
Abstract
For humans, strong evidence indicates that some mucosal papillomavirus (PV) types cause genital and oral neoplasia, and weaker evidence suggests that some cutaneous PVs may cause cutaneous squamous cell carcinomas (SCC). For nonhuman species, strong evidence supports a causal role of PVs in the development of feline and equine sarcoids. Likewise, PVs are believed to cause cutaneous SCCs in rabbits, western barred bandicoots, and some rodents. Furthermore, some evidence suggests that PVs may influence the development of both feline and canine cutaneous SCCs. This review discusses the evidence that PVs cause human cutaneous SCCs and the proposed mechanisms for this action. It then reviews preneoplastic and neoplastic skin diseases that are associated with PV infection in nonhuman mammals.
Papillomaviruses (PVs) used to be contained, along with the polyomaviruses, within the Papovaviridae; however, they are now recognized to be within their own family, the Papillomaviridae. 28 PVs are small circular double-stranded DNA viruses that generally infect epithelium. 28 Most PVs are highly host specific, and they have been detected in most species that have been extensively studied. 28,105 PVs are usually site specific, and lesions are often restricted to limited locations on the body. 105 Numerous PV types have been identified and are classified using the highly conserved L1 gene. 28 PV types are grouped into genera, with most of the more than 100 human PVs (HPVs) classified within the alpha or beta PVs. 28 The alpha PV genus include the oncogenic “high-risk” mucosal types, as well as PV types that cause benign mucosal or cutaneous lesions. 28 The beta PV genus contains cutaneous PVs that rarely cause visible lesions without immunosuppression. 28 Some evidence suggests that the hair follicle is a reservoir for many beta PV types. 15 Within the nonhuman mammals, the delta PV genus is important because its PVs cause benign fibropapillomas in ungulates. 28 Delta PVs have the unique ability to infect multiple species and cause equine sarcoids (Table 1 ). 28,79 Two members of the lambda PV genus have been associated with skin lesions in cats and dogs: Felis domesticus PV 1 (FdPV-1) and canine oral PV (COPV). 16,107,112 However, the other feline and canine PVs associated with cutaneous neoplasia—FdPV-2 and Canis familiaris PV (CfPV) 2, 3, and 4—have yet to be assigned to a named PV genus.
Summary of pre-neoplastic and neoplastic skin lesions associated with fully-sequenced papillomaviruses in non-human mammals. a

a PV, papillomavirus; FdPV, Felis domesticus papillomavirus; IHC, immunohistochemistry; PCR, polymerase chain reaction; SCC, squamous cell carcinoma; CfPV, Canis familiaris papillomarvirus; EM, electron microscopy; COPV, canine oral papillomavirus; BPV, bovine papilloma virus; ISH, in situ hybridization; CRPV, cottontail rabbit papillomavirus; BPCV, rousettus aegyptiacus papillomavirus type 1; RaPV,; MnPV, Mastomys natalensis papillomavirus; SB, Southern blot.
When considering infection of the epidermis by PVs, it is convenient to subdivide the keratinocytes into a population of self-renewing basal cells and a population of terminally differentiated keratinocytes. 3 Infection of basal cells by PVs can result in a persistent asymptomatic infection. 77 However, for viral replication to occur, infected basal cells have to become terminally differentiated. 77 Therefore, to maximize viral replication, PVs attempt to increase both proliferation of basal cells and terminal keratinocyte differentiation. 72 If successful, PVs stimulate epidermal proliferation that can result in the folding of the epidermis and the development of a viral papilloma. 28 The development of an immune response causes resolution of most viral papillomas, 40 and these will not be further discussed.
PVs can influence cell growth and differentiation; as such, they have the potential to induce neoplastic transformation of infected cells. Mucosal PVs are estimated to cause 5.2% of all human cancers, and PV infection is strongly associated with squamous cell carcinomas (SCCs) of the cervix, vagina, penis, anus, and oral cavity. 37,80,126 It remains uncertain whether cutaneous PVs cause human skin cancer. 2,7,62,126 This review discusses the evidence supporting a causal association between PVs and cutaneous neoplasia in people, along with the proposed mechanisms by which cutaneous PVs influence skin cancer development. It then reviews the cutaneous preneoplastic and neoplastic diseases that have been associated with PVs in nonhuman mammals.
Humans
The hypothesis that cutaneous PVs cause human skin cancer is supported by the high rates of cutaneous neoplasia seen in immunosuppressed people and by the epidemiological associations between PV infection and skin cancer identified in immunocompetent people. Immunosuppressed renal transplant recipients were found to have increased rates of cutaneous PV infection, 6 to frequently develop large numbers of viral warts, 14 and to have a markedly elevated risk of cutaneous SCC development. 99 Although it cannot be proven that the increased rate of cutaneous PV infection causes the increased cancer development, SCCs from immunosuppressed people more frequently contained PV DNA than do similar neoplasms from immunocompetent individuals. 41 The strongest predisposition to SCC owing to immunosuppression is observed in people with epidermodysplasia verruciformis (EV). 2,77 This inherited defect prevents an immune response against beta-PVs. 77 The genetic defect has been localized to the EVER1 and EVER2 genes that code for transmembrane channel–like (TMC) proteins 6 and 8, respectively. 50 Why TMC protein dysfunction increases beta-PV infection is currently uncertain, but all other immune functions are normal. 77 People with EV develop multiple cutaneous viral plaques during childhood that often progress to SCCs before 40 years of age. 2,77 The progression from viral plaque to SCC provides strong evidence of a significant role of PVs in SCC development in people with EV. However, most cutaneous SCCs in immunocompetent people do not develop as a progression of a viral plaque. 63,87 Therefore, although PVs appear to be an important cause of SCCs in people with EV, SCCs may develop by different mechanisms in immunocompetent people.
Epidemiological studies revealed that cutaneous SCCs in immunocompetent people are strongly associated with sun exposure. 33 However, not all differences in SCC rates between populations can be explained by differences in sun exposure, indicating that additional factors may influence SCC development. 29 Evidence that beta-PVs could be one of these additional factors includes the more frequent detection of PV DNA in cutaneous SCCs than in samples of nonneoplastic skin. 46,98 Additionally, immunocompetent people with cutaneous SCCs more frequently had antibodies against beta-PVs than did people without SCCs. 24,49 Furthermore, variations in the EVER2 gene were found to predispose to increased beta-PV infection and increased cutaneous SCC development in immunocompetent people. 81 However, PV DNA was amplified from 80% of nonneoplastic skin samples, 6 and antibodies against beta-PVs were detected in half of people without cutaneous neoplasia. 24 Because such a high proportion of people have cutaneous PV infections, it is difficult to differentiate between PVs causing neoplasm development and PVs being “innocent bystanders.” Although PV DNA has been detected within some nonepithelial human skin neoplasms, the significance of these findings remains uncertain. 4
When determining whether or not PVs cause neoplasia, it is important to identify mechanisms for an oncogenic action. Human cervical and oral SCCs are caused by infection with “high risk” mucosal alpha-PVs, predominantly HPV-16 and HPV-18. 37,96 The mechanisms for the oncogenic action of these mucosal PVs are well established. 124 The dominant oncoprotein is the product of the E7 gene, which strongly associates with and inactivates the retinoblastoma (RB) tumor suppressor protein, promoting cell cycling. 95,124 The E6 oncoprotein additionally promotes neoplastic transformation by activating telomerase, impairing cell cycle regulation, and inactivating the transformation-related protein 53 (TRP53, p53). 95,124
In contrast, human cutaneous SCCs are mainly associated with beta-PVs, especially HPV-5 and the species 2 beta-PVs. 36,42,117,126 Evidence that beta-PVs are oncogenic includes a high rate of cutaneous SCC development in beta-PV E6/E7 transgenic mice 31,89 and the increased proliferation and life span of cultured keratinocytes expressing beta-PV E6/E7 proteins. 18 However, unlike with the high-risk mucosal alpha-PVs, the molecular mechanisms by which beta-PVs cause neoplastic transformation are less clear.
One possible mechanism is by prevention of apoptosis by beta-PV E6 protein-mediated degradation of BCL2 homologous antagonist/killer (BAK). 47 Additionally, beta-PV E6 proteins have been found to activate telomerase, prolonging cell life. 8 Furthermore, although beta-PV E6 proteins cannot degrade TRP53, they can inhibit TRP53 function. 1 In contrast to the mucosal alpha-PVs, beta-PV E7 proteins appear weakly oncogenic and are unable to transform cultured cells that do not have preexisting RAS mutations. 121 However, some beta-PVs produce E7 proteins that inhibit RB protein function. 18,90 Other actions of the beta-PVs that may promote neoplastic transformation include impairment of the host immune response by disrupting the interferon pathway 27 and dysregulation of cell survival, differentiation, and growth factor responsiveness by altered serine/threonine kinase (AKT) activity. 78
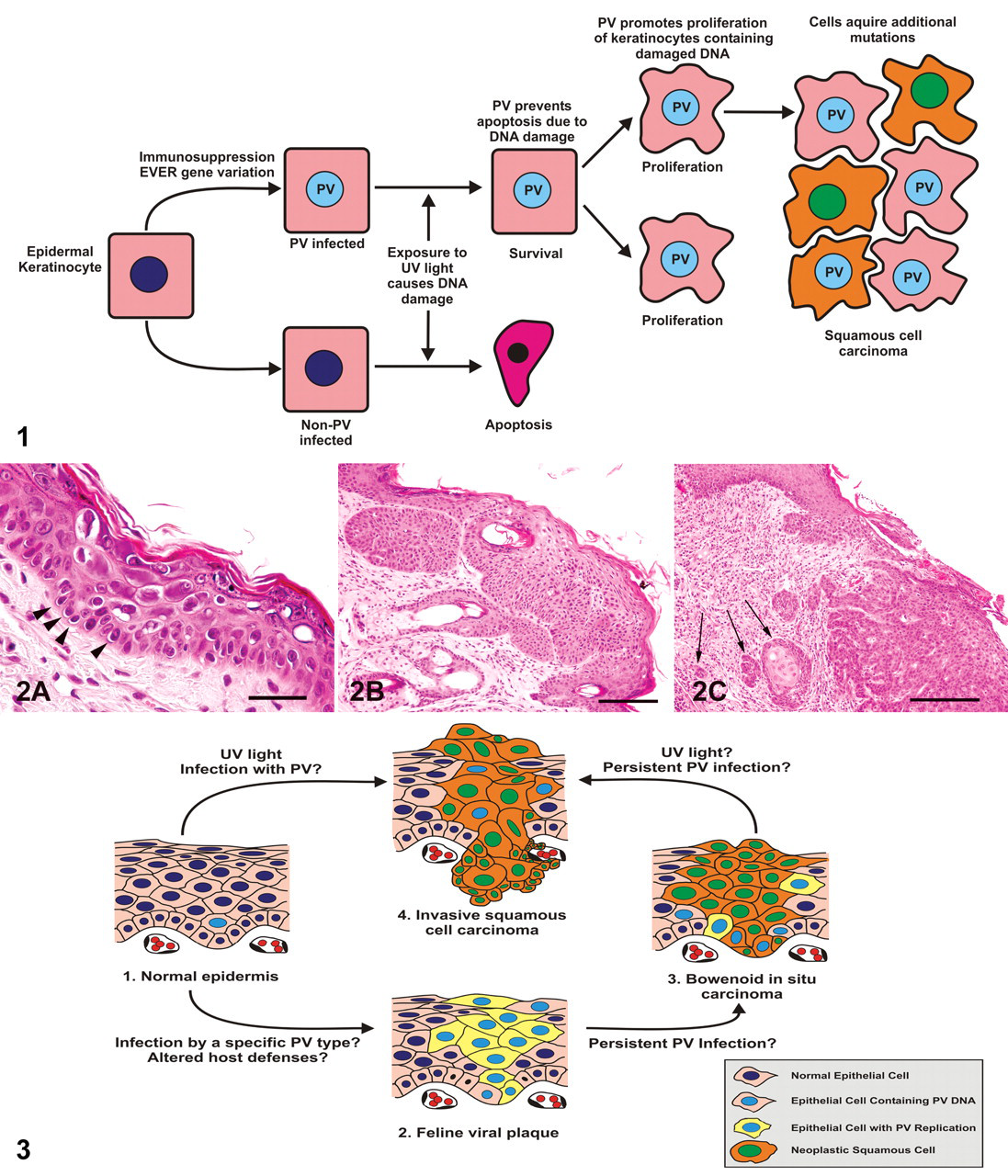
Ultraviolet (UV) light causes neoplasia by damaging DNA, increasing the chance of an oncogenic mutation. 52 In a normal cell, this DNA damage activates the TRP53 protein, resulting in either DNA repair or cell apoptosis. 52 The hypothesis is that in a beta-PV-infected cell, the inhibition of TRP53 protein function prevents repair of the UV damaged DNA and apoptosis. 1,101 Apoptosis is further inhibited by the PV-induced degradation of BAK. 47 The ability of some beta-PVs to promote epithelial proliferation may further promote carcinogenesis by increasing replication of epithelial cells that contain UV-damaged DNA. 72 Therefore, beta-PVs and UV light may act as cocarcinogens in the development of cutaneous SCCs in immunocompetent people (Fig. 1 ). 2
Cats
As in humans, PV infections in cats can be asymptomatic. 67,71 However, in some cats, cutaneous PV infection results in the formation of a feline viral plaque. Feline viral plaques can progress to Bowenoid in situ carcinomas (BISCs), which can progress to invasive SCCs (ISCCs), 40,118 and all three lesions can be present on the same cat (Fig. 2). Figure 3 presents a proposed pathogenesis of epithelial lesions associated with PVs in cats.
Feline viral plaques typically appear as multiple, scaly, flat, variably pigmented lesions that can develop anywhere on the body. 40 Feline viral plaques have been observed in 2 cats receiving long-term immunosuppressive therapy and in 2 cats that were seropositive for feline immunodeficiency virus (FIV). 21,32,70 Although this suggests that immunosuppression may predispose to plaque formation, plaques have been reported in cats without any identified immunosuppressive disease. 58,118 Histologically, they appear as well-demarcated foci of epidermal hyperplasia and hyperkeratosis. 40 Keratinocytes within the plaques often have PV cytopathic changes, such as nuclear shrinkage, perinuclear clearing (koilocytosis), and increased quantities of blue-grey foamy cytoplasm. 21,32 The histological appearance of plaques containing prominent PV cytopathic changes is similar to that of the viral plaques seen in people with EV. 64 PV antigen was detected in a high proportion of feline viral plaques using immunohistochemistry (IHC). 118 IHC detects PV L1 protein, which is produced in only the late stages of viral replication. 57 Because replication of PVs is restricted to terminally differentiated keratinocytes, 77 PV L1 protein is present in only the superficial layers of the epidermis overlying the viral plaque, especially the stratum granulosum. 70 With polymerase chain reaction (PCR), PV DNA was amplified from a feline viral plaque. This lambda-PV was closely related to COPV and designated FdPV-1. 28,107,112 However, DNA sequences from FdPV-1, FdPV-2, and a novel PV were all amplified from a swab of a feline viral plaque. 70 All 3 sequences were also detected in a swab of nonlesional skin from the same cat; as such, the PV that causes feline viral plaques remains uncertain. 70
Feline BISCs typically appear as crusting, hyperpigmented, roughly circular lesions. 34 BISCs can develop on haired, pigmented skin suggesting that, unlike with ISCCs, UV exposure is not a significant cause of lesion development. 34,94 Immunosuppression may predispose to BISCs; however, as with viral plaques, cats can develop BISCs without identifiable immunosuppressive disease. 40,67 Histologically, BISCs are characterized by thickening and dysplasia of the epidermis with loss of keratinocyte nuclear polarity. 34,40 BISCs are suggested to represent neoplastic transformation of a feline viral plaque. 118 Cytopathic evidence of PV infection can be present in early lesions but is usually absent in more mature neoplasms. 34,68 PV antigen was detected in 11 118 and 48% 34,55 of BISCs using IHC. PV DNA that was amplified from a BISC using PCR was designated FdPV-2. 54 With consensus PCR primers, short segments of PV DNA have been amplified from 5 of 21 76 and 11 of 18 BISCs. 68 Sequencing of 6 amplicons in the latter study revealed that 5 BISCs contained FdPV-2, whereas the other contained a PV that had previously been amplified from a swab of human skin. 68 The use of primers designed to amplify FdPV-2 detected PV DNA within 20 of 20 BISCs. 67
ISCCs are the most common malignant cutaneous neoplasm of cats. 40 Although a small proportion may develop as a progression of a feline viral plaque or BISC, the majority of feline ISCCs develop within areas of actinic keratosis. 40 Owing to the strong association between ISCCs and sun exposure, ISCCs are most common in lightly pigmented, poorly haired areas, such as the eyelids, nose, and pinnae. 40 Although high rates of feline SCC development were reported in cats infected with FIV, 45 most cats that develop cutaneous ISCCs are not immunosuppressed. Histologically, ISCCs appear as nests and trabeculae of poorly differentiated epithelial cells extending through the basement membrane into the dermis. 40 Neither cytopathic evidence of PV infection nor PV antigen detection has been reported within feline ISCCs. 102 This suggests that little PV replication is possible within the dysplastic keratinocytes. An inverse relationship between epithelial cell dysplasia and PV replication is also seen as human cervical preneoplastic lesions progress to ISCC. 35 PCR with consensus primers detected PV DNA in 4 of 22 ISCCs. 76 PCR using specific primers amplified segments of FdPV-2 DNA from 17 of 20 ISCCs. 67 One feline ISCC was found to contain DNA segments from two different PVs. 66
The detection of FdPV-2 DNA in 20 of 20 BISCs and 17 of 20 ISCCs confirms an association between PVs and cutaneous squamous neoplasia in cats. 67 However, this does not prove that PVs cause feline skin neoplasia. It is possible that the association between cutaneous SCCs and PVs is due to the cutaneous neoplasms providing a more permissive environment that allows greater PV persistence or increased viral replication. Therefore, PV DNA may be present within the neoplastic cells without having influenced neoplastic transformation. With specific primers, FdPV-2 DNA is detectable in half of normal feline skin swabs. 71 Therefore, many cats infected by FdPV-2 do not develop cutaneous neoplasia. As with humans, it is currently uncertain whether feline cutaneous SCC development is significantly influenced by PV infection.
In addition to their association with epithelial tumors, PVs are associated with feline sarcoids. 91,111 These are uncommon dermal neoplasms that tend to occur on the head, neck, or digits of younger cats from rural areas. 40 These neoplasms are also known as feline cutaneous fibropapillomas and histologically consist of proliferating fibroblasts covered by hyperplastic epidermis. 40 Neither histological nor immunohistochemical evidence of PV infection has been reported, suggesting that little viral replication is present within the neoplasms. 91 However, PV DNA was amplified from a high proportion of feline sarcoids, 69,91,111 and PV DNA has been localized to proliferating fibroblasts using in situ hybridization (ISH). 111 Short sections of the feline sarcoid–associated PV (FeSarPV) L1 gene were 72% similar to BPV-1 and ovine PV 2 and 71% similar to BPV-2. 69 Although definitive classification of FeSarPV is not possible without the complete L1 gene sequence, 28 the virus appears likely to be a novel type of delta-PV, which suggests that, like equine sarcoids, feline sarcoids may be due to cross-species infection. Cross-species infection is further supported by the failure to detect FeSarPV DNA within 120 nonsarcoid feline samples. 69 However, because this PV has been detected in only feline sarcoids, the reservoir host of FeSarPV is unknown. Feline sarcoids are much less common than equine sarcoids. 38,40 If feline sarcoids are caused by cross-species infection, this could indicate that cats are infrequently exposed to the reservoir host. However, the comparative rarity of feline sarcoids could also be due to many other viral or host factors. Interestingly, similar PV sequences were amplified from a sarcoid on a mountain lion, 92 suggesting that the causative PV is partially adapted to felids. Both names used to describe these lesions—feline sarcoid and feline cutaneous fibropapilloma—are poorly descriptive of their pathogenesis and behavior. However, because feline sarcoids have not been reported to metastasize and may yet be proven to represent a hyperplastic process rather than a true neoplasm, an alternative name such as papillomaviral-associated cutaneous fibrosarcoma is also considered poorly descriptive. As understanding of the pathogenesis of the virally induced cutaneous neoplasms increases, names more appropriate than BISC 105 and sarcoid will likely be developed.
Dogs
As in humans and cats, cutaneous PV infections in dogs can induce viral plaques that may subsequently progress to in situ or invasive carcinomas. Canine pigmented viral plaques typically present as multiple lesions (< 3 cm in diameter) that are most common on the ventrum and limbs. 115 Pugs and Miniature Schnauzers have been reported to be predisposed to plaque formation, 73,115 whereas plaques in other breeds have generally been restricted to immunosuppressed dogs. 19,100 Although neoplastic transformation is rare, progression of a viral plaque to an in situ carcinoma has been reported in 3 dogs, 19,73,114 with a single report of progression to ISCC. 100 Histological examination of a canine viral plaque reveals papillary thickening of the epidermis, hyperkeratosis, hypermelanosis throughout the epidermis and superficial dermis, and the presence of large keratohyaline granules. 73 Histological evidence of viral infection is often subtle. 73 However, PV antigen has been demonstrated within plaques, 19,73,100 and PV particles have been detected in keratinocyte nuclei using electron microscopy. 73,100 PV DNA was amplified from a Rhodesian ridgeback with viral plaques and in situ carcinomas and was designated CfPV-3. 114 The closely related CfPV-4 has been consistently amplified from viral plaques on pugs. 74,109,115 Serology revealed that 26.9% of South African dogs and 1.3% of Swiss dogs had antibodies against CfPV-3. 53 This suggests that, as with humans and cats, asymptomatic PV infection of canine skin is common, but PVs rarely cause visible lesions.
PVs have also been associated with canine endophytic papillomas. These rare masses develop more frequently on the footpad and ventrum of younger dogs and histologically appear as well-circumscribed flasklike proliferations of hyperplastic epidermis extending into the dermis. 20 They commonly contain PV antigen 20 and PV DNA, subsequently designated CfPV-2, as amplified by PCR from a footpad endophytic papilloma. 122 Neoplastic transformation of endophytic papillomas was reported in bone marrow–transplanted severe combined immunodeficiency (SCID) dogs. 39 Out of 24 dogs, 17 developed endophytic papillomas containing CfPV-2 antigen. 39 Of these dogs, 11 were euthanatized owing to persistent or widespread papillomas, whereas 4 dogs had papillomas progress to ISCCs within 4 years. 39 The predicted CfPV-2 E6 protein has some similar binding sites to the HPV-16 E6 protein, which may explain the putative oncogenic activity of this virus. 122
Infection by COPV typically causes transient papillomas within the oral mucosa. 16 Like FdPV-1, COPV is classified as a lambda-PV. 28 Although COPV can infect the skin of immunosuppressed dogs, 104 natural infection by COPV has not been associated with canine cutaneous neoplasia. In contrast, administration of a live COPV vaccine to 4,500 dogs resulted in cutaneous ISCCs at the injection sites of 9 dogs. 16 PV antigen was detected using IHC within 5 of the ISCCs. 16
Whereas canine pigmented viral plaques and endophytic papillomas can undergo malignant transformation, most canine ISCCs do not develop from these precursor lesions. 40 However, investigation using IHC detected PV L1 antigen in 5 of 9 102 and 10 of 100 93 routine diagnostic ISCC samples. The immunohistochemical evidence of PV replication is surprising because PV cytopathic changes have not been reported within canine ISCCs. The presence of PVs within the neoplasms could not be confirmed using electron microscopy. 93 With consensus PCR primers, PV DNA segments were amplified from 2 of 2 in situ carcinomas and 2 of 17 ISCCs. 125 No neoplasm contained segments from the same PV, with 1 that was most similar to BPV-1 and with 3 that were most similar to HPVs. 125
Horses
There is less evidence that PVs cause cutaneous epithelial neoplasia in horses. PV L1 antigen was not detected using IHC in a series of 80 equine cutaneous, genital, and ocular ISCCs. 48 However, PV antigen was present within a vulval equine papilloma that had evidence of neoplastic transformation to a SCC in situ. 97 The contribution of the PV to the neoplastic transformation of the papilloma is uncertain. 97 PV DNA was not amplified by PCR from 6 equine ISCCs. 22,26
The association between equine sarcoids and PVs has been extensively studied and is the subject of 2 recent reviews. 25,75 Sarcoids are the most common neoplasm of horses and have been reported in donkeys, zebra, and tapir. 38,51,56,59 Horses less than 4 years old most frequently develop sarcoids, and areas predisposed to trauma, such as the head, legs, and ventral trunk, are most commonly affected. 38 Histologically, equine sarcoids appear as proliferations of fibroblasts covered by hyperplastic epithelium. 38 Equine sarcoids do not contain histological or IHC evidence of viral infection and are considered nonproductive. 5,75,83 However, BPV-1 or BPV-2 DNA was amplified by PCR from most equine sarcoids. 22,79 With ISH, PV DNA has been localized within the proliferating fibroblasts, 110 and PV proteins have been detected within the neoplasms. 23
Although BPV-1 and BPV-2 are the accepted cause of equine sarcoids, the epidemiology of the viral infection remains uncertain. Initial studies detected BPV in bovine fibropapillomas and equine sarcoids but not within any nonsarcoid equine sample, 22,26,79 which suggested that equine sarcoids were the result of cross-species infection of horses by a PV from cows. 22,26,79 However, BPV-1 or BPV-2 DNA was recently detected in 57% of skin samples from clinically normal horses 10 and in samples of equine dermatitis. 26,123 Analysis of the variability within BPV sequences from equine sarcoids and bovine papillomas suggested that “equine-adapted” BPV strains exist. 75 If horses are commonly asymptomatically infected with “equine-adapted” BPVs, 10 entry of the virus into the dermis (due to trauma, eg) may be more important in sarcoid development than exposure to the PV. In addition, because some major histocompatibility complex class II haplotypes predispose to equine sarcoid development, 61 the immune response to the PV appears to be important in sarcoid development.
BPV-1 and BPV-2 are both delta-PVs. 28 These PVs have the unique ability to infect multiple species, causing self-resolving fibropapillomas in one species and fibroblastic neoplasia in others. 28 In addition to causing equine sarcoids and being the likely cause of feline sarcoids, delta-PVs have been reported to cause fibrosarcomas when injected into mice and hamsters. 11,85 Although the mechanisms by which delta-PVs transform mesenchymal cells have not been fully resolved, BPV E5 oncoprotein is able to bind to and activate platelet-derived growth factor β receptor (PDGFβ-r). 12,30 In cultured fibroblasts, PDGFβ-r activation caused numerous downstream effects that promoted fibroblast growth and loss of cellular contact inhibition. 82 Studies of equine sarcoids likewise demonstrated PDGFβ-r activation and increased expression of the phosphoinositide 3-kinase-AKT-cyclin D3 pathway and c-Jun N-terminal kinase. 12 Evidence suggests that coexpression of BPV E5 and E7 is important in neoplastic transformation. 13
Others
The ability of cottontail rabbit PV (CRPV) to cause skin cancer was first recognized in 1935. 88 Infection of cottontail rabbits results in the formation of multiple papillomas that progress to ISCCs in around 25% of rabbits. 106 A higher proportion of papillomas progress to ISCCs when domestic rabbits are experimentally infected with CRPV 106 and neoplasm formation is accelerated by the application of carcinogens to the papillomas. 86 Like HPV-16 E7 protein, CRPV E7 protein inhibits Rb protein function. 43 Rabbits infected with CRPV were used as an animal model for the development of HPV vaccines. 17
Cutaneous papillomatosis and carcinomatosis is a significant cause of mortality in western barred bandicoots, an endangered species of marsupial. 119 This disease presents with multifocal papillomas, in situ carcinomas, and ISCCs that often involve the digits and mucocutaneous junctions of the lips. 119 Although cytopathic and IHC evidence of PV infection was restricted to papillomas and in situ carcinomas, 119 PV DNA was detected with ISH in 81% of the papillomas, 70% of the in situ carcinoma, and 29% of the ISCCs. 9 DNA sequencing revealed the causative virus, BPCV1 (rousettus aegyptiacus papillomavirus type 1), which has features of PVs and polyomaviruses, possibly suggesting a novel virus family. 120 A captive Egyptian fruit bat developed multiple papillomas and basosquamous carcinomas. 60 PV antigen was demonstrated by IHC within the lesions, 60 and a novel PV was subsequently amplified and designated RaPV-1. 84 Captive and wild Natal multimammate mice (Mastomys natalensis) develop keratoacanthomas and ISCCs in association with PV infection. 65,108 The PV associated with these lesions was designated MnPV (Mastomys natalensis papillomavirus), and transgenic mice expressing MnPV genes develop high rates of cutaneous ISCCs. 44 Seven trichoepitheliomas and 3 sebaceous carcinomas were observed in a colony of European harvest mice (Micromys minutus). 103 With Southern blot hybridization, PVs were detected in all trichoepitheliomas and in the single examined sebaceous carcinoma. 103 DNA was sequenced and the virus designated MmPV. 116 In sheep, PVs were detected with DNA hybridization within 15 of 16 samples of perianal SCC and 1 of 3 samples of normal skin. 113
Conclusion
There is strong evidence that immunosuppressed people are predisposed to PV-induced flat warts and plaques. 2 Cats and dogs, especially immunosuppressed animals, can develop histologically similar lesions. 39,40 Evidence of PV replication is often visible in feline and canine plaques, thereby supporting a viral etiology of these lesions. 100,118 Asymptomatic epidermal infection by the PVs that cause cutaneous viral plaques in humans, cats, and dogs appears to be common. 53,71,77 Therefore, the development of viral plaques is likely due to altered host defenses allowing increased viral replication. 77 Feline and canine viral plaques can both undergo neoplastic transformation, 100,118 although it is not known if the PVs within the viral plaques influence neoplasm formation. However, the frequent and rapid progression of endophytic papillomas to ISCCs observed in SCID dogs suggests that CfPV-2 infection can cause neoplastic transformation. 39
Most ISCCs in humans, cats, and dogs develop in immunocompetent animals within areas of actinic keratosis. 40,63 Evidence that PVs increase the development and progression of these lesions is less conclusive. Because ISCCs are associated with exposure to UV light, it is possible that PVs act as cofactors by preventing apoptosis of cells that contain UV light–damaged DNA. 2 However, although PV DNA has been detected within cutaneous ISCCs of humans, 36 cats, 67 and dogs, 125 the common presence of PVs in normal skin makes it difficult to prove that PVs influence neoplasia formation and are not “innocent bystanders.”
Feline and equine sarcoids are strongly associated with PVs. 75,111 However, in both cases, the epidemiology of the causative PV remains uncertain. 75,111 Feline and equine sarcoids could be the result of cross-species PV infections. However, some evidence suggests that equine sarcoids could be caused by “equine-adapted” strains of BPV. 10,75 Cats do not appear to be commonly asymptomatically infected by FeSarPV 69 ; however, the source of the causal PV is unknown.
Footnotes
The authors declared that they had no conflicts of interests with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.