
Abstract
Molecular regulation of fibrosis in chronic canine hepatitis is poorly understood. The authors employed quantitative polymerase chain reaction (PCR) to determine the expression levels of genes reported to be related to fibrosis in other species (human, mouse, and rat) and to elucidate the relationship of these genes with the degree of fibrosis and the presence or absence of ascites and/or jaundice in dogs with hepatitis. Nine fibrosis-related genes were assayed: PDGFB, PDGFD, MMP2, TIMP1, THBS1, COL1A1, COL3A1, TGFB1, and TGFB2. Liver samples of 15 dogs with chronic hepatitis and 4 healthy control dogs were obtained via laparoscopic biopsy and subjected to histologic and quantitative PCR analyses. The expression of all 9 genes showed significant positive correlation (P < .01, r > .70) with the degree of fibrosis. Furthermore, the expression levels of all genes except TGFB1 were significantly higher (P < .05) in dogs with hepatic failure–related symptoms (ascites/jaundice). Results suggest that these 9 genes are integral to the development of fibrosis in canine chronic hepatitis.
Chronic hepatitis in dogs is well recognized and a documented liver disorder. 9,18,22,24 The disease is characterized by hepatocellular apoptosis or necrosis, a variable mononuclear or mixed inflammatory cell infiltrate, regeneration, and fibrosis. 24 Fibrosis is not only used to determine the stage of hepatitis and prognosis of the disease 24 but may also serve as a therapeutic target. 4
Hepatic fibrosis is the excessive accumulation of extracellular matrix proteins, including collagen I, III, and IV, as well as fibronectin, elastin, laminin, and proteoglycans. 11,12 Collagens I and III are the two chief components of the fibrous tissue in the liver. These proteins are encoded by several genes, but major components are produced by COL1A1 and COL3A1. 11,25 Increased synthesis and decreased degradation of these proteins result in their accumulation. Both processes are mediated by activated by hepatic stellate (Ito) cells, 11,12 which are activated when the liver is involved in an inflammatory process. Stellate cells produce various components of the fibrous tissue, remodel liver architecture by expressing matrix metalloproteinases (eg, matrix metalloproteinase 2 [MMP2]), and inhibit degradation of collagens by expressing tissue inhibitors of matrix metalloproteinases (eg, TIMP1) 5 by transforming growth factor β (TGFB) and platelet-derived growth factor (PDGF). TGFB acts as a potent growth inhibitor for many cell types, and it is involved in the control of parenchymal apoptosis and the stimulation of extracellular matrix component synthesis. 11 The TGFB family contains 3 isoforms—namely, TGFB1, TGFB2, and TGFB3—but studies concerning the function of TGFB in hepatic fibrosis primarily focus on TGFB1. 11 TGFB is secreted as a precursor and functions only upon activation. 11,12
The PDGFs influence the development of liver fibrosis primarily by inducing the expression of thrombospondin 1 (THBS1), a member of the thrombospondin family of multimeric, multidomain glycoproteins that function at cell surfaces and in the extracellular matrix. 1,7 Thrombospondin 1 expression activates TGFB proteins and leads to the development of liver fibrosis. 3,8 The PDGF family is a group of proteins involved in embryonic development, cell proliferation, cell migration, and angiogenesis. They typically form homodimers or heterodimers with the same or other members of the PDGF family 2 to carry out their functions. Two members of the PDGF family, PDGFB and PDGFD, are believed to play important roles in hepatic fibrosis through activation of TGFB and induction of hepatic stellate cell proliferation by binding to the PDGFβ receptor. 2,6
To date, research on the molecular mechanisms of hepatic fibrosis in dogs has been limited to elucidating the functions of the TGFB pathway in disease development and progression. 20,21 Here, we describe the expression of a number of other genes that are reported to be related to hepatic fibrosis in other species (human, mouse, and rat), with the aim of enhancing basic and comprehensive knowledge about the mechanisms of hepatic fibrosis in canine chronic hepatitis.
Materials and Methods
Dogs
Fifteen dogs were included in this study, as admitted to the Veterinary Medical Center at the University of Tokyo or the Nihon University Animal Medical Center and clinically and histologically diagnosed with canine chronic hepatitis/cirrhosis. The ages of the dogs ranged from 2 to 11 years. The breeds included Labrador Retriever (n = 6), Doberman Pinscher (n = 2), Yorkshire Terrier (n = 2), Shiba Inu (n = 1), Miniature Dachshund (n = 1), Chihuahua (n = 1), Irish Setter (n = 1), and American Cocker Spaniel (n = 1). The study group comprised 2 males, 4 females, 2 castrated males, and 7 neutered females. The clinical signs varied among the dogs and included ascites (n = 6), jaundice (n = 5), lethargy (n = 6), and weight loss (n = 3). The concentration of copper in the liver tissue was determined (SRL Inc, Tokyo, Japan) for 4 of 6 Labrador Retrievers and all the Doberman Pinschers; however, none showed the high copper accumulation consistent with copper-related hepatitis reported in these breeds (range, 77.2–616.0 μg/g dry weight).
Four beagles were used as controls (age range, 2 to 4 years). Of these, 3 were neutered females and 1 was an intact male. None of the control dogs exhibited any clinical signs, and the results of serum biochemical screening tests—including alanine aminotransferase activity, alkaline phosphatase activity, urea nitrogen concentration, and serum creatinine concentration—were within reference ranges. All the samples obtained from the beagles were obtained in accordance with the University of Tokyo Institutional Animal Care and Use Committee regulations.
Liver tissue was obtained by laparoscopic biopsy. Two or more samples were immediately immersed into 10% neutral buffered formalin, and 1 sample was placed into 1 ml of RNAlater solution (Ambion, Austin, TX) or liquid nitrogen. Formalin-fixed samples were embedded in paraffin for histologic analysis. The RNAlater-dipped samples were stored at 4°C overnight and then at –80°C until RNA extraction. Fresh-frozen samples were stored at –80°C, treated with RNAlater-ICE (Ambion), and stored until assayed.
RNA Extraction
For RNA isolation, the samples were thawed on ice, cut into 25- to 30-μg pieces, and homogenized using TissueRuptor (Qiagen, Germantown, MD) in 600 μl of lysis buffer of the RNeasy Mini Kit (Qiagen). RNA extraction was performed according to the manufacturer’s protocol. The extracted RNA was then treated with DNase I (Invitrogen, Carlsbad, CA) and reverse-transcribed with a PrimeScript RT reagent kit (Takara Bioscience, Tokyo, Japan) according to the manufacturer’s protocol. RNA integrity was confirmed with gel electrophoresis.
Quantitative Polymerase Chain Reaction Analysis
Nine fibrosis-related genes were selected for quantitative analysis: MMP2, TIMP1, PDGFB, PDGFD, THBS1, TGFB1, TGFB2, COL1A1, and COL3A1. Two genes, RPL13A and RPL32, were included as internal controls.
Gene expression was quantified in duplicate samples using the Thermal Cycler Dice Real-Time System (Takara Bio Inc, Tokyo, Japan) with SYBR premix EX Taq (Takara Bio Inc) or SYBR II premix EX Taq in 96-well plates according to the manufacturer’s instructions. Primers were designed using Primer 3 (http://primer3.sourceforge.net/) or PrimerExpress (Applied Biosystems, Carlsbad, CA,) or were selected by referring to previous publications. 16 The sequences of the primer pairs used are shown in Table 1 .
Primers List for Quantitative Polymerase Chain Reaction Analysis
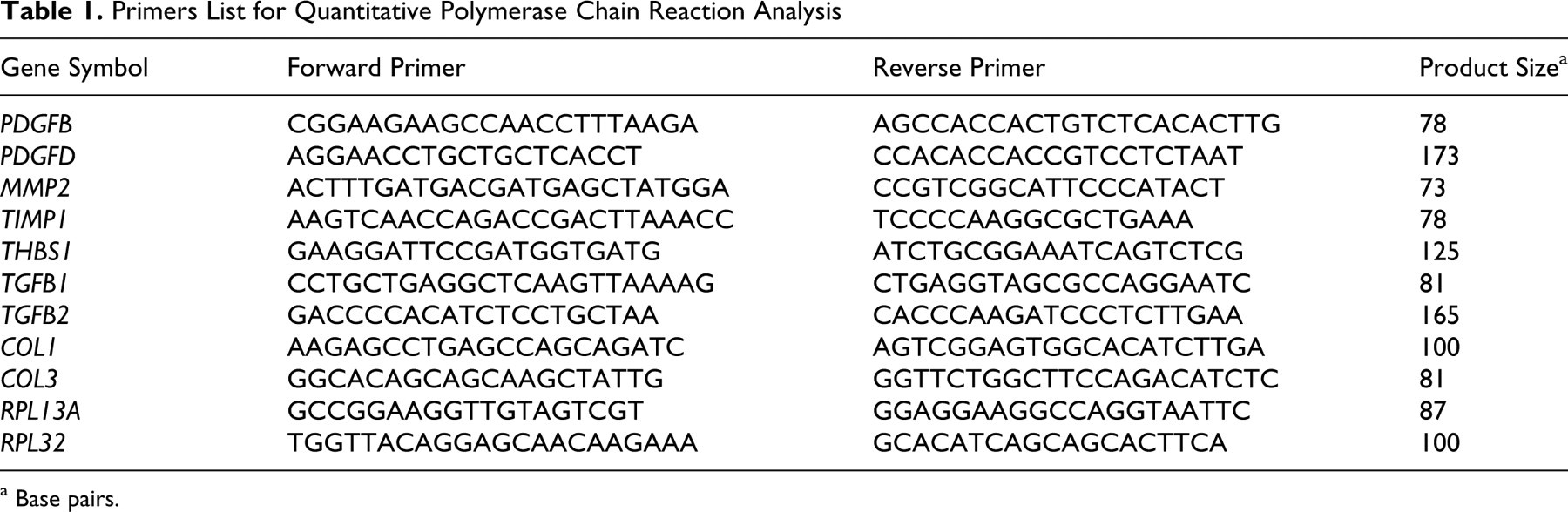
a Base pairs.
Initial incubation at 95°C for 10 seconds was followed by 40 polymerase chain reaction (PCR) cycles consisting of denaturation at 95°C for 5 seconds and annealing/elongation at 60°C for 30 seconds. The final annealing was carried out for 15 seconds at 95°C, followed by the melting curve reaction consisting of a gradual temperature increase from 60°C to 90°C.
The threshold cycle (Ct value) was determined using the second derivative maximum method. Before the samples were quantified, standard curves of the relative starting amount versus the Ct value were constructed using pooled liver cDNA. The amplification efficiency of each reaction was within 80% to 120%. The dissociation curve was confirmed to have a single peak in each experiment. Moreover, the PCR products were confirmed to be in agreement with their theoretic sizes by using agarose gel electrophoresis. The quantity of each transcript was determined by comparing the relative amount of the sample mRNA to that of RPL13A mRNA using the ratio of mean Ct values. The relative quantity of each transcript was confirmed in the same manner by using another housekeeping gene, RPL32 (the data presented here were collected using RPL13A as the internal control gene).
Histologic Procedures
Paraffin-embedded liver specimens were sliced into 4-μm-thick sections for hematoxylin and eosin (HE) staining, Sirius red staining, 13 and rubeanic acid staining. The degree of fibrosis was assessed semiquantitatively and assigned to 1 of 5 stages, with reference to the stages of fibrosis in human liver disease: 0, absent (no fibrosis); 1, mild (mild fibrous expansion of portal area); 2, moderate (moderate fibrous expansion of portal area); 3, severe (bridging fibrosis); 4, cirrhosis. 10 This staging system has been applied for the evaluation of fibrosis of canine hepatitis. 19
Statistics
The correlation between the transcript level for each gene and the degree of fibrosis was determined using the Spearman rank correlation test. The differences in expression of each gene between the dogs, with and without “poor prognostic” clinical signs (ie, ascites and/or jaundice 18 ), were determined using the Mann-Whitney U test. A P value less than .05 was considered significant. All statistical tests were performed using R (http://cran.r-project.org/).
Results: Histopathologic Evaluation and Correlation Between the Degree of Fibrosis and Gene Expression
With HE and Sirius red staining, the fibrosis among the chronic hepatitis/cirrhosis group had a wide range of severity (Figs. 1–6).
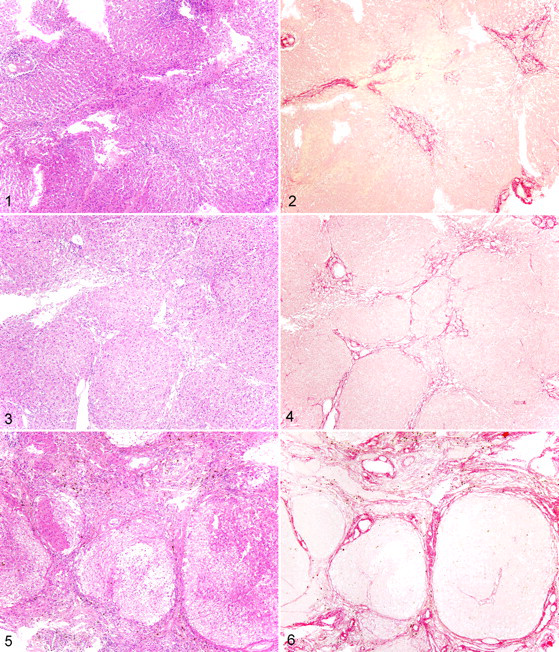
Liver; dog. Stage 2 fibrosis. Fibrous tissue is within portal areas. Bridging fibrosis is not present. HE.
Five dogs exhibited mild (n = 2, stage 1) or moderate (n = 3, stage 2) fibrosis (Figs. 1, 2). Most of these cases were also mild with regard to the severity of inflammatory cell influx or degenerative findings, with cell infiltration limited to the portal areas of the liver. In the liver parenchyma, few changes were observed. In the case of all the control dogs (n = 4), the findings of the histologic examination were normal (stage 0).
Five dogs had severe fibrosis with portal–portal bridging fibrosis (stage 3; Figs. 3, 4). Inflammation was observed primarily around the fibrous tissue or was surrounded by fibrous tissue but extended into the liver parenchyma. Most of the inflammatory cells were lymphocytes, with minor infiltration by macrophages and neutrophils. The severity of inflammation varied. Hepatocytes were degenerated or necrotic, and limiting plates were affected with some apoptotic bodies. A red pigment resembling copper was occasionally observed in the cytoplasm of the hepatocytes in HE and rubeanic acid staining.
Three dogs were diagnosed with cirrhosis (stage 4; Figs. 5, 6), with conversion of the normal liver architecture into pseudolobules and/or regenerative nodules. Mixed inflammatory cell infiltrates (macrophages, neutrophils, lymphocytes, and plasma cells), fibrosis, hepatocyte degeneration, apoptosis, necrosis, and hemorrhage were moderate to intermediate within the lobular or pseudolobular areas, but these changes were quite severe in the peripheral or fibrotic areas of the cirrhotic livers.
The relationship between the expression level of the fibrosis-related genes and degree of fibrosis is shown in Figure 7. The control samples were included in stage 0 fibrosis, and all the clinical samples had a fibrosis score equal to or greater than 1. The expression levels of all the genes were significantly correlated with the extent of histologic fibrosis (P < .01, r > .70).
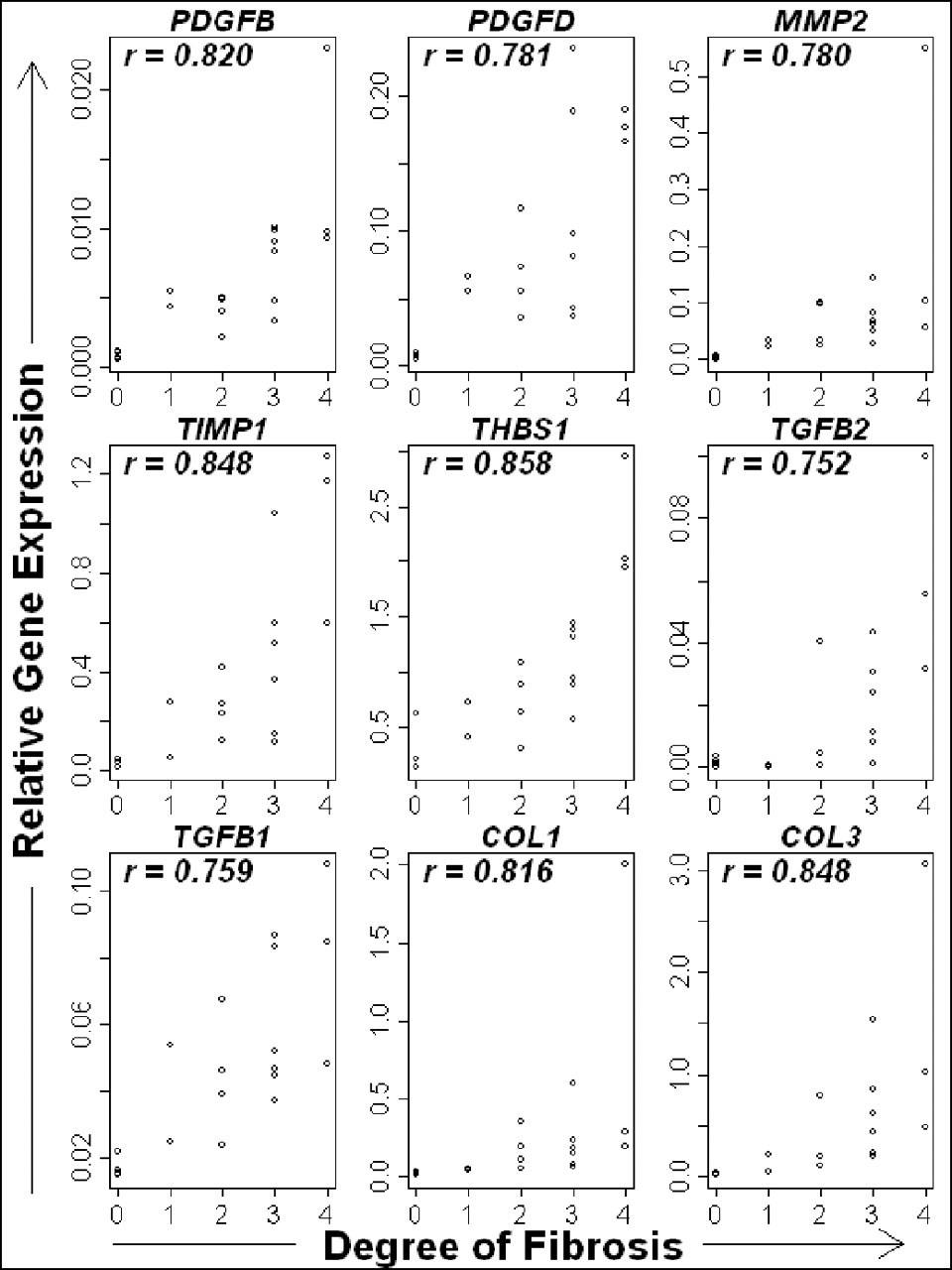
The relationship between degree of fibrosis and the expression of candidate fibrosis-related genes. The expression of all genes significantly (P < .005) and strongly (r > .70) correlated with the degree of fibrosis.
Ascites and jaundice have been identified as clinical signs indicative of a negative prognosis in canine liver diseases. 18 The expression levels of the 9 candidate genes in our study in dogs with and without ascites and/or jaundice are shown in Figure 8. Subgroup A included 7 dogs with either or both of these clinical signs (ascites only, n = 2; jaundice only, n = 1; ascites and jaundice, n = 4), whereas subgroup B (neither of the clinical signs) included 8 dogs. The expression levels of all the genes except TGFB1 were significantly (P < .05) higher in subgroup A (presence of ascites and/or jaundice) than in subgroup B (absence of both clinical signs).
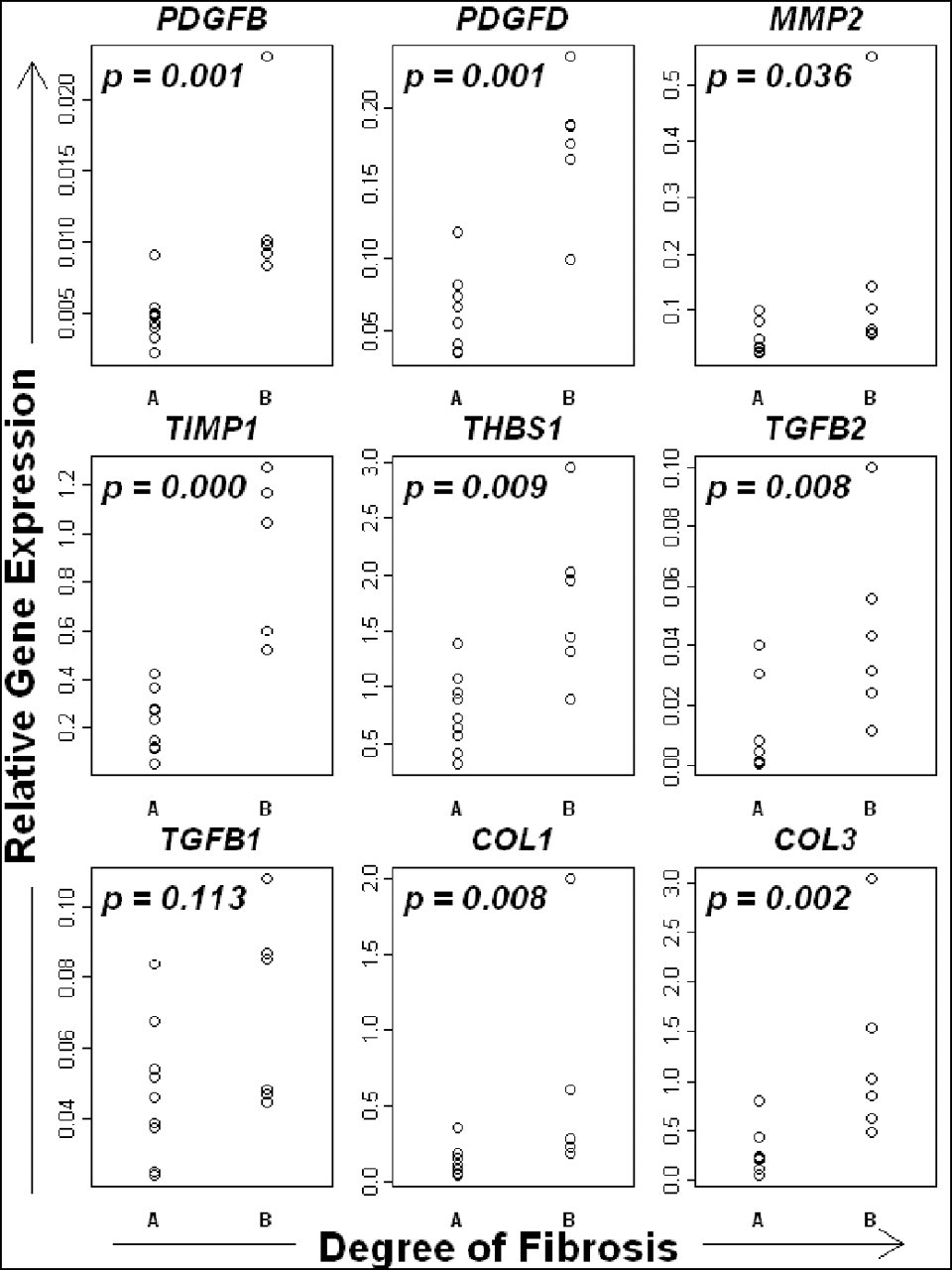
The expression of candidate fibrosis-related genes in cases of chronic hepatitis, with or without clinical signs related to poor prognosis (ie, jaundice and/or ascites). The expression of all genes examined was significantly higher in cases with clinical signs (subgroup B) than in those without them (subgroup A).
Discussion
Our data indicate that at the transcriptional level, MMP2, TIMP1, THBS1, PDGFB, and PDGFD are associated with hepatic fibrosis in dogs, in addition to TGFB and the collagen families of genes. This suggests that the molecular mechanisms underlying fibrosis in dogs are analogous to those in experimental models and in humans.
In canine chronic hepatitis, persistent inflammation, hepatocellular necrosis, and apoptosis trigger fibrosis. Increased expression of fibrogenic genes is temporally associated with clinical and histologic progression of liver disease. This finding suggests that as the disease progresses, the degree of fibrogenesis increases. Even in cirrhotic livers, expressions of the 9 genes that we examined were highly upregulated. These results are consistent with those of previous studies in dogs and other species. 4,14,15,17,20,23
If the cause of fibrosis (ie, chronic inflammation or fibrogenic processes) is eliminated, a reversal of hepatic fibrosis would be expected to occur in rodents and humans. 4 However, in most cases of dogs with chronic hepatitis, the cause of inflammation cannot be determined. 26 Moreover, because of the larger reserve capacity of the liver, the disease has progressed to the point that the patient responds poorly to anti-inflammatory treatment by the time that clinical signs emerge. Our results concerning the expression of a number of fibrosis-related genes in dogs with chronic hepatitis may aid the development of antifibrotic therapies for treating dogs and humans with liver diseases.
The results of this study may also contribute to the development of new markers of liver fibrosis. Although the extent of hepatic fibrosis can be assessed by histologic examination, the assessment is neither objective nor quantitative. Furthermore, it involves the assessment of only small pieces of liver samples. Instead, a serum marker can reflect the pathologic changes in the whole liver. To develop such a serum marker of liver fibrosis, it is necessary to quantify the products of the genes that we studied and elucidate the relationship of these genes with clinical and histologic features.
The positive correlation between TGFB1 mRNA level and the fibrotic stage indicates that TGFB1 plays a significant role in fibrosis. Unexpectedly, the expression level of TGFB1 was not significantly higher in dogs with poor prognostic signs (ie, later stage of fibrosis) than those without them. These results suggest that from the early stage of canine chronic hepatitis, the necroinflammatory response induces the TGFB1-mediated fibrogenic response. In fact, TGFB1 expression does not dramatically increase in the final stage of fibrosis compared to the earlier stage; the expression pattern of TGFB1 was different from the expression pattern of the other genes.
The present study has several limitations. First, we examined only gene expression, not protein expression. In addition, we used whole liver tissue rather than specific cellular fractions; therefore, our analyses included a variety of cell types. To properly quantify protein expression and determine which cells express the genes that we examined, further studies involving techniques such as immunohistochemical analysis, in situ hybridization, and laser microdissection would be helpful. The second limitation of our study is that all the dogs included were diagnosed with “chronic hepatitis.” There may be various subtypes of this disease in dogs. One of the major criteria of subclassifying hepatitis is the presence or absence of copper accumulation. 18,24 The present concentration of copper in the liver of the Labrador Retrievers and Doberman Pinschers, two breeds commonly affected by copper-associated hepatitis, were measured but not found to be high enough to diagnose these dogs with copper-associated chronic hepatitis. In the case of the other breeds, copper was not quantified. However, the results of rubeanic acid staining indicated that the histologic accumulation of copper was not as abundant as would be expected in cases of true copper-associated hepatitis and that copper was localized mainly in the periportal areas (data not shown). These findings suggest that the dogs in this study should be classified as idiopathic.
In conclusion, expression of the fibrosis-related genes PDGFB, PDGFD, MMP2, TIMP1, THBS1, COL1A1, COL3A1, TGFB1, and TGFB2 were found to correlate well with the histologic degree of fibrosis and clinical disease progression. The results of this study suggest that the mechanism of hepatic fibrosis in dogs is similar to that in other species, and they provide information that may be useful for developing strategies to treat hepatic fibrosis in dogs.
Footnotes
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
This work was supported by Grant-in-Aid for Scientific Research (B) of Japan Society for the Promotion of Science (KAKENHI 21380193).