
Abstract
A 9-month-old p53-null female mouse was found dead in its cage. At necropsy, a large thymic mass encompassed the heart. Microscopically, the mass was composed of numerous varying-sized cysts lined with simple squamous epithelial cells to columnar ciliated cells. Also present within this mass was a large aggregate of loosely arranged fusiform-shaped cells. These cells also were found in smaller numbers in the connective tissue surrounding the cysts. The larger aggregate of fusiform cells was positive for desmin and S-100 and negative for smooth muscle actin. Electron microscopy revealed well-formed Z lines and I bands of skeletal muscle phenotype. A diagnosis of rhabdomyoma within a congenital multilocular thymic cyst was made. The thymus contains a small population of myoid cells, which should be taken in consideration when evaluating thymic tumors.
The thymus in the mouse is similar to that of other mammalian species in respect to anatomy, cell populations, and involutionary changes. In mice, the thymus undergoes involutionary changes at around 35 to 80 days of age, 31 with the ability to repopulate. 32 Cystic cavities within the thymus are usually seen as incidental findings in adult mice. These cysts are usually small, isolated, and within the corticomedullary junction. 14 Besides containing lymphoid cells and epithelial components (Hassall corpuscles), the thymus contains few to rare numbers of myoid cells. 23 This article describes a multilocular thymic cyst, an uncommon finding in mice, associated with a myoid cell rhabdomyoma.
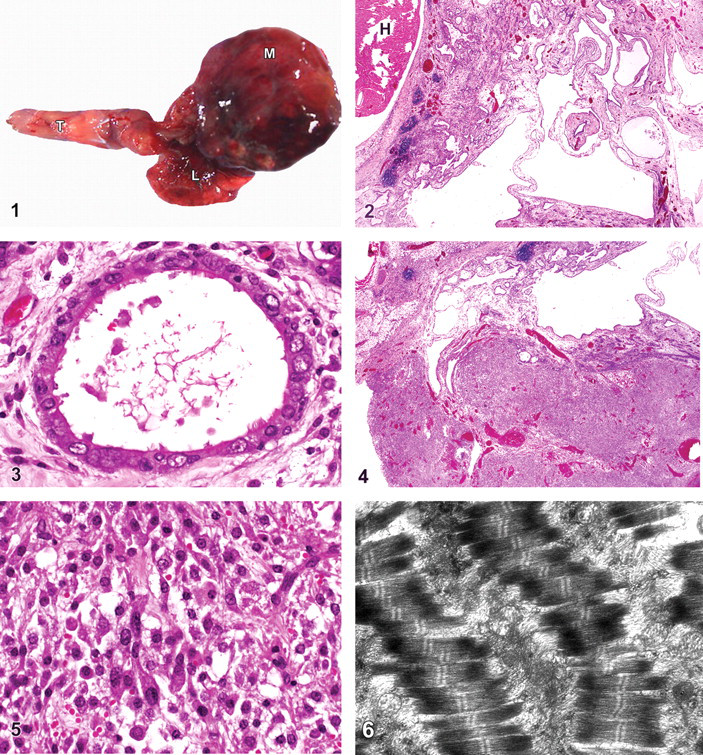
A 9-month-old female black p53-null mouse (B6.129S2-Trp53tm1Tyj/J) was housed at the National Institutes of Health (NIH; Bethesda, MD). The care of the animal followed the NIH’s Guide to the Care and Use of Laboratory Animals. The mouse was found dead during daily observations by the animal caretakers. The animal was submitted for diagnostic necropsy to the Diagnostic and Research Services Branch of the Division of Veterinary Resources at the NIH. On examination, the animal was obese with mild alopecia of the muzzle. Upon opening of the pleural cavity, a multilobulated reddish-purple mass was found (approximately 2 cm in diameter), which encompassed the heart and compressed the lungs (Fig. 1). On cut surface of the mass, serosanguineous fluid exuded from what appeared to be cystic structures. No other lesions were observed in the remainder of the animal.
The entire mass with associated heart, lungs, trachea, and tongue was fixed in 10% buffered formalin, processed, and embedded in paraffin; 5-μm sections were then cut and stained with hematoxylin and eosin (HE), Masson trichrome, and periodic acid–Schiff (PAS).
Immunohistochemistry for desmin, S-100, smooth muscle actin, and F4/80 (mouse macrophage) was performed on the mass. Briefly, 5-μm sections were deparaffinized in xylene, placed in absolute alcohol, and processed in a Dako Autostainer Universal Staining System (Dako, Carpinteria, CA). Endogenous peroxidase was inhibited by 5-minute incubation in 3% hydrogen peroxide. Antigen retrieval (desmin, S-100) was accomplished by pressure cooker heating for 20 minutes at 37°C in citrate buffer or with proteinase K for 5 minutes (F4/80) at room temperature. Sections were blocked with a commercially available protein block (Dako) for 5 minutes at room temperature. The primary antibodies were rabbit anti-desmin (1:3000; Thermo-Scientific, Fremont, CA), rabbit anti-S-100 (1:10,000; Dako), mouse anti–smooth muscle actin (1:100; Biocare Medical, Concord, CA), and rat anti-mouse F4/80 (1:400; AbD Serotec, Raleigh, NC). The detection system utilized a biotinylated goat–antirat antibody with streptavidin–horseradish peroxidase label for F4/80, polymerized horseradish peroxidase antirabbit secondary for desmin and S-100, and a mouse on mouse kit for smooth muscle actin. DAB chromogen (Dako) was added, and the slides were counterstained with Mayer hematoxylin. Positive control slides consisted of known positive mammalian tissue incubated with the relevant monoclonal antibody. Negative control slides consisted of the test tissue incubated with an isotype matched immunoglobulin.
An area of interest within the mass was trimmed from the paraffin block and processed for electron microscopy. The tissue was washed 3 times in xylene for 1 hour each and placed into 10% OsO4 in xylene overnight. Subsequently, the tissue was washed 3 times in xylene for 30 minutes each, changed to 2:1 propylene oxide:Epon and then 1:2 propylene oxide:Epon, and embedded in EMBed 812 resin (Electron Microscopy Sciences, Hatfield, PA). Thin sections, approximately 80 nm, were obtained by utilizing the Leica Ultracut-UCT Ultramicrotome (Leica, Deerfield, IL), placed onto 300 mesh copper grids, and stained with saturated uranyl acetate in 50% methanol and then with lead citrate. The grids were viewed in the JEM-1200EXII electron microscope (JEOL Ltd., Tokyo, Japan) at 80 kV, and images were recorded on the XR611M midmounted 10.5-Mpixel CCD (charge-coupled device) camera (Advanced Microscopy Techniques Corp., Danvers, MA).
Histopathology of the mass revealed numerous varying-sized cystic structures (Fig. 2). The largest cysts were primarily lined by a simple squamous epithelium with occasional plump ciliated epithelial cells. Most of the smaller cysts had simple cuboidal to columnar, most ciliated, epithelial cells (Fig. 3), whereas other cysts had stratified squamous epithelial cells. PAS-positive goblet cells were admixed with the ciliated epithelial cells. The lumina of many of these cysts were partially filled with a wispy fibrillar material admixed with PAS-positive proteinaceous fluid, plump macrophages, lymphocytes, and neutrophils. Multifocal to coalescing cords and islands of squamous epithelial cells (occasionally admixed with a mild to moderate neutrophilic and lymphocytic infiltrate) or forming small incomplete lumina were observed. Many of these lumina contain necrotic debris admixed with a few neutrophils and/or PAS-positive proteinaceous fluid. All cysts were within a mild to moderately edematous fibrous connective tissue matrix as demonstrated on Masson trichrome stain. Thymic lymphocytic remnants were observed at the periphery of the cysts.
Within an extension of the connective tissue that contained the cysts, and near the base of the heart, was a large circumscribed, nonencapsulated mass of mild to moderately anisocytotic and anisokaryotic round to fusiform cells admixed with large areas of necrosis and hemorrhage (Fig. 4). The cells contained round centrally located nuclei, with finely stippled chromatin and fibrillar eosinophilic cytoplasm. There were a few foci where cells had a spider web–like appearance primarily due to contraction of cells. Small clusters of cells were aligned haphazardly with each other (Fig. 5). Similar cells were also present in smaller numbers within the connective tissue containing the cysts. To establish if these cells were myoid in origin, immunohistochemistry was performed for desmin, S-100, smooth muscle actin, and F4/80 (macrophage). Results of the immunohistochemistry revealed a desmin-positive, S-100-positive, smooth muscle actin–negative, F4/80-negative population of cells that were identified as immature striated skeletal myoid cells. To confirm the striated muscle phenotype, electron microscopy of the mass was performed. Within these cells were multiple parallel myofibrils, each containing a single I band and Z line with A bands on either side (Fig. 6). Given the large population of myoid cells along with the numerous varying-sized cysts, a diagnosis of rhabdomyoma within a multilocular thymic cyst was made.
The thymus is a dynamic organ that involutes over time. The primary cell components consist of lymphoid cells and epithelial cells (Hassall corpuscles). Another cell type is the myoid cell, which occurs in small numbers in humans and mice but is quite common in reptilian and avian species. 1,10,23,27 Myoid cells are often isolated to the medulla. It is believed that they have an embryological origin from the neural crest and hence are S-100 positive. 24 Ultrastructurally, myoid cell myofibrils are parallel bundles; however, they are sometimes irregularly arranged, forming angles in various orientations within the cytoplasm, a consistent finding for an immature myoid cell. 23 These cells have been described in healthy humans during fetal and perinatal periods. Their exact role within the thymus is not known. The thymus contains significant autoantigens, and the presence of myoid cells may be necessary for the induction of tolerance in lymphocytes. Myoid cells may also have a role as myogenic precursors or stem cells in muscle. 33 Myoid cells play a role in sensitization and subsequent formation of antimuscle antibodies present in humans with myasthenia gravis. 15 Proliferation of myoid cells within the thymus contributing to tumor formation has been documented in humans. 11,22 Myoid cells have also been observed in, and have contributed to, human diseases such as thymomas. 7,20 A few case reports of rhabdomyomatous and rhabdomyosarcomatous neoplasms of the thymus, as well as thymic carcinomas with skeletal muscle differentiation, have been described in humans. 6,9,25,30 Only one case report of a rhabdomyomatous multilocular thymic cyst in human has been documented. 2
Thymic cysts have been documented in multiple animal species (mice, cow, sheep, dog, cat, dolphin, chicken, human) 4,14,17 and may be of diverse pathogenesis (congenital, infective, inflammatory, neoplastic). 29 Single small cysts are a frequent finding in the thymus and are thought to be congenital and to arise from the endodermal vestiges of third and fourth fetal branchial arches. Cysts can be lined by squamous, cuboidal, or columnar epithelial cells. Cysts can be lined by cilia and associated with goblet cells, similar in appearance to respiratory epithelium, which also arises from the branchial arches. 14 Small numbers of thymic cysts, primarily in the corticomedullary junction, are common in mice and are vital structures that may be engaged in active transportation of lymphocytes and secretions into the circulation. 14 Numerous thymic cysts in mice are uncommon for most strains, except for nude mice, whose bulk of the thymus is composed of small cysts. 3 In humans, multilocular cysts are thought to be an acquired reactive process due to some inflammatory condition that leads to cystic formation. 29 Although this mouse did not have much in the way of inflammation, the term multilocular cyst was used to identify the numerous varying-sized and lined cysts that were present.
The cysts were most likely congenital in origin and became enlarged by the presence of the mass of myoid cells. In humans, multilocular thymic cysts have been associated with thymoma, 21 carcinoma, 19 follicular hyperplasia, 12 and pseudoepitheliomatous hyperplasia. 28
Although the cysts presented in this case are presumed to have arisen from the thymus, one cannot rule out other origins. Cysts arising from the mediastinum are rare in humans and animals. Pericardial and bronchogenic cysts are the most commonly described cysts in humans. 16,26 In the veterinary literature, 1 dog had a bronchogenic cyst, 5 9 cats had a solitary mediastinal cyst of unknown origin (histology was not performed), 34 and 2 cats had cysts of thymic origin. 18
The p53 gene regulates the cell cycle. Alterations in this gene (mutations and loss) can lead to formation of numerous types of neoplasms, including lymphosarcoma, osteosarcoma, hemangiosarcoma, and rhabdomyosarcoma. 8,13 The most common thymic tumor in p53-null mice is lymphoma. The myoid cell rhabdomyoma in the thymus of this p53-null mouse may have the same fundamental growth pattern as that of rhabdomyosarcoma. Loss of the p53 gene may have allowed uncontrollable growth of myoid cells in the thymus.
We believe this to be the first report of an animal with a rhabdomyoma composed of myoid cells within a multilocular thymic cyst. Myoid cells are a normal yet rare cell type found in the thymus of animals and humans and should be considered a possible cell type in the formation of tumors within the thymus.
Footnotes
Acknowledgements
We would like to thank Jorge Chavez and Annie Merriweather for technical assistance. This work was supported in part by the Intramural Research Program of the National Institutes of Health, Office of Research Services (MFS), and in part by the Eunice Kennedy Shriver National Institute of Child Health and Human Development, National Institutes of Health intramural project Z01-HD-000642-04 (CAS).
The authors declared that they had no conflicts of interests with respect to their authorship or the publication of this article.
The authors declared that they received no financial support for their research and/or authorship of this article.