds, the liver was mottled pale (3 of 38) or had multifocal petechiae (1 of 38). Gross lesions were present in the kidneys of 6 birds (Nos. 1, 2, 4, 22, 23, 35), with diffuse pallor and moderate enlargement. In 3 cases (Nos. 5, 16, 26), heart lesions consisted of large locally extensive myocardial pallor (Fig. 1) with small occasional petechiae on the epicardial surface. Multifocal petechiae on the gizzard serosa were observed in 1 case (No. 6). In some birds (6 of 38), diffuse lung hyperemia (4 of 38), pulmonary edema (2 of 38), and marked congestion of the intestinal tract (2 of 38) were observed. Gross lesions were absent in a green-naped rainbow lorikeet (No. 15).
Histologic Findings
Histologic lesions were most commonly observed in liver, kidney, heart, intestine, and spleen. Hepatic lesions occurred in 21 birds. Moderate to severe, periportal to random chronic lymphoplasmacytic and histiocytic hepatitis was the most common finding (Fig. 2), with multifocal acute coagulative necrosis of hepatocytes in 9 out of 38 cases (Nos. 6, 12, 13, 15, 20–24). The kidneys were effaced by a mild to moderate chronic lymphoplasmacytic interstitial nephritis (20 of 38) (Fig. 3), with rare tubular necrosis (3 of 38) and little or no involvement of glomeruli. Myocardial lesions were present in 17 of 38 birds, with variable degree of severity and involvement of epicardium, myocardium, and endocardium. Lymphocytes, plasma cells, and histiocytes were the predominant cell types of the inflammatory infiltrates (Fig. 4). Multifocal to coalescing myocardial necrosis and multifocal acute hemorrhages were observed in 12 cases (Nos. 1, 4, 15–18, 24–26, 28, 31, 35) and 1 case (No. 20), respectively. Several birds (14 of 38; Nos. 1, 3, 5–7, 26, 28, 30–36) had moderate to severe infiltration of lymphocytes, plasma cells, and histiocytes into the mucosa, lamina propria, and submucosa of the intestinal tract, with variable serosal involvement and crypt necrosis (Fig. 5). Same inflammatory infiltrate was present in the proventriculus of a scarlet-chested parakeet (No. 32) and in the gizzard of a green-naped rainbow lorikeet (No. 30) and a black-headed caique (No. 31). Moderate to severe lymphoplasmacytic and histiocytic splenitis was present in 13 birds (1, 6, 8, 13–15, 19, 24–26, 32, 33, 37), rarely associated with multifocal lymphoid necrosis (5 of 38). A nonspecific reactive lymphoid hyperplasia was observed in 4 cases (Nos. 3, 7, 17, 18). Eleven birds (Nos. 1, 4, 5, 7, 15, 17, 22, 25–27, 35) had moderate to severe pancreatitis characterized by a lymphoplasmacytic and histiocytic inflammatory infiltrate and multifocal necrosis (Fig. 6). Brain lesions were observed in only 7 birds (Nos. 2, 18, 20, 31, 33, 35, 36), mainly in the cerebrum and brainstem and occasionally in the cerebellum. Multifocal perivascular cuffing was primarily composed of lymphocytes and histiocytes and ranged from 2 to 4 cell layers in thickness. Meninges were either minimally or not involved (No. 35). Additional histologic findings included multifocal histiocytic and lymphoplasmacytic steatitis (3 of 38; Nos. 24, 26, 30), lymphocytic (1 of 38; No. 22) or necrotizing (1 of 38; No. 25) adrenalitis, multifocal lymphocytic dermatitis (1 of 38; No. 32), and lymphoplasmacytic, histiocytic, and necrotizing myositis (1 of 38; No. 35). In some birds, the following histologic findings were observed: aspergillosis with variable involvement of air sacs, lung, trachea, sinus, and turbinates (3 of 38; Nos. 4, 21, 25); bursal lymphoid depletion due to psittacine beak and feather disease virus (1 of 38; No. 19); candidiasis in crop and nasal cavities (2 of 38; 1, 13); and Pacheco disease (1 of 38; No. 2).
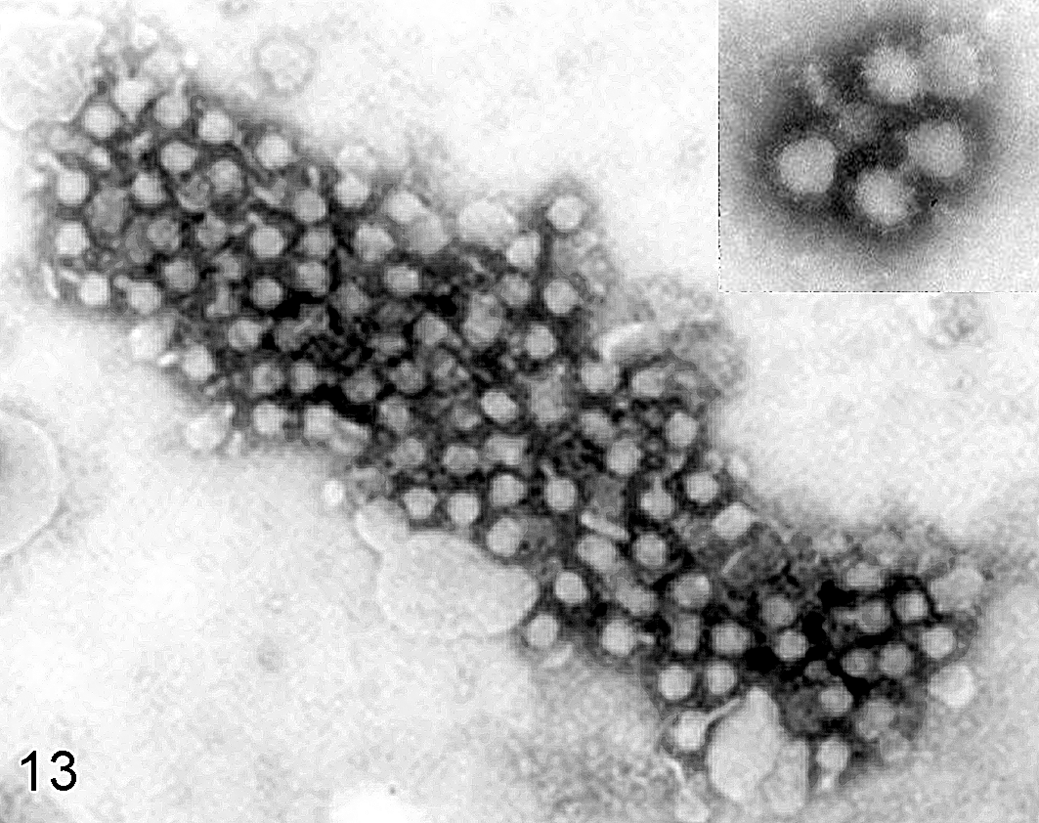
Negative-staining electron micrograph of allantoic fluid from 10-day-old embryonated chicken egg infected with West Nile virus; case No. 6. Multiple spherical virions with approximately 45 nm of diameter. Inset: higher magnification of viral particles.
Immunohistochemistry
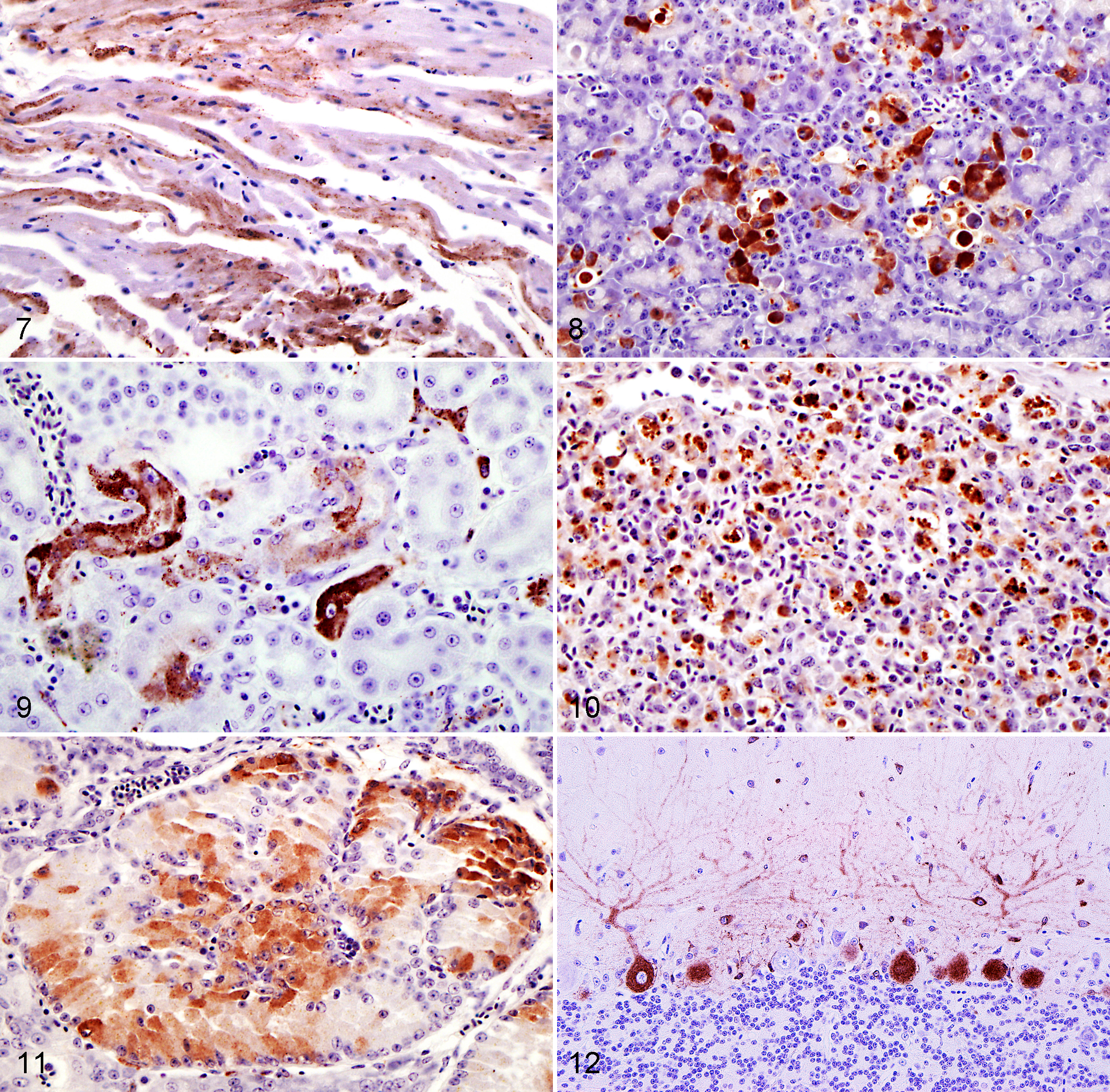
IHC revealed intracytoplasmic staining of WNV viral antigen in many tissues (Table 1). Whereas monoclonal antibody used for testing cases of 2005 is specific for WNV antigen, polyclonal antibody can detect other flaviviruses (including St Louis encephalitis virus and tick-borne encephalitis group of flaviviruses), 32 but the application of RT-PCR confirmed the suspicion of WNV infection in these cases. WNV antigen was consistently detected in the heart (97.1%), pancreas (90.6%), kidney (89.2%), intestine (88.6%), gizzard (81.8%), ovary (80.0%), spleen (79.4%), and liver (78.9%). The association between viral antigen and histologic lesions occurred in 70.0% of liver, 60.6% of kidney, 50.0% of heart, 48.1% of spleen, 45.2% of intestine, 41.2% of brain, 37.9% of pancreas, 14.3% of adrenal, 7.4% of gizzard, 6.7% of skin and skeletal muscle, 4.7% of proventriculus. In the crop or esophagus, testes, ovary, oviduct, thymus, lung, WNV antigen was not associated with histologic changes. 32 Tissue macrophages and blood monocytes appeared to be the target of infection, as demonstrated by the strong immunolabeling of splenic macrophages, Kupffer cells, blood monocytes, and macrophages in interstitial connective tissues and inflammatory foci. Viral antigen was commonly detected in the cytoplasm of endothelial cells and in rare smooth muscle cells of large and small blood vessels. WNV antigen was abundant in the heart of most birds (34 of 35, 97.1%), labeling myocardial fibers, infiltrating macrophages, and interstitial fibroblasts (Fig. 7). Staining was especially common within macrophages infiltrating the epicardial adipose tissue. Most cases of histologically detectable lymphoplasmacytic and histiocytic myocarditis (11 of 17), as well as myocardial necrosis (10 of 12), were associated with abundant antigen within inflammatory and parenchymal cells. Flattened endothelial cells of the endocardium contained antigen in a few cases (Nos. 24, 25, 38), whereas rare Purkinje cells were strongly labeled in only 1 bird (No. 27). Antigen was also detected in large vessels of the heart in a few birds, within endothelial cells and in rare smooth muscle cells (Nos. 6, 7, 21, 23, 33, 37). Within the pancreas (29 of 32), viral antigen was observed mainly in the exocrine acinar cells (Fig. 8) and in a few pancreatic islets in 3 of 29 cases (Nos. 21, 24, 30). Moderate (5 of 11) to abundant (3 of 11) amount of antigen was associated with inflammatory and degenerative processes. In the kidney, antigen was present in interstitial cells and infiltrating macrophages, rarely within tubular epithelium and multifocally in the glomeruli (Fig. 9). In most birds, WNV antigen in the kidney appeared to predominate in glomeruli and interstitial cells. In the intestine, crypt epithelial cells, enterocytes, macrophages, and interstitial cells of the mucosa, lamina propria, submucosa, and muscular layer contained antigen, showing strongest immunoreactivity in 35.7% of enteritis. Variable labeling of smooth muscle cells was observed, whereas the myenteric plexus was constantly negative. In proventriculus, viral antigen was present in macrophages of the interglandular and submucosal lamina propria, in rare glandular epithelial cells of the compound tubular alveolar glands, and in columnar epithelium of luminal surface. Viral antigen was also common in the gizzard, within columnar epithelial cells of mucous membrane, epithelial cells of glandular tubules, smooth muscle cells, and interstitial cells of lamina propria and muscular wall. Multifocal squamous epithelial cells and rare smooth muscle cells of the wall of the crop and esophagus were positive. In the liver, antigen was observed in virtually all Kupffer cells, whereas positive staining of hepatocytes was uncommon (6 of 30; Nos. 20, 22, 24, 25, 30, 33) and bile duct epithelial cells were negative. Out of 21 cases of hepatic histologic changes, the amount of viral antigen was high (n = 11) to moderate (n = 6). In most cases (66.7%), areas of hepatic necrosis were characterized by strong and diffuse immunoreactivity. Antigen-positive monocytes of macrophages were detectable in the sinusoids of the red pulp of the spleen and widespread throughout the splenic parenchyma (Fig. 10). Rare endothelial cells and smooth muscle cells of central arteries contained viral antigen. Positive splenic lymphocytes were not evident. The degree of inflammation, necrosis, and hyperplasia was associated with variable amount of viral antigen. Among endocrine organs, WNV antigen was most commonly detected in the adrenals (14 of 21, 66.7%), as localized in cortical interrenal cells, interstitial fibroblasts, and infiltrating macrophages (Fig. 11). Parenchymal cells of thyroids and parathyroids were negative, whereas rare positive interstitial cells and macrophages were observed. In the ovary, follicular and thecal cells of secondary follicles and large follicles undergoing cystic atresia were strongly labeled. Positive follicular cells were more numerous in atretic than secondary follicles. Stromal cells variably contained antigen whereas oocytes were negative. In 10 of 16 cases (Nos. 1, 7, 13, 14, 15, 18, 20, 23, 25, 29), testes were positive, not only in infiltrating macrophages and resident interstitial cells, but also in germ cells within seminiferous tubules (Nos. 23, 25, 29). Brain was positive in 17 of 30 cases. Viral antigen was detected in neurons and glial cells of cerebrum and brain stem, capillary-associated cells (astrocytes, microglia or extravasating macrophages), Purkinje cell bodies, and dendrites of cerebellum. Occasionally, basket cells, granule cells, and glial cells of the molecular cerebellar layer contained antigen (Fig. 12). Rare labeling was detected in the meninges (Nos. 22, 23, 25, 31), mainly in infiltrating macrophages and endothelial cell of blood vessels. In 2 of 11 cases (Nos. 23, 38), rare ganglion cells in the peripheral nervous system contained viral antigen. Spinal cord and peripheral nerve were negative. Immunohistochemistry identified variable expression of WNV antigen in nasal cavity (4 of 7), syrinx (2 of 4), trachea (4 of 10), and lungs (23 of 32). Within the upper and lower respiratory tract, besides infiltrating macrophages and resident fibroblasts, rare epithelial cells of the upper respiratory tract and lining the air capillaries in the lung contained viral antigen. Within thymus (1 of 6; No. 33) and bursa (4 of 6; Nos. 19, 20, 24, 33), the labeling was restricted to interstitial cells and rare medullary reticular cells and macrophages of thymic parenchyma and bursal lymphoid follicles. Hematopoietic cells of bone marrow were positive in 5 of 8 cases (Nos. 14, 19, 22, 24, 38). Viral antigen was present in 55.5% of the skin sections examined (15 of 27), mostly within fibrocytes and macrophages of the dermis or subcutis. In the sections containing growing feathers, the antigen was also present in interstitial cells and in rare epithelial cells of the feather pulp. Mild to moderate labeling was observed in skeletal muscle fibers (15 of 34). No virus antigen was detected in bone, eye, and ear.
Biotechnology
Kidney was positive for WNV by RT-PCR in all 21 birds tested (Nos. 14–16, 20–26, 28–38).
Virology and Electron Microscopy
Infected chicken embryos died in 3 to 5 days. The embryos were stunted and occasionally hemorrhagic. Negative-staining electron microscopy in samples of the allantoic fluid revealed round to spherical viral particles measuring approximately 40 to 50 nm in diameter in large aggregates and some in small numbers (Fig. 13). Some of the viral particles were surrounded by pale granules, probably representing immunoglobulins (Fig. 13, inset).
Discussion
The immunohistochemistry and RT-PCR results, in combination with the histopathologic changes, indicate that all 38 psittacine species died as a result of WNV infection and that psittacines should be considered susceptible to WNV. Although the most common affected species in this limited study were Australian parakeets such as the rosella, the susceptibility to WNV infection could not be inferred, given that it is affected by different husbandry, contact with mosquitoes, and the number of species submitted to diagnostic service.
In affected birds (as demonstrated by IHC), the virus showed a nonrestricted organ tropism, infecting essentially all major tissue systems and a wide variety of individual cell types, in most cases without microscopic changes. This widespread viral dissemination has been reported in other native and exotic birds.9,21,32,37,38 WNV in psittacines exhibited a prevalent localization in myocardium, cells of the mononuclear phagocyte system (MPS), fibroblasts, smooth muscle cells, endothelial cells, and multiple epithelial cell types. In particular, replication of the virus within MPS cells and dissemination throughout the body could be of primary importance in the wide variety of infected tissues. Macrophages are the most common cells to be stained by IHC in all infected organs, as followed later by parenchymal cells and, last, epithelial cells of some organ systems. 36 Although several elements of the systemic immune system work together during WNV infection to control viral growth and dissemination, the role of macrophages remains controversial. 11 Dissemination of WNV occurs before development of a full adaptive immune response; thus, the innate immune system and, especially, the macrophages are critical for resistance to infection. 2 However, WNV itself is able to suppress human macrophage functions by inhibiting the production of proinflammatory and antiviral cytokines, such as interleukin 1β and type 1 interferon, and by interfering with the Janus kinases / signal transducers and activators of transcription signaling pathway, necessary for macrophage activation. 22 Moreover, infiltrating and infected macrophages likely contribute to underlying immunopathology during WNV encephalitis. 35
As in owls, red-tailed hawks, and Cooper’s hawks,15,37 WNV infection in psittacines appears to be capable of causing a chronic disease, characterized by severe chronic inflammatory lesions associated with poor nutritional state. In contrast, crows die acutely of WNV infection20,21 and frequently lack inflammatory lesions. 38 It is possible that psittacines are somewhat more resistant than crows to infection and that the infection may exist longer in these birds, thereby stimulating a greater inflammatory response before illness and death.
On gross examination, no prominent lesions were observed, except hepatosplenomegaly and loss of weight, considered nonspecific findings. Lack of specific gross findings has been seen in psittacines and in other avian species infected with this virus.6,9,15,21,32 Histologically, several birds had nonspecific lesions in a variety of tissues, but lesions considered to be specifically virus induced were present in the heart, spleen, pancreas, liver, kidney, and intestine with lymphoplasmacytic and histiocytic inflammatory infiltrate associated with necrotizing lesions or not. These organs should be considered the optimal tissues to collect for routine histologic evaluation from psittacines suspected of WNV infection. The main difference observed in psittacines compared to other avian species is the variable involvement of the nervous system, with only 18.4% cases of encephalitis with lymphohistiocytic perivascular cuffing. This histologic finding is consistent with the rare neurologic signs observed and the immunohistochemical distribution of the viral antigen. Only 56.7% of the birds were positive for WNV antigen in the brain, often associated with minimal staining score. Moreover, peripheral nerve and spinal cord were not involved. In most species, the brain seems to be one of the main targets of infection.9,20,21 Carboni et al 6 recently described histologic lesions in brain and spinal cord in a sun conure; however, even highly susceptible birds (eg, crows, magpies, blue jays, owls) could not exhibit an evident neurotropism, with relatively limited viral antigen in the brain and mild or no histologic changes.15,32,38
The evaluation of immunohistochemical score in our cases revealed that intestine, pancreas, heart, kidney, spleen, liver, and gizzard have high levels of antigen and that these tissues should be considered the optimal ones for WNV IHC staining in psittacines. Lower levels were present in the kidney and little viral antigen was evident in the lungs, adrenal, brain, proventriculus, and ovary. The high concentration of antigen in intestinal epithelial cells suggests that the transmission might occur by means other than mosquito bites, such as the orofecal route or direct contact, as proposed in experimentally and naturally infected geese and crows.1,10,21 WNV was detected for a few days in feces of experimentally infected turkeys and chickens, although contact animals did not developed infection.31,33 Furthermore, the findings in WNV infection of alligators and other aquatic animals suggest that contaminated environment, including food and water, may contribute to the spread of WNV. 17 The main problem with psittacines is the minimal viral competence associated with the lack of oral or cloacal shedding and the low level and duration of viremia. 21 Apart from the study by Komar et al 21 —which, because of the extensive scope, included only 12 psittacines (6 budgerigars and 6 monk parakeets) for viremia determination—detailed experimental and field studies are lacking that assess the competency of psittacines for WNV. These results should be consolidated with other data, including higher numbers and species of Psittaciformes, to better estimate the host potential. Moreover, in all avian species, there is a need for additional studies to examine both the viability of WNV in fecal material (and other bodily secretions in the environment) and the potential for zoonotic transmission.
Viral detection in psittacines has been observed in 55.6% of examined skin, with prevalent involvement of the feather pulp. A high detection rate of WNV has been demonstrated in vascular feather pulp from the base of the growing immature feathers of WNV-infected corvid carcasses; 13 feather follicular epithelium and pulp were also positive by IHC in Northern American owls. 16 These results, as obtained in different avian species (including psittacines), suggest that feathers might be a source of transmission, other than a useful tool for the WNV infection diagnosis. Feather testing is already used in psittacines for the surveillance of psittacine beak and feather disease and avian polyomavirus infection.3,29 The only limitation to this test is the variable expression of WNV antigen in the feather pulp, somewhat correlated to the viremia levels. 26 In association with the well-known vector-borne transmission and the suggested but not proven orofecal and direct contact routes, the detection of high amount of antigen within the reproductive tract, especially ovary, raises the question of a possible vertical transmission. 28 A high viral load has been discovered within the ovary of blue jay, American crow, Black-billed magpie, ring-billed gull, heron, cormorant, and owl.16,21,32,36 Even if the oocytes had been negative in all our cases, positive granulosa cells can convey viral antigen into the yolk sac as they slough. The presence of virus in the testes leaves open the possibility of a sexual transmission. WNV antigen has rarely been reported in the testes in birds, usually associated with macrophages and interstitial cells and rarely within seminiferous tubules.16,36 Unlike with IHC, no comment can be made on the best tissue for RT-PCR, because kidney was the only tissue routinely processed for this assay. However, previous studies have demonstrated that viral RNA appears to be present in high concentration or recovered in greater amount from kidney tissue.15,26 Compared to virus isolation and RT-PCR, IHC provides a better understanding of pathogenesis and confirms the sites of virus replication; nonetheless, both tests are approximately equal in their ability to detect WNV. 14
Although it is widely recognized that crows and other corvids are highly susceptible to WNV, many other bird species are susceptible, including psittacines. WNV infection in psittacines poses a diagnostic challenge because clinical signs and lesions are nonspecific, such as hepatosplenomegaly and loss of weight. Nephritis, myocarditis, splenitis, hepatitis, enteritis, and pancreatitis represent the primary lesions, and IHC—especially on intestine, pancreas, heart, kidney, spleen, and liver—can be used to confirm the disease. Even if the epidemiology and route of transmissions of WNV infection in psittacines are not yet understood, the high rate of viral antigen detection emphasizes that necessary precautions must be taken to ensure the safety of those in contact with a suspect bird and to prevent the possible exposure to this zoonotic pathogen.
A retrospective search of the CAHFS database over the last 4 years (2006 to 2009) revealed 31 additional cases of WNV diagnosed in psittacines with similar clinical signs and lesions and with the virus confirmed by RT-PCR on the kidneys. Therefore, the growing prevalence of WNV infection in Psittaciformes should be taken as an important diagnostic consideration by avian practitioners and pathologists alike, especially in birds with nonspecific clinical signs or in birds found dead with no prior symptoms and a lack of pathognomonic lesions. In fact, most of the birds examined in this study were routine submissions from the aviaries, and WNV was not suspected by the owners. To our knowledge, this is the most comprehensive study of WNV infection in psittacines ever undertaken describing the species, age, and sex of birds, as well as clinical signs, virology, electron microscopy, histopathology, and immunohistochemistry.
Footnotes
Acknowledgment
We thank E. J. Hurley, M. Manzer, K. Sverlow, A. Hernandez, R. Nordhausen, H. Moriyama, R. Gonzales, R. Rezvani, D. Ramirez, and L. Alsing for their technical assistance.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.