
Abstract
Tumors of the jaw bones and oral soft tissue are relatively common lesions in dogs. The aim of this study was to find cell markers to differentiate odontogenic epithelium from nonodontogenic epithelium for future research on the pathogenesis and pathology of odontogenic neoplasms in dogs. Keratin 14 and 19 staining was observed in odontogenic and nonodontogenic epithelium, whereas amelogenin and p75 neurotrophin receptor immunoreactivity was observed in certain odontogenic epithelial cells at various stages of development but not in other epithelial cells. Calretinin staining was observed in the alveolar epithelial cells directly overlying the developing tooth germ in 28 of 39 sections (71.8%), as well as the dental laminae in 30 of 35 sections (85.7%) and Serres rests in 24 of 28 sections (85.7%). Focal positivity was detected in the respiratory mucosa, some hair follicles, and fusion epithelium of the palate, but no calretinin staining was observed in other oral epithelial cells; therefore, calretinin has potential to be utilized as a marker to differentiate odontogenic form nonodontogenic epithelium.
Keywords
Dogs frequently present with oral epulides defined as tumors of the gingiva. Many of these can easily be identified as one of the commonly recognized odontogenic neoplasms. 13,38,39 Problematic entities, however, do exist when the tumors do not have the classic histological features of any specific odontogenic tumor or cyst. In an attempt to elucidate the origin and character of these epulides of uncertain histogenesis, it is imperative to find molecular markers that could discriminate between odontogenic epithelium and other types of epithelium in the mouths of dogs. Such markers could then be utilized in future molecular research on the pathogenesis and pathology of odontogenic versus nonodontogenic neoplasms in dogs. To the best of our knowledge, no in situ markers for odontogenic epithelium have been described in developing dog teeth. We therefore evaluated keratin 14 (K14) and 19 (K19), amelogenin, p75 neurotrophin receptor (p75NTR), and calretinin on developing dog teeth—all of which have been described as markers of odontogenic epithelium in developing human and rat odontogenic tissues. 7 –9,19,26
Materials and Methods
Twenty-four fetuses of large-breed dogs were obtained under ethical clearance of the Animal Use and Care Committee of the Faculty of Veterinary Sciences, University of Pretoria, South Africa, from female dogs scheduled for elective pregnancy termination. The fetuses were fixed in 10% buffered formalin and the heads carefully cut into coronal sections, dehydrated, and embedded in paraffin wax blocks. Those samples that contained calcified bone or dental hard tissues were decalcified in routine decalcifying solution (70 ml HNO3, 50 ml HCL, 880 ml distilled water) for 60 minutes and then rinsed in running tap water for 60 minutes. Tissue specimens were sectioned at 3 μm, stained with hematoxylin and eosin, and microscopically examined to select slides with well-formed enamel organs in which the respective odontogenic epithelial cells were morphologically clearly identifiable.
For the purpose of this study, the internal enamel epithelium (IEE) was defined as short columnar cells with centrally placed nuclei situated opposite undifferentiated ectomesenchymal cells of the dental papilla without any signs of odontoblast differentiation or dental hard tissue formation (Fig. 1 ). Secretory ameloblasts were defined as tall columnar cells with prominent reversed polarity of the nuclei in close approximation to formed enamel and dentin. Presecretory ameloblasts were defined as tall columnar cells with reversed polarity but without the presence of dental hard tissue adjacent to these cells. The cells of the outer enamel epithelium (OEE) were defined as low cuboidal cells with little cytoplasm at the outer periphery of the enamel organ. The oral epithelium on the alveolar ridge area, from where the tooth germ originated, was referred to as the overlying alveolar epithelium. The term dental lamina was used when there was continuity between the overlying oral epithelium and the developing tooth germ, whereas discrete islands of epithelial cells in this location were classified as Serres rests.
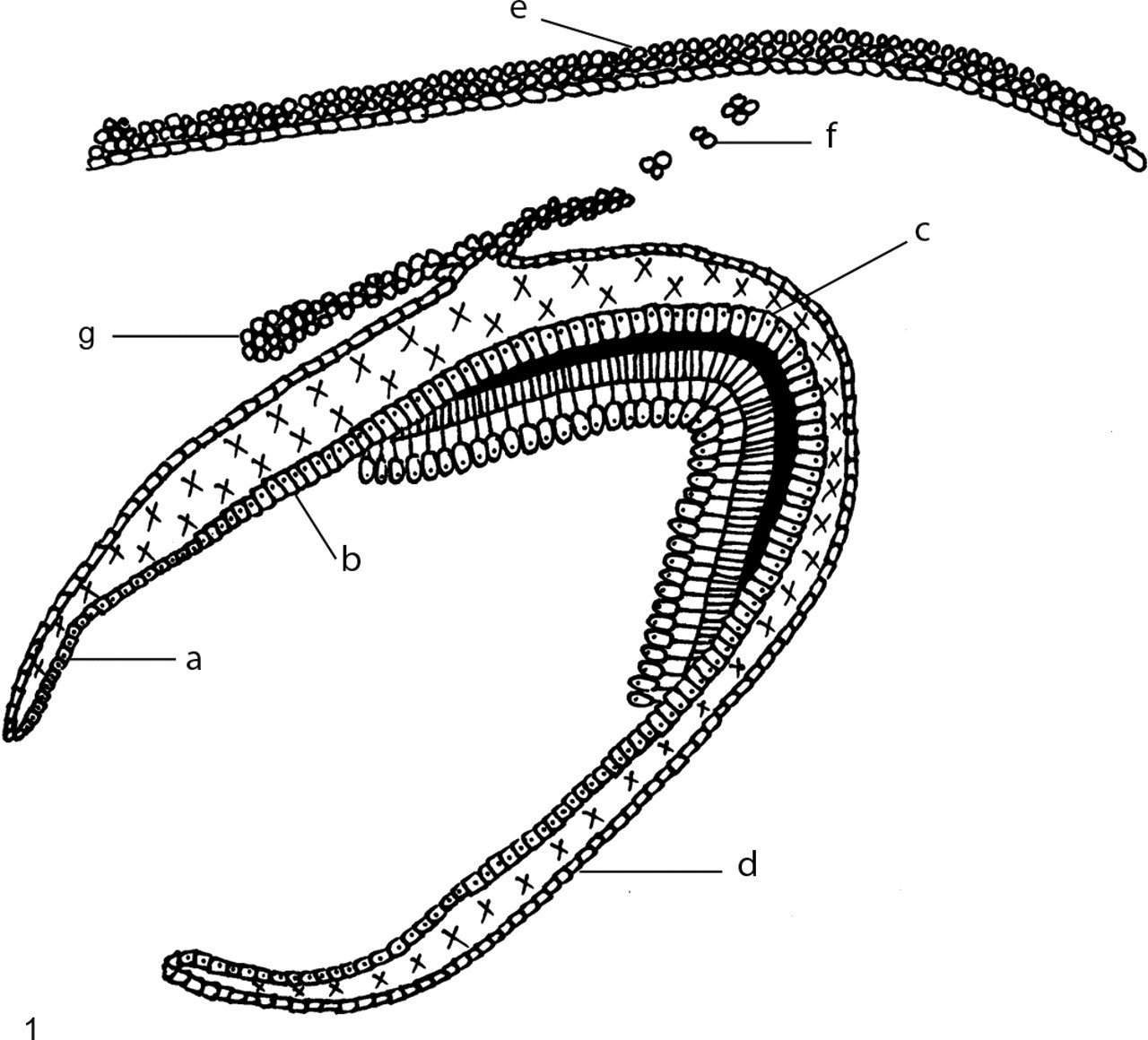
Diagrammatic representation of cells in developing tooth germ: a, internal enamel epithelium; b, presecretory ameloblasts; c, secretory ameloblasts; d, outer enamel epithelium; e, overlying alveolar epithelium; f, Serres rests; and g, dental lamina.
The chosen tissue sections were dewaxed in xylene and hydrated with graded alcohol solutions. Heat-induced epitope retrieval in citric acid buffer (pH 6) was performed on the sections for the low-affinity nerve growth factor receptor (NGFR) (gp75, Novocastra NCL-NGFR, Novocastra Laboratories Ltd, Newcastle Upon Tyne, UK), cytokeratin 14 (Novocastra NCL-LL002), and amelogenin (FL-191, Santa Cruz Biotechnology sc-32892, Santa Cruz Biotechnology, Inc, Santa Cruz, CA) using a Pascal pressure chamber (Dako Cytomation, Inc, Carpintera, CA) according to the manufacturer’s instructions. Sections were cooled for 20 minutes at room temperature and treated with hydrogen peroxide for 5 minutes at 37°C to quench endogenous peroxidase activity. Sections stained for calretinin (Novocastra NCL-Calretinin) and cytokeratin 19 (Novocastra NCL-CK19) were first treated with hydrogen peroxide; then, heat-induced epitope retrieval was done in ethylenediaminetetraacetic acid buffer (pH 8) using the Pascal.
Sections were then incubated in their various antiserum: cytokeratin 14 (1:60), cytokeratin 19 (1:100), NGFR-gp75 (1:50), calretinin (1:100), and amelogenin (1:50) for 60 minutes at room temperature. All the sections, except the amelogenin, were then incubated in Envision + System-HRP Labelled Polymer Mouse (Dako K4000, Dako Cytomation, Inc) for 35 minutes at room temperature. The amelogenin sections were incubated in Envision + System-HRP Labelled Polymer Rabbit (Dako K4009, Dako Cytomation, Inc) for 35 minutes at room temperature. All the sections were then stained with AEC+ chromogen (Dako Cytomation, Inc) for 4 minutes at 37°C. Sections were counterstained with hematoxylin, mounted with Faramount Aqueous Mounting Media (Dako S3025, Dako Cytomation Inc) and then examined microscopically.
Results
The exact gestation time of the developing dog fetuses was unknown, so they were classified into two groups based on their stage of odontogenic development. Fetus Nos. 1–8 (group 1) had tooth germs in only the bud and cap stages of development, with no ameloblast differentiation or dental hard tissue formation. The tooth germs of fetus Nos. 9–24 (group 2) were in the bell stage of development, with visible cell differentiation and hard tissue formation. In 15 cases, a rostral (anterior) section and a caudal (posterior) section of the same fetus were used for comparison. In the remaining 9 cases, only one section complied with the inclusion criteria. A total of 39 sections were therefore harvested from the 24 fetuses. Although the number of tooth germs on a single section varied from 1 to 4, only 1 was chosen for analysis, defined as the best representative of the bell stage in the given section.
Calretinin
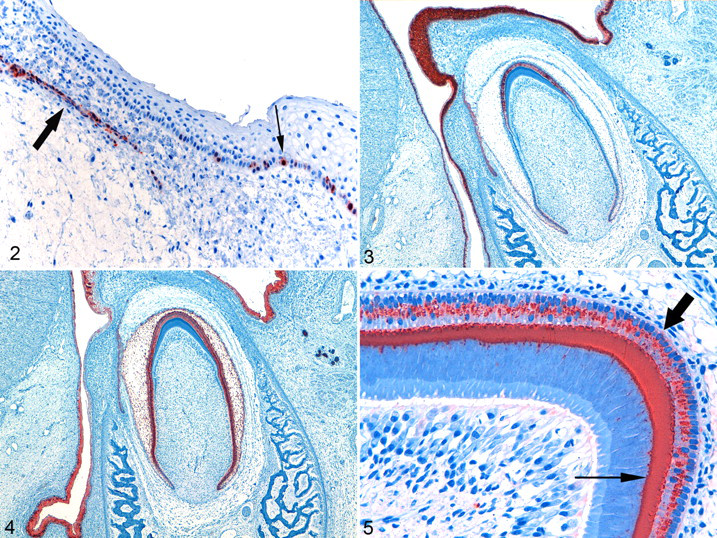
Group 1 fetuses showed calretinin staining of the OEE, but no calretinin staining was observed in the OEE of group 2; furthermore, no staining was observed in the IEE, ameloblasts, odontoblasts, or epithelial cells of the stellate reticulum or stratum intermedium at any stage of development (ie, in either group). Of the 35 dental laminae evaluated, 30 stained positive for calretinin (85.7%). Of 28 cases with Serres rests, 24 showed positive staining in the epithelial rests (85.7%). The overlying alveolar epithelium, in close approximation to the lamina and Serres rests, stained positive in 28 of the 39 cases evaluated (71.8%) (Fig. 2 ).
Keratin 14
Intense and diffuse K14 staining was present in the IEE of 5 of 5 group 1 fetuses, whereas the group 2 fetuses mainly showed focal staining in the IEE in 20 of the 22 cases examined (91%). No staining of presecretory ameloblasts could be seen in group 2, although intense and diffuse staining of the secretory ameloblasts was seen in all cases. The staining intensity of the secretory ameloblasts decreased as enamel secretion continued. The OEE in all sections (36 of 36) stained positive, although the distribution and intensity varied. Intense and diffuse staining of the stellate reticulum (6 of 6) and stratum intermedium epithelium (5 of 5) was seen in group 1. In group 2, 15 of 30 cases (50%) showed positive staining in the stellate reticulum, and 14 of 29 (48%) showed positive staining in the stratum intermedium epithelium. Intense staining patterns were observed throughout the full thickness of the alveolar epithelium, dental laminae, and Serres rests in all cases examined. No positive staining was detected in any of the odontoblasts present in the sections (Fig. 3).
Keratin 19
In group 1 and group 2, the epithelial cells of the enamel organs of all the sections stained positive with variable intensities and distribution patterns. The IEE cells of group 1 exhibited a heterogeneous pattern of staining, given that staining intensities varied. In group 2, diffuse expression of K19 was observed in the IEE cells, except in the cervical loop region. The presecretory and secretory ameloblasts stained diffusely positive in all cases, although with varied intensities. Positive staining was observed in the full thickness of the overlying alveolar epithelial cells in all the group 1 fetuses, but in group 2, it was mainly restricted to the superficial epithelial cell layers. Diffuse staining was observed in all the dental laminae and Serres rests in all cases examined. No positive staining was detected in any of the odontoblasts present on the sections (Fig. 4).
Amelogenin
No amelogenin staining was present in group 1. In group 2, small amounts of amelogenin were observed in presecretory ameloblasts seen in close approximation to the cell nucleus. Diffuse granular staining for the amelogenin protein was observed in all secretory ameloblasts present in group 2 sections. No staining was detected in the IEE, OEE, stellate reticulum, stratum intermedium, dental laminae, Serres rests, or alveolar epithelium of both groups. Of the 30 sections in group 2 evaluated, 6 revealed weak staining in some odontoblasts (20%). Staining seemed to be localized to the intercellular spaces between the odontoblasts and other pulp cells. Staining of these sections was repeated, but the same results were obtained. No amelogenin staining was observed in or around odontoblasts at advanced stages of odontogenesis (Fig. 5).
p75NTR
No epithelial staining was observed in bud-stage dental organs. Focal positivity was observed in the IEE of 5 of 5 cap-stage germs, as well as in the region of the cervical loops of 9 of 28 bell-stage tooth organs (32%). No staining was observed in OEE, stellate reticulum, stratum intermedium, ameloblasts, dental lamina, Serres rests, or overlying alveolar epithelium. In 22 of 27 cases (81%), some preodontoblasts stained positive. As soon as the odontoblasts differentiated, the positivity disappeared. Faint positivity was observed throughout the pulp cells in 3 of 5 cap-stage tooth germs (60%), but during the bell stage in 21 of 28 tooth germs (75%), staining was restricted to the apical region of the pulp only. The dental follicle cells stained diffusely positive throughout the evaluated sections.
Discussion
Calretinin staining could be demonstrated only in the epithelium of the dental laminae, Serres rests, and associated overlying alveolar epithelium (Fig. 2, Table 1 ), whereas Mistry and coworkers described different levels of calretinin expression in the dental lamina, OEE, stellate reticulum, stratum intermedium, IEE, and ameloblasts of rat teeth. 22 To the best of our knowledge, their study was the only one describing calretinin expression in developing odontogenic tissue. They suggested calretinin, a calcium-binding protein, to play a role in enamel formation because calretinin expression remained negative in the molar teeth over the cusp tips, where enamel is never formed. 22 Differences in antibodies and techniques used, differences in species, 2,32 and differences in the developmental stages of the fetuses examined could offer some possible explanations for the differences in results found. 34 In the current study, focal positivity was detected in the respiratory mucosa, some hair follicles, and fusion epithelium of the palate. We hypothesize that calretinin could be a useful marker for odontogenic epithelial cells, given that the positive staining found in the overlying alveolar epithelium could not be detected in the rest of the oral epithelial cells. From this study, we propose that calretinin should be investigated further in this respect.
Expression Patterns Observed in Dog Odontogenic and Nonodontogenic Tissue a

a +, focal positive staining observed; ++, diffuse positive staining observed; –, no positive staining observed.
b Secretory.
c Presecretory and secretory.
d During mantle dentin formation.
e Preodontoblasts.
K14 is a type I intermediate filament said to confer physical resilience to basal epithelial cells. 6 It is the main intermediate filament of odontogenic epithelium and a marker often utilized in the detection of ameloblasts and IEE. Immunohistochemical studies have shown strong K14 positivity in IEE of human fetuses at the early bell stage, 9 with weaker staining in fully differentiated ameloblasts at a later developmental stage. 9,44 In contrast, others have shown the IEE cells of rat tooth germs in the earlier stages to have weak expression of K14, with more intense staining in preameloblasts and even more so in fully differentiated ameloblasts. 35 The staining patterns of K14 in the current study are unique. Intense and diffuse staining was observed in the IEE of all the group 1 fetuses, whereas the same cells in the group 2 fetuses revealed only focal staining in 20 of 22 cases (91%). No K14 staining was seen in any presecretory ameloblasts, but this changed to intense and diffuse staining of all secretory ameloblasts in all cases (Fig. 3), which again decreased as enamel secretion continued. This phenomenon of decreased and ultimately no K14 staining in the presecretory ameloblasts was difficult to explain. K14 has been described as an anchorage protein that might have a role in supporting and preserving the epithelial–mesenchymal interactions. 9,11 It has been speculated that ameloblasts lose K14 to disengage its anchorage to migrate for enamel matrix deposition. 9 The dynamic nature of intermediary filament expression as well as the occurrence of selective or complete masking of the intermediate filament determinant 12 could offer more explanations for our observed staining pattern. However, the mechanism of the in vivo regulation of keratins remains to be determined. It is possible that the various authors' definitions of preameloblasts, presecretory ameloblasts, and secretory ameloblasts differ from one another and therefore offer another reason for the differences in the respective results found. This study showed K14 expression throughout the odontogenic epithelium, with no staining in the dental mesenchyme (Table 1); but staining of the odontogenic epithelium was not exclusive, because positivity was also observed in the other types of epithelium present on the coronal sections (oral epithelium, skin, salivary glands and ducts, some respiratory mucosa).
K19, another type I intermediate filament, has been described in the epithelium of the stellate reticulum and stratum intermedium, secretory ameloblasts, and ameloblasts in the maturation stage of monkeys (Macaca mulatta and Macaca nemestrina). 21 In human fetuses, K19 (in association with K8) has been described in all cells of the dental lamina and enamel organ, 9,16 with intense positive staining in the fully differentiated ameloblasts. 9 For this reason, K19 has been postulated to be a marker of ameloblast differentiation. 9 Our study agreed with others that found K19 staining patterns in the IEE to vary from weak single cell to intense and diffuse but with constant diffuse and intense staining of the presecretory and secretory ameloblasts (Fig. 4). 9 Positive staining for K19 was observed in hair follicles, salivary gland ducts, respiratory mucosa, and oral epithelium; therefore, K19 did not stain odontogenic epithelium exclusively and could not be regarded as a specific marker for odontogenic epithelium (Table 1). Because the pattern of staining for K14 and K19 appeared to change with stage of development, it is possible that another profile will be seen in adult tissues and should thus be investigated further.
Amelogenins are enamel matrix proteins secreted by ameloblasts; they constitute more than 90% of the developing enamel extracellular matrix during odontogenesis. 10,25,36 In the past, amelogenin was widely accepted as an enamel-specific protein 1,17,30,37,42 that was proposed and used as a stage-specific marker for ameloblast differentiation. Its expression was not detected in IEE, although expression was found in preameloblasts and ameloblasts. 8,35,44 However, amelogenin was detected in hamster mantle dentin, 18 which was proposed to be the result of diffusion or translocation of amelogenin into the predentin and preodontoblast layer with endocytosis of the amelogenin protein by the odontoblasts. 18,27 It has also been suggested that the presence of trace amounts of amelogenin splice products in dentin could be the result of contamination. 28 Since then, amelogenin mRNA was detected in odontoblasts of porcine and rat teeth, 29,31 and in 2006 Ye et al suggested that odontoblasts actually synthesize and secrete amelogenins, as based on in situ hybridization and immunohistochemistry studies of human odontogenic tissue. 45 The mRNA of splice forms of amelogenin have been detected in periodontal ligament cells (cementoblasts) of wild-type mice, 14 and amelogenin mRNA and protein was recently described in dog and rat osteoblasts and osteoclasts, in some osteocytes, and in articular cartilage chondrocytes. 15 It was therefore proposed that amelogenin is expressed in mesenchymal cells and is not only an ameloblast-specific protein as previously believed. 14,15
Expression of the amelogenin protein in this study was similar to that of previous studies that found it to vary from only small amounts in presecretory ameloblasts 17,30 to diffuse positivity throughout differentiated ameloblasts in the secretory stage (Fig. 5). 17,30,37 Single-cell positivity of odontoblasts was observed in 6 of 30 sections (20%) with odontoblasts present. On close examination, the staining appeared to be localized to the intercellular spaces. It is therefore possible that the single-cell positivity observed in the odontoblasts in our study could in fact be artifactual. The amelogenin protein actually located outside the cell could have been misinterpreted as intracellular staining owing to superimposition on the slide. No staining was observed in odontoblasts at advanced stages of odontogenesis, and the single cells that did seem to stain were associated with the mantle dentin only. Although staining has been described in other tissues, 15 in this study amelogenin was detected only in odontogenic tissues, and it seems to be a promising marker to distinguish between odontogenic and nonodontogenic tissues in dogs (Table 1).
In 1985, Buxser et al observed that two classes of NGFRs exist—namely, high- and low-affinity receptors, 4 which are today better known as the Trk (tropomyosin-related kinase) tyrosine kinase receptors (high affinity) and the p75NTR (low affinity). 33 In fact, p75NTR has been described in epithelial cells of the growing tooth germ, as well as in the associated ectomesenchymal cells of rat teeth. 5,23,24 In the bell stage, staining has been observed in proliferating cells of the IEE, 5,23,43 in some cells of the stratum intermedium, and in preodontoblasts and polarizing odontoblasts. 5,23,24 More recent studies have revealed immunoreactivity against p75NTR to be restricted to the IEE, the dental papilla, and the dental follicle of rat incisors; therefore, p75NTR has been used as a marker for IEE cells. 19,26 In the current study, p75NTR expression was restricted to focal regions of the IEE in 14 of 33 sections (42%), and no staining was observed in other epithelial cells (Table 1). It could therefore be utilized as a marker to differentiate between odontogenic epithelium in the early stages of development versus nonodontogenic epithelium. The widespread staining observed in the mesenchymal tissue does not permit p75NTR to be a specific marker for odontogenic epithelium, although the morphological differences between epithelium and mesenchymal tissue would aid in the histological investigation. The precise function of p75NTR expression in the IEE cells remains a speculative topic.
Odontogenic tumors may originate from the epithelial or ectomesenchymal cells of the developing tooth germ or its remnants. 20 Most odontogenic tumors are found in adult dogs where all that is left of the odontogenic epithelium is the Serres and Malassez odontogenic epithelium rests and, in certain cases, the reduced enamel epithelium associated with an unerupted (or impacted) tooth. One can therefore assume that the odontogenic tumors in adult dogs probably originate from these epithelial remnants in the gingiva and the periodontal ligament space. 3 As shown in this study, the expression profile of certain markers was altered in group 2 compared to group 1 in many respects. This expression profile may change even more in postnatal and adult tissue as previously indicated. 34,40,41,43 Therefore, it is possible that a marker could retain its expression in Serres and Malassez rests but lose its expression in oral epithelial cells that lost their odontogenic potential. We propose that the oral epithelial cells gain more mature differentiated characteristics in adult tissue as opposed to the Serres and Malassez rests, which may retain their original fetal characteristics and expression profiles, given that they are quiescent remnants of fetal developmental cells. Finding a suitable marker that will stain the odontogenic rests without staining the remainder of the oral epithelial cells in adult tissue could aid in finding a marker to distinguish between tumors originating from odontogenic tissue compared to those taking origin from the adjacent oral epithelium. The expression of the same markers used in this study should therefore be tested in adult dog tissue, specifically to evaluate the immunoreactivity of the Serres and Malassez odontogenic epithelium rests and associated gingiva.
Footnotes
Acknowledgements
We would like to thank the Veterinary Academic Hospital at Onderstepoort and Dr M. F. Visser (Lakeside Veterinary Clinic) for providing us with the canine fetuses used in this study.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.