
Abstract
An outbreak of diarrhea on a large commercial mink farm affected 5,000 of 36,000 neonatal mink kits, with 2,000 dying within a 2-week period. Affected kits were severely dehydrated, and their furcoats and paws were covered with yellow- to green-tinged mucoid feces. On necropsy, the small intestines of examined animals were markedly distended by serous to mucoid fluid. Microscopically, there was prominent colonization of the intestinal villar epithelium by gram-positive bacterial cocci in the absence of inflammation and morphologic changes in villous enterocytes. The colonizing bacteria were phenotypically identified as belonging to the Staphylococcus intermedius group of bacteria. This was confirmed by nucleic acid sequence analysis of the 16S ribosomal RNA gene. Further nucleic acid sequencing of polymerase chain reaction (PCR) amplicons from the superoxide dismutase gene and the heat shock protein 60 gene differentiated the isolate as Staphylococcus delphini. Production of staphylococcal enterotoxins A and E was demonstrated with a commercial ELISA-based immunoassay. Sequencing of PCR amplicons confirmed the presence of the enterotoxin E gene, but PCR amplification of the enterotoxin A, B, C, or D genes was not successful. Although direct causation was not confirmed in this study, the authors postulate that the observed hypersecretory diarrhea in these mink kits was the result of colonization of the small intestine by S delphini and subsequent production of enterotoxin.
Preweaning diarrhea is common in farm-raised neonatal mink and has been recognized to occur in outbreak situations with high morbidity and mortality. 4 As such, neonatal diarrhea is an economically significant disease on many mink ranches. Owing to the mucoid to tacky nature of the diarrheic feces and secondary contamination of the fur of affected animals, this pattern of neonatal diarrhea is often referred to as “sticky kits,” “greasy kits,” or “wet kits” syndrome. Excessive secretion from the cervical apocrine glands is often reported concomitant with diarrhea, adding to the wetness and stickiness of affected kits. 4 The onset of clinical signs generally occurs preweaning, at 1 to 4 weeks of age. 4 The cause of this syndrome is unknown and may be multifactorial. Infectious agents that have been identified in affected kits include Campylobacter jejuni, Escherichia coli, reovirus, calicivirus, and astrovirus. 4,6,7,14 Many of these agents have also been identified in clinically normal kits, so the significance of their presence in affected kits is uncertain. Other factors that have been suggested to play a role in previous outbreaks include environmental factors such as overcrowding, suboptimal hygiene, the age of the dam, and the presence of mastitis in dams. 4
This report describes the association of colonization of the small intestine by staphylococci with neonatal diarrhea in mink that presented with clinical signs consistent with sticky kit syndrome. Furthermore, this report represents the first description of disease associated with Staphylococcus delphini in mink and indicates that some isolates of S delphini may possess genes that produce enterotoxin, as demonstrated by PCR and ELISA.
In May 2004, an Idaho-based commercial mink ranch experienced a sudden outbreak of diarrhea in preweaning kits. This farm was composed of approximately 7,200 breeding females with an estimated 36,000 kits. Animals were housed in blocks of cages in a total of 30 whelping sheds. Each cage contained a single female and her litter. Sheds had open sides and were separated from one another by 14 feet (4 m). Kits were affected in all sheds, but not all sheds were equally affected.
The first signs of disease were noted on 7 May 2004, when mothers of 5 to 6 litters appeared agitated and were unable to keep kits clean. Disease rapidly spread throughout the farm, affecting all sheds to variable degrees, with a peak in disease incidence on 11 May 2004. Bedding was changed every other day in nest boxes containing affected kits, and kits were sprinkled with a powdered form of oxytetracycline. Females were treated with a combination of penicillin, sulfadiazine, and tetracycline in the feed. By 19 May 2004, twelve days after the first signs of disease were noted, the outbreak seemed to have subsided. Aside from general agitation, dams were unaffected and showed no signs of mastitis, dermatitis, or diarrhea.
Affected kits generally ranged from 10 to 14 days of age. Fur and paws of affected litters were sticky owing to contamination by yellow- to green-tinged mucoid feces. Kits appeared agitated with excessive vocalization. Clinical signs often rapidly progressed to dehydration, lethargy, and death. Many kits were found dead without premonitory clinical signs. In total, an estimated 5,000 kits were clinically affected, 2,000 of which died. These numbers correspond to a 13.9% morbidity rate in the entire population and a 40% mortality rate within affected kits.
Similar outbreaks of neonatal diarrhea had reportedly occurred in 2 of the previous 10 years at this farm. In previous outbreaks, affected kits were older, being between 3 and 4 weeks of age. According to the rancher, the outbreak described here began the day after a sudden change in weather. Several days before the outbreak, the weather had been warm and dry, with average high temperatures near 80°F. The day immediately before the first noted clinical signs, the high temperature dropped to below 70°F and it rained. Many of the nest boxes were reported to be cool and damp. Whether or not this contributed to the outbreak is unclear. It is possible that this sudden change in weather resulted in some degree of stress to the kits and the dams, and dampness in the nest box could have predisposed for the environmental overgrowth of bacteria.
Materials and Methods
Necropsy
Over the course of the outbreak, a total of 23 kits were submitted to the Michigan State University Diagnostic Center for Population and Animal Health, East Lansing, Michigan. Twenty-two of these animals were frozen and submitted whole for necropsy. A field necropsy was performed on 1 animal. Collected tissues from this kit were fixed in a commercial alcohol-based fixative (Z-Fix, Anatech Ltd, Battle Creek, MI) and submitted to the center for histopathologic and ultrastructural examination. Kits necropsied at the center were divided into 4 groups based on the litter of origin for histopathologic examination and microbial testing (Nos. 3113870-1 [6 kits], 3113878-1 [6 kits], 3113878-2 [6 kits], and 3113878-3 [4 kits]).
Histopathology
Representative samples of liver, lung, heart, kidney, spleen, mesenteric lymph node, adrenal glands, thyroid, cerebrum, cerebellum, brain stem, stomach, pancreas, proximal, middle and distal small intestine, and colon from all groups were fixed in 10% buffered formalin and routinely processed and paraffin embedded. Five-μm sections of all tissues were stained with HE, and sections from several portions of small intestine were Gram stained.
Transmission Electron Microscopy
For ultrastructural analysis, sections of small intestine that had been fixed in Z-Fix were dehydrated in graded alcohols and embedded in Poly/Bed 812-Araldite resin (Polysciences, Inc, Warrington, PA) by routine methods for transmission electron microscopy. Thin sections were cut and stained with methylene blue for selection of areas to be processed for ultrastructural examination. Ultrathin sections were cut and stained with uranyl acetate and lead citrate and examined on a Philips 301 electron microscope.
Bacterial Culture
For bacterial culture, representative sections of small intestine were pooled according to litter number. For each batch of pooled small intestine, intestinal contents were plated onto 5% enriched sheep-blood agar, Columbia colistin-nalidixic acid agar with 5% sheep blood for selective isolation of gram-positive cocci and inhibition of gram-negative bacilli, and MacConkey agar medium selective for gram-negative bacilli and differential based on lactose fermentation. Intestinal contents were also plated on brilliant green agar with novobiocin and inoculated in tetrathionate broth for the enriched and selective growth of Salmonella species other than Salmonella typhi and Salmonella paratyphi. Isolated staphylococci were further tested for growth on P agar and the ability to produce acetoin (VP test).
ELISA Testing for Enterotoxins
Crude extracts of cultures of the Staphylococcus intermedius isolates from small intestinal batch Nos. 3113870, 3113878-1, and 3113878-3 were prepared by filtration (0.22-μm Millipore filter, Millipore, Billerica, MA) of overnight cultures in tryptic soy broth. These batches were selected because of adequate bacterial load in the samples (> 100 colony-forming units) and histological evidence of mucosal colonization. Extracts were screened for the production of enterotoxins with a commercial ELISA-based system (RIDASCREEN SET A, B, C, D, E kit; R-Biopharm GmbH, Darmstadt, Germany) as previously described. 15 The employed ELISA system utilizes monovalent capture antibodies against staphylococcal enterotoxin types A through E (abbreviated SEA to SEE).
Polymerase Chain Reaction Fragments and Nucleic Acid Sequence Analysis
A series of polymerase chain reaction (PCR) assays was done to characterize the Staphylococcus isolated from batch No. 3113870. Briefly, DNA was extracted from several bacterial colonies and stored frozen at –20° until use. The DNA was amplified using standard procedures; the PCR products were separated by electrophoresis through 1.5% agarose gels containing ethidium bromide; and PCR amplicons were visualized with the aid of ultraviolet light. A fragment of the 16S ribosomal RNA gene was amplified with the forward primer AGAGTTTGATCMTGGCTCAG and the reverse primer GGGTTGCGCTCGTTR, as previously described. 20 A fragment of the superoxide dismutase (sodA) gene was amplified using forward primer CACATTGATAAAGAAACTATGG and reverse primer TATCTTGGTTTGSAGTTGTAA. A fragment of the heat shock protein 60 (hsp60) gene was amplified with forward primer GCACAAGCRATGATTCAAGAAG and reverse primer CTGTYAATGTACCACGCATACG. The PCR primers used for amplification of fragments of the Staphylococcus aureus enterotoxin genes SEA, SEB, SEC, SED, and SEE were as previously published. 5 The PCR amplicons were cut from gels, purified, and submitted to the Research Technology Support Facility at Michigan State University for nucleic acid sequencing. Sequences were edited with Sequencher software (Gene Codes Corporation, Ann Arbor, MI), trimmed to remove primer sites, and analyzed with BLAST (Basic Local Alignment Search Tool), available through the National Center for Biotechnology Information.
Additional Diagnostic Testing
PCR for coronavirus and rotavirus was preformed as previously described. 21,22
Results
Necropsy and Histopathology
Gross and histologic changes were similar for kits from all litters. Bedding and scant tacky, light green fecal material was adherent to the limbs and perineum of most animals. The fur of the entire body, particularly in these areas, was damp and matted. Animals were moderately to markedly dehydrated. The abdomens of many kits were mildly to moderately expanded owing to marked, diffuse distention of the small intestines. The small intestines and, to a lesser degree, the colons were filled with pale white to yellow-tinged, turbid, serous to mucoid ingesta and mild amounts of gas (Fig. 1).
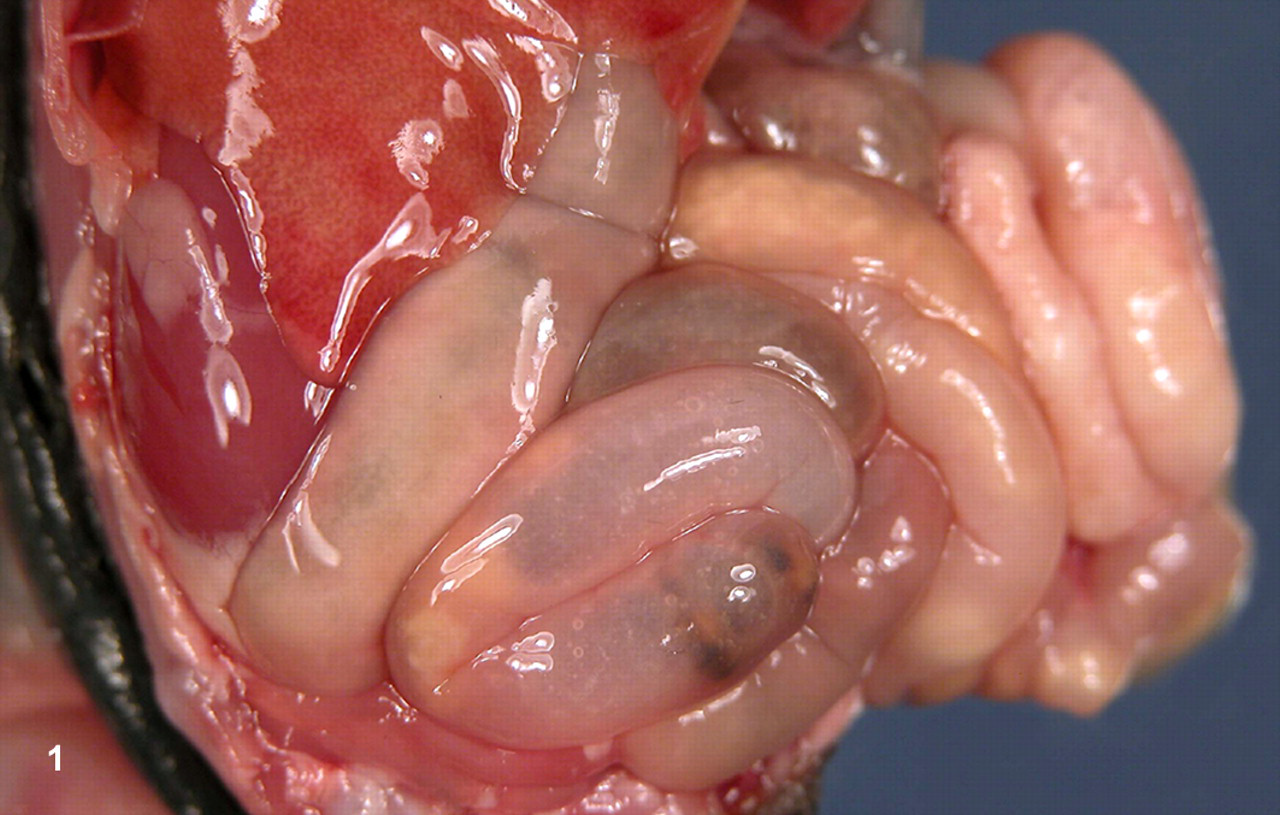
Abdominal cavity. The small intestines are diffusely distended and contain markedly increased amounts of turbid, yellow-tinged, serous to mucoid ingesta.
Microscopic examination of sections of small intestine from all animals demonstrated villus length and architecture within normal limits (Fig. 2). Numerous gram-positive bacterial cocci (1 to 2 μm in diameter) were intimately associated with the brush border of epithelial cells along the proximal two-thirds of the intestinal villi (Fig. 3). These bacterial cocci were most often arranged in dense bands of 5 to 10 layers, but rarely formed a single layer (Fig. 4). There was no histologic evidence of associated inflammation or cellular degeneration in the superficial mucosal epithelium. No other significant lesions were observed in other tissues.
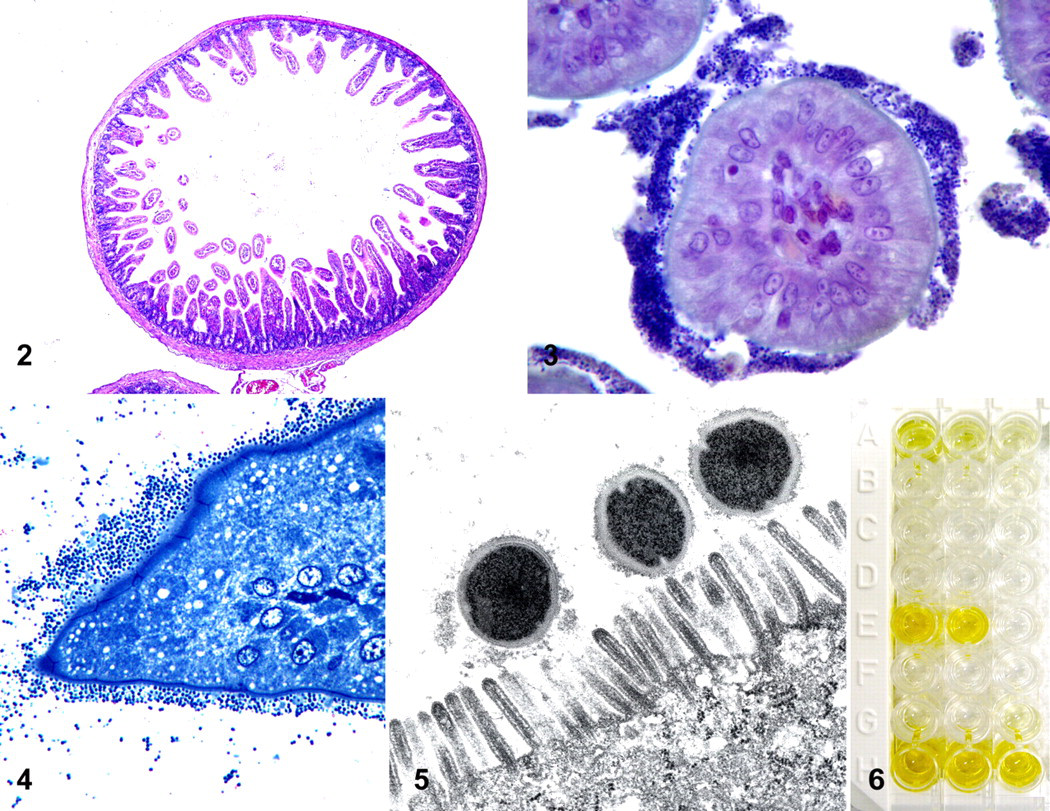
Transmission Electron Microscopy
On ultrastructural examination, bacterial cocci were intimately associated with the apical brush border of enterocytes and were surrounded by an irregular, indistinct, moderately electron-dense rim potentially representing a glycocalyx (Fig. 5). There were no significant alterations in the associated enterocytes. The architecture of the microvillus brush border was maintained; tight junctions were intact; and mitochondrial and nuclear membranes were smooth and well delineated.
Bacterial Culture
Bacterial culture of the small intestine from all litters yielded moderate growth of gram-positive cocci from each of the 4 batches of pooled intestines in addition to bacterial species considered to be part of normal intestinal flora (coliforms, nonhemolytic streptococci, and Corynebacterium spp). Gram-positive cocci were identified as Staphylococcus sp by colony morphology, Gram stain, and production of catalase. Staphylococci were β-hemolytic as determined by hemolysis characteristics on enriched sheep-blood agar and by the presence of coagulase enzyme. In addition, the isolated staphylococci were VP negative and failed to grow on P agar, distinguishing the organisms as belonging to the S intermedius group.
Polymerase Chain Reaction Fragments and Nucleic Acid Sequence Analysis
The nucleic acid sequence of the PCR amplicon from the 16S ribosomal RNA gene used for analysis was 718 base pairs (bp) in length and began at base 12 of the 16S ribosomal RNA gene. The amplified sequence was 99 to 100% similar to that of S intermedius, Staphylococcus pseudintermedius, S delphini, and Staphylococcus schleiferi. The nucleic acid sequence from the sodA gene was 313 bp in length and began at base 23 or 28 of the sodA gene, owing to variation of available sequences in GenBank. The sequence was 99% similar to that of S delphini, 97% similar to S pseudintermedius, 93% similar to S intermedius, and 85% similar to S schleiferi. The nucleic acid sequence of the hsp60 gene was 485 bp in length and began at base 35 of the hsp60 gene. The sequence was 95 to 97% similar to that of S delphini and 92% similar to S intermedius and S pseudintermedius. Taken together, the nucleic acid sequences from the 16S ribosomal RNA gene, the sodA gene, and the hsp60 gene indicated the bacterium isolated was S delphini. Amplification of fragments from the enterotoxin genes was successful for only the SEE gene. The amplified fragment was 302 bp in length and began about base 69 of the SEE gene of S aureus. The amplified sequence was 91% similar to the SEE gene sequence of S aureus and was 77 to 79% similar to other enterotoxin or enterotoxin-like genes from various Staphylococcus sp.
ELISA Testing for Enterotoxin
Significant levels of SEE were detected in extracts from batch Nos. 3113870 and 3113878-1 (Fig. 6). There was weak positivity for SEA in batch No. 3113870 (Fig. 6) and no evidence of enterotoxin production in batch No. 3113878-3.
Additional Diagnostic Testing
Diagnostic PCR assays were negative for rotavirus and coronavirus.
Discussion
The described clinical, gross, and histologic findings observed in this outbreak, in combination with the absence of overt enteritis or mucosal damage, are consistent with a secretory diarrhea, as would be expected with enterotoxin exposure. Furthermore, histopathologic examination of sections of intestine demonstrated colonization of intestinal villi by small gram-positive bacteria consistent with staphylococci. No evidence of any other infectious process was observed in this outbreak. Bacterial cultures failed to demonstrate pathogenic strains of E coli. PCR was negative for rotaviruses and coronaviruses. No evidence of an underlying viral infection was observed histologically or ultrastructurally. In light of the lack of other demonstrable infectious agents or alterations in the intestinal architecture, we postulate that the observed diarrhea in these mink kits was due to the colonization of the small intestine by enterotoxigenic strains of S delphini.
In this case series, bacteria that were isolated from the intestine of neonatal mink affected by clinical signs consistent with sticky kit syndrome were phenotypically consistent with the S intermedius group. PCR further speciated such isolates as S delphini and showed that the isolates possessed enterotoxin genes. Two of the three S delphini isolates in our series produced SEE, and one of these strains was weakly positive for SEA, as determined by ELISA. The weak signal elicited by the immunoassay for SEA in isolate No. 3113870 may simply indicate low levels of enterotoxin A expression. It is also possible that cross-reactivity of antienterotoxin A antibodies with enterotoxin E is present, given that these two enterotoxins exhibit 90% amino acid sequence homology. 7 Alternatively, it is possible that enterotoxin A is significantly expressed by this isolate but that polymorphisms within the antigenic regions mask its immunogenicity in the detection system used in this case.
Recently, reclassification of phenotypically identified S intermedius strains has been suggested on the basis of genomics. According to a report by Sasaki et al, speciation of staphylococci is often not possible on the basis of phenotype or commercially available typing kits alone. 16 Vulfson et al reported that coagulase-positive β-hemolytic staphylococci isolated from mink were almost exclusively S intermedius, but the evidence behind this conclusion was not provided. 19 One strain of phenotypcially identified S intermedius from a mink was reclassified as S delphini group A in the study by Sasaki et al. 16 Such findings make interpretation of previous reports of S intermedius difficult, given that isolated bacteria that were assumed to be S intermedius based on phenotypic characteristics may have been a different species, such as S delphini or S pseudointermedius.
Bacteria that are phenotypically consistent with the S intermedius group are relatively ubiquitous and occasionally isolated from the external surfaces of clinically normal animals. Such bacteria are generally seen as opportunistic pathogens and, as such, have been implicated in cases of dermatitis, cellulitis, otitis externa, arthritis, mastitis, and endocarditis in a variety of species. 19 In mink, bacteria that were phenotypically consistent with S intermedius have been isolated from cases of adenitis of the cervical apocrine glands of neonatal mink and pyometra, vaginitis, cystitis, pyelonephritis, pleuritis, peritonitis, and mastitis in adults. 19 In a necropsy survey of mink kits that died before weaning on a commercial mink farm in Canada, systemic bacterial infection was the most common cause (19%) of mortality in kits ranging in age from 4 days to weaning, with β-hemolytic staphylococci being one of the three most commonly isolated bacteria. 17 It is interesting that bacteria phenotypically consistent with S intermedius have been associated with adenitis of the cervical apocrine glands given that hypersecretion from these glands is reported in cases of sticky kit syndrome. 19 Cervical apocrine glands were not examined in this outbreak, so the presence or absence of Staphylococcus-associated adenitis cannot be confirmed.
In contrast to other species, bacteria that are phenotypically consistent with S intermedius have been identified as part of the gastrointestinal flora of clinically unremarkable kits. 19 One study found that fecal counts of β-hemolytic staphylococci peaked during the first week of life, subsequently declined, and were rare in adults. 19 In a study that investigated astrovirus as a potential cause of neonatal mink diarrhea, Englund et al reported that colonization of the intestinal villi by coccoid bacteria significantly increased the risk of preweaning diarrhea. 4 Such colonization was seen in 15 mink kits from farms affected by sticky kit syndrome and in 6 kits from unaffected farms. 4 Of the 15 kits from the affected farms, 11 had clinical signs, but 4 appeared clinically healthy. 4 Because only formalin-fixed tissue was available in their study, no bacteriologic testing was preformed. 4 Because of the isolation of S intermedius from clinically normal animals and the lack of associated histologically evident changes in the intestinal tract, the significance of intestinal colonization was unclear. 4 Pathogenicity of such bacteria may depend on acquiring the ability to produce enterotoxins, as found in the strains isolated from affected animals in our cases.
Staphylococcal enterotoxins are commonly incriminated in outbreaks of food poisoning in humans. 1,2 The mechanism by which vomiting and diarrhea are induced is poorly understood. Staphylococcal enterotoxins are most often viewed as superantigens, but this mechanism poorly describes the predominant acute emesis and enteric signs seen in cases of food poisioning. 3,12 Food-borne enterotoxicoses are most commonly attributed to SEA, but other staphylococcal enterotoxins can be involved in outbreaks. 12 Vomiting and diarrhea have been experimentally produced in adult mink through the ingestion of feed spiked with SEA, suggesting that mink are susceptible to staphylococcal enterotoxicosis. 8
S aureus is the species of staphylococci most commonly associated with enterotoxicosis. Strains of S intermedius that produce enterotoxins have been described; however, only a few outbreaks of food poisoning have been attributed to enterotoxins produced by this bacterium, 1,10 although this may be a reflection of a lack of testing rather than a lack of incidence. In contrast to that of S aureus, a systematic screening for enterotoxins was only recently performed at the genomic level for the coagulase-positive species S intermedius. 1 Such studies showed, on molecular and immunological levels, that 90% of animal isolates of S intermedius that possessed enterotoxin genes produced enterotoxins, highlighting a potential role in zoonotic enterotoxicosis. 1 Expression of enterotoxin genes and production of enterotoxins by S delphini have not been reported.
Interestingly, S aureus has been shown to colonize the intestine of humans and mice, which is consistent with our observations of S delphini colonizing the small intestine of mink in this outbreak. 9,11,13 There are reports of enteritis in humans following surgery owing to enterotoxin-producing, methicillin-resistant S aureus that colonized the small intestine. 11,18 Whether enterotoxin production by staphylococci that colonize the intestines is involved in the production of natural disease in species other than mink is unclear.
In summary, the present study describes an outbreak of neonatal diarrhea on an Idaho mink farm, consistent with the previously described sticky kit syndrome. Disease was associated with intimate surface colonization of affected preweaning mink kit small intestine by strains of S delphini that produced enterotoxin types A and E. Although direct causation cannot be confirmed, we postulate that the observed diarrhea in the mink kits was due to hypersecretion as a result of staphylococcal enterotoxicosis.
Footnotes
Acknowledgments
We thank Roger Maes and Annabel Wise of the Michigan State University Diagnostic Center for Population and Animal Health, Lansing, Michigan, for diagnostic testing of mink intestinal samples for coronavirus and rotavirus.
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.