
Abstract
Objective
To investigate the effect of the casein kinase 2 interacting protein 1 (CKIP-1) on the apoptosis of the intestinal type of gastric cancer (GC).
Methods
The levels of CKIP-1 protein and the rates of apoptosis were measured in tissue samples of the intestinal type of GC and human GC cell lines. The rate of apoptosis and the protein levels of B cell lymphoma-2 (Bcl-2), Bcl-2 associated X protein (Bax), cleaved cysteinyl aspartate specific protease 3 (cleaved caspase-3), cleaved caspase-9, rat sarcoma (Ras), extracellular signal-regulated kinase 1 and 2 (ERK1/2) and phosphorylated extracellular signal-regulated kinase 1 and 2 (p-ERK1/2) were analysed in SGC7901 cells expressing CKIP-1 short hairpin RNA (shRNA; knockdown) and SGC7901 cells overexpressing CKIP-1.
Results
The levels of CKIP-1 protein were significantly lower in the intestinal type of GC tissues compared with the samples of intestinal metaplasia. Both the levels of CKIP-1 protein and the levels of apoptosis decreased gradually with decreasing cell differentiation in the intestinal type of GC tissue and cell lines; and they were positively correlated. In the CKIP-1 shRNA group, the rate of apoptosis and the levels of Bax, cleaved caspase-3 and cleaved caspase-9 were decreased; and the levels of Bcl-2, Ras and the ratio of p-ERK/ERK were increased, compared with the control group. Opposite results were observed in the CKIP-1 overexpression group.
Conclusion
High levels of CKIP-1 protein may promote apoptosis in the intestinal type of GC, possibly via the downregulation of the Ras/ERK signalling pathway.
Introduction
Gastric cancer (GC) is one of the most common malignant tumours of the digestive tract and is divided into intestinal, diffuse, mixed and uncertain types by the Lauren classification. 1 In China, the incidence of GC is high and the main histopathological type is intestinal. 2 It has been suggested that the intestinal type of GC develops through a sequence of histological changes, including diffuse chronic gastritis, mucosal atrophy, intestinal metaplasia (IM), dysplasia, and finally invasive carcinoma, 3 but the exact molecular mechanisms involved in the development of the intestinal type of GC remain unclear. Casein kinase 2 interacting protein 1 (CKIP-1) is a casein kinase 2 interacting protein. 4 Currently, research suggests that the CKIP-1 gene might play an important role in the occurrence and development of several malignancies.5–9 However, the specific role of CKIP-1 in GC remains unclear. Previous research by the current authors demonstrated that the CKIP-1 gene was related to GC cell differentiation and involved in the proliferation, migration and invasion of GC, acting as a tumour suppressor gene. 10 Recently, research showed that CKIP-1 might promote apoptosis. 11 Apoptosis plays an important role in the occurrence and development of tumours. 12 The relationship between CKIP-1 and apoptosis in the intestinal type of GC has not been reported. Whether CKIP-1 acts as a tumour suppressor gene by promoting cancer cell apoptosis remains unclear. The present study investigated the relationship between CKIP-1 and apoptosis in the intestinal type of GC.
Patients and methods
Patients and tissue samples
This study obtained resected gastric tissue specimens from the Department of Pathology, Guizhou Provincial People’s Hospital, Guiyang, Guizhou Province, China between 2013 and 2016. Based on the Lauren classification system, two senior pathologists (Y.C. and M.C.) reviewed the haematoxylin-eosin (H&E) stained sections and confirmed that they were specimens of the intestinal type of GC. In each patient, there was gastric mucosal epithelium adjacent to the tumour and samples with areas of IM. The H&E slides of GC were also reviewed to analyse clinical and pathological parameters. The patients that were included in the study were those with complete clinical information available. None of the patients had received chemotherapy or radiation therapy before surgery.
The Research Ethics Committee of Guizhou Provincial People’s Hospital approved this study (2019 no. 54) and the study design was exempt from the requirement for informed consent. The waiver did not affect the rights and welfare of the study participants that provided tissue specimens.
Cell lines
Human cell lines of the intestinal type of GC (well differentiated MKN28 cells, moderately differentiated SGC7901 cells and poorly differentiated BGC823 cells) were obtained from the Shanghai Institute of Biological Sciences Cell Bank (Shanghai, China). Cells were cultured in Dulbecco’s modified Eagle’s medium (Hyclone Laboratories, Logan, UT, USA) supplemented with 10% fetal calf serum (Gibco BRL Life Technologies, Gaithersburg, MD, USA) in a humidified atmosphere containing 5% CO2/95% air at 37°C.
Immunohistochemistry
Immunohistochemical staining of CKIP-1 protein in the samples of the intestinal type of GC was performed as described previously. 13 Briefly, antigen retrieval was performed in 3% hydrogen peroxide at room temperature for 10 min and then the sections were treated with blocking buffer for 30 min at room temperature. The sections were then incubated with anti-CKIP-1 antibody (1:500 dilution; ab113663; Abcam, Cambridge, MA, USA) at 4°C overnight. Next, the sections were washed three times for 5 min each in 0.05% phosphate-buffered saline (PBS)-Tween-20 (pH 7.4–7.8) and then incubated with biotinylated goat anti-rabbit immunoglobulin G (IgG) (1:500 dilution; Thermo Fisher Scientific, Waltham, MA, USA) for 60 min at room temperature. Next, the sections were washed three times for 5 min each in 0.05% PBS-Tween-20 (pH 7.4–7.8). The sections were subsequently incubated with an avidin-biotinylated enzyme complex and 3,3'diaminobenzidine. The optical density (OD) of CKIP-1-like immunoreactivity (IR) was measured using a CM-2000B Biomedical Image Analysis System (Beihang University, Beijing, China). Five random fields of each area of interest were measured and the OD measurements were expressed as the mean.
TUNEL
Terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) was used to determine the levels of apoptosis in tissue sections. The following procedures were performed according to the manufacturer’s instructions. Staining of fragmented DNA in apoptotic cells was carried out using a TUNEL Apoptosis Detection Kit (with fluorescein isothiocyanate [FITC] as the fluorescent label; Yeasen, Shanghai, China) and the apoptosis index (AI) was calculated. The number of apoptotic cells and the total number of cells were counted in five randomly selected fields under a high-power automated inverted microscope (DMI4000 B; Leica, Wetzlar, Germany). The AI was calculated as follows: number of apoptotic cells/total number of cells × 100%. Slides in which the terminal deoxynucleotidyl transferase enzyme was replaced with double-distilled water served as the negative controls and slides treated with DNase I served as the positive controls.
Plasmids, lentiviral production and transfection
A sequence (5ʹ-CCTGAGTGACTATGAGAAGCTTCTCATAGTCACTCAGG-3ʹ) was designed and cloned into the pLKO.1 vector (Addgene Plasmid #1864) to generate a lentiviral plasmid of CKIP-1 short hairpin RNA (shRNA). The cDNA of CKIP-1 was subcloned into the PLenti CMV vector (Addgene Plasmid #17482) to get a lentiviral overexpression plasmid. Empty plasmid was used as a negative control. Lentiviruses were produced and transfected as previously described. 10
Western blot analysis
The protein levels of CKIP-1, B cell lymphoma-2 (Bcl-2), Bcl-2 associated X protein (Bax), cleaved cysteinyl aspartate specific protease 3 (cleaved caspase-3), cleaved caspase-9, rat sarcoma (Ras), extracellular signal-regulated kinase 1 and 2 (ERK1/2), phosphorylated extracellular signal-regulated kinase 1 and 2 (p-ERK1/2) and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in SGC7901 cells in the CKIP-1 shRNA group and CKIP-1 overexpression group were detected using Western blot analysis as described previously. 13 Briefly, total protein was extracted and separated by 10% sodium dodecyl sulphate–polyacrylamide gel electrophoresis and then transferred onto polyvinylidene difluoride (PVDF) membranes (Pall Biotech, Westborough, MA, USA). After blocking with 5% dry milk for 1 h at room temperature, the PVDF membranes were incubated with anti-CKIP-1 antibody, anti-Bcl-2 antibody, anti-Bax antibody, anti-cleaved caspase-3 antibody, anti-cleaved caspase-9 antibody, anti-Ras antibody, anti-ERK1/2 antibody and anti-p-ERK1/2 antibody (all 1:2000 dilution; Abcam) or anti-GAPDH antibody (1:20000 dilution; Sigma-Aldrich, St Louis, MO, USA) for 2 h at room temperature. After washing three times in 0.05% PBS-Tween-20 (pH 7.5), the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG or anti-mouse IgG (1:5000 dilution; Thermo Fisher Scientific) for 90 min at room temperature. Next, the membranes were washed three times in 0.05% PBS-Tween-20 (pH 7.5). Finally, the PVDF membranes were incubated with enhanced chemiluminescence reagent (ECL Plus; Thermo Fisher Scientific) for 5 min and the signals were visualized by exposure to Hyper Performance Chemiluminescence film (Pall Biotech).
Flow cytometry
The levels of apoptosis were measured in the intestinal type of GC cell lines using flow cytometry. Briefly, cells were seeded in 6-well plates at 1 × 106 cells/well for these experiments. When the cell density reached 80%, the cells were collected using 0.25% trypsin without ethylenediaminetetra-acetic acid. The cells were washed twice with 0.01 mM PBS (pH 7.2–7.4). Next, the cells were incubated in 400 µl binding buffer with 5 µl of FITC-labelled Annexin V and 5 µl of 50 µg/ml propidium iodide (PI) solution for 30 min at room temperature in the dark. Then, the samples were placed on ice in the dark and analysed by flow cytometry within 1 h using a FACSCalibur™ flow cytometer (BD Biosciences, San Jose, CA, USA). In the resulting cytograms, the top right quadrant represented the proportion of late apoptotic cells (Annexin V/FITC and PI double stained), while the bottom right quadrant represented the proportion of early apoptotic cells (Annexin V/FITC stained). The proportion of apoptotic cells (early apoptotic and late apoptotic cells) was expressed as a percentage of the total number of cells. Each treatment was performed in triplicate and three replicate experiments were performed.
Statistical analyses
All statistical analyses were performed using the SPSS® statistical package, version 15.0 (SPSS Inc., Chicago, IL, USA) for Windows®. Data from the groups are expressed as the mean ± SD. The means were compared using Student’s t-test or one-way analysis of variance. The correlation between CKIP-1 expression and apoptotic level was evaluated using Spearman correlation analysis. A P-value < 0.05 was considered statistically significant.
Results
The present study analysed surgically resected gastric tissue specimens from 55 patients. For each patient, there was gastric mucosal epithelium adjacent to the tumour and 40 samples with areas of IM were also collected. Positive immunohistochemical staining of CKIP-1 was mainly located in the cytoplasm. In gastric mucosal epithelial cells adjacent to cancer tissues, CKIP-1 was weakly stained (Figure 1a, white arrow). However, the CKIP-1-like IR was strikingly increased in areas of IM (Figure 1a, black arrow). Compared with CKIP-1 staining in the IM areas, the CKIP-1-like IR declined significantly in the intestinal type of GC tissues (Figures 1b–1d) (P < 0.01). The ODs of the CKIP-1-like IR in gastric mucosal epithelium, IM and intestinal type of GC group of patients are shown in Table 1.

Representative photomicrographs showing immunohistochemical staining of casein kinase 2 interacting protein 1 in samples of gastric mucosal epithelium, intestinal metaplasia (IM) and intestinal type of gastric cancer from patients (n = 55). (a) gastric mucosal epithelium (white arrow) and IM (black arrow); and (b, c, d) well, moderate and poorly differentiated intestinal type of GC (black arrow), respectively. Scale bar 100 µm. The colour version of this figure is available at: http://imr.sagepub.com.
Optical density levels determined during the immunohistochemical staining of casein kinase 2 interacting protein 1 (CKIP-1) in samples of gastric mucosal epithelium, intestinal metaplasia and intestinal type of gastric cancer from patients (n = 55).
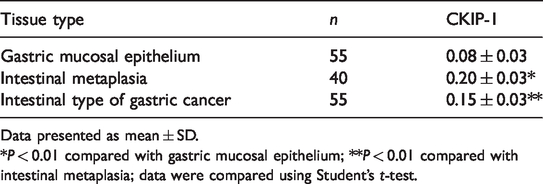
Data presented as mean ± SD.
*P < 0.01 compared with gastric mucosal epithelium; **P < 0.01 compared with intestinal metaplasia; data were compared using Student’s t-test.
The correlations between CKIP-1 levels in the intestinal type of GC and the clinicopathological characteristics of the patients are presented in Table 2. The levels of CKIP-1 were significantly associated with cancer cell differentiation and the age of the patients (P < 0.01 for both comparisons). The higher the level of differentiation of the cancer cells, the higher the levels of CKIP-1 (Figures 1b–1d). However, there was no significant correlation between CKIP-1 levels and sex, depth of invasion, lymph mode metastasis, tumour-node-metastasis (TNM) stage and vascular or nerve invasion.
The relationship between levels of casein kinase 2 interacting protein 1 (CKIP-1) in samples of intestinal type of gastric cancer from patients (n = 55) and their clinicopathological characteristics.

Data presented as mean ± SD.
aData were compared using Student’s t-test or one-way analysis of variance; NS, no significant between-group difference (P ≥0.05).
TNM, tumour-node-metastasis.
The level of apoptosis within the intestinal type of GC tissues of patients was measured using the TUNEL method and observed under a fluorescence microscope. As shown in Figure 2, in well differentiated cancer tissues, a large number of apoptotic cells were found, while in moderately and poorly differentiated cancer tissues, there were fewer apoptotic cells. The AI decreased with the decreasing degree of differentiation of cancer cells (P < 0.01; well differentiated cancer tissues compared with the moderately and poorly differentiated cancer tissues). The levels of CKIP-1 protein were positively correlated with the AI in the intestinal type of GC tissues from patients (r = 0.761; P < 0.05).
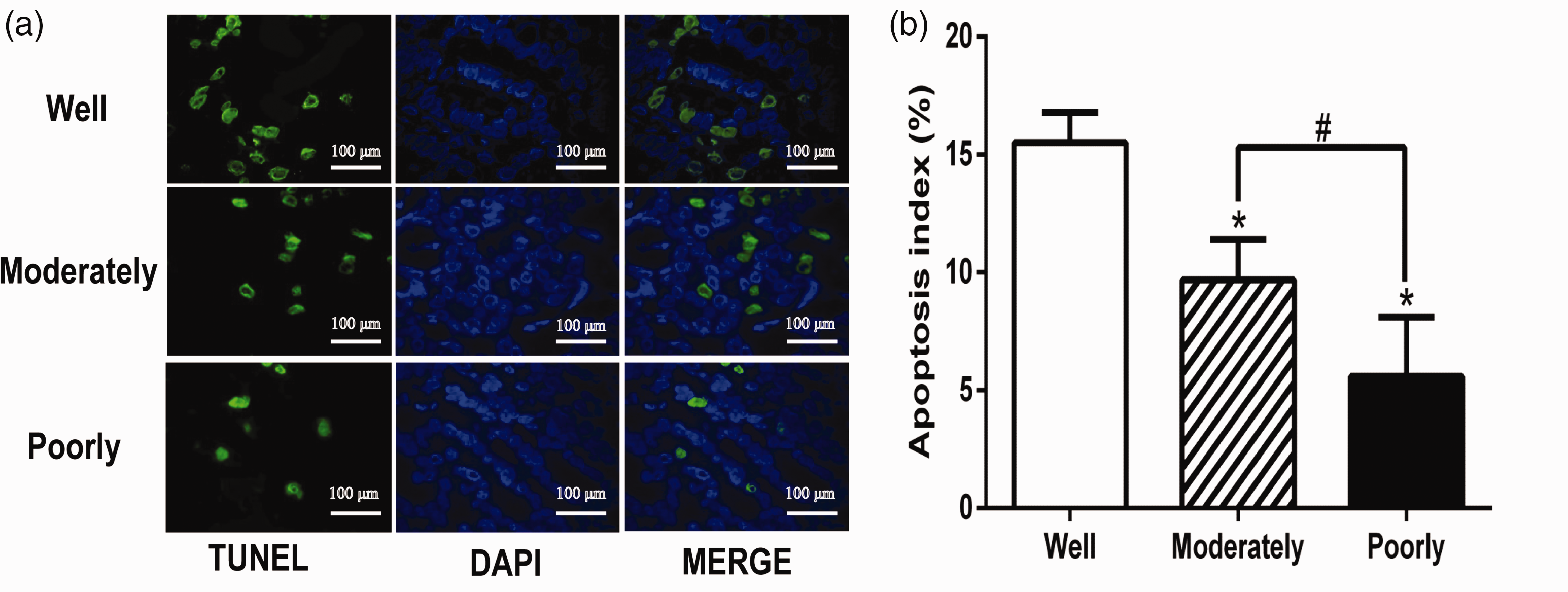
The levels of apoptosis as measured using terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) of tissue sections of intestinal type of gastric cancer from patients (n = 55) categorized based on the degree of differentiation (a). Scale bar 100 µm. The mean ± SD apoptosis index for each corresponding differentiation status (b). *P < 0.01 compared with the well differentiated tissues; #P < 0.05 compared with the moderately differentiated tissues; data were compared using one-way analysis of variance. DAPI, 4′,6-diamino-2-phenylindole. The colour version of this figure is available at:http://imr.sagepub.com.
Both the levels of CKIP-1 protein and the rate of apoptosis gradually decreased with the decreasing degree of differentiation of the intestinal type of GC cell lines (P < 0.05 for all comparisons) (Figure 3). Spearman correlation analysis showed that the levels of CKIP-1 protein were positively correlated with the rate of apoptosis in the intestinal type of GC cell lines of various differentiation states (r = 0.967; P < 0.01).

The protein levels of casein kinase 2 interacting protein 1 (CKIP-1) as measured by Western blot analysis (a); the levels of apoptosis as measured by flow cytometry (b); and the rate of apoptosis (c) in intestinal type of gastric cancer cell lines. Data presented as mean ± SD. *P < 0.05, **P < 0.01 compared with MKN28 cells; #P < 0.01 compared with SGC7901 cells; data were compared using one-way analysis of variance. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; FITC, fluorescein isothiocyanate. The colour version of this figure is available at: http://imr.sagepub.com.
The study established CKIP-1 shRNA and overexpression cell models using the SGC7901 cell line (Figure 4a) and measured the rate of apoptosis in the CKIP-1 shRNA and CKIP-1 overexpression groups. The results showed that the rate of apoptosis in the CKIP-1 shRNA group was significantly reduced compared with the control group (P < 0.01); and the rate of apoptosis in the CKIP-1 overexpression group was significantly increased compared with the control group (P < 0.05) (Figures 4b and 4c).

The protein levels of casein kinase 2 interacting protein 1 (CKIP-1) as measured by Western blot analysis (a); the levels of apoptosis as measured by flow cytometry (b); and the rate of apoptosis (c) in CKIP-1 short hairpin RNA (shRNA) SGC7901 cells and CKIP-1 overexpression SGC7901 cells. Data presented as mean ± SD. *P < 0.01 and **P < 0.05 compared with the control cells; data were compared using one-way analysis of variance. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; FITC, fluorescein isothiocyanate. The colour version of this figure is available at: http://imr.sagepub.com.
Compared with the blank control group, the levels of Bcl-2 protein were significantly increased, and the levels of Bax, cleaved caspase-3 and cleaved caspase-9 were significantly decreased, in the CKIP-1 shRNA group (P < 0.05 for all comparisons) (Figure 5). In contrast, the levels of Bcl-2 protein were significantly decreased, and the levels of Bax, cleaved caspase-3 and cleaved caspase-9 were significantly increased, in the CKIP-1 overexpression group compared with the control group (P < 0.01 for all comparisons).
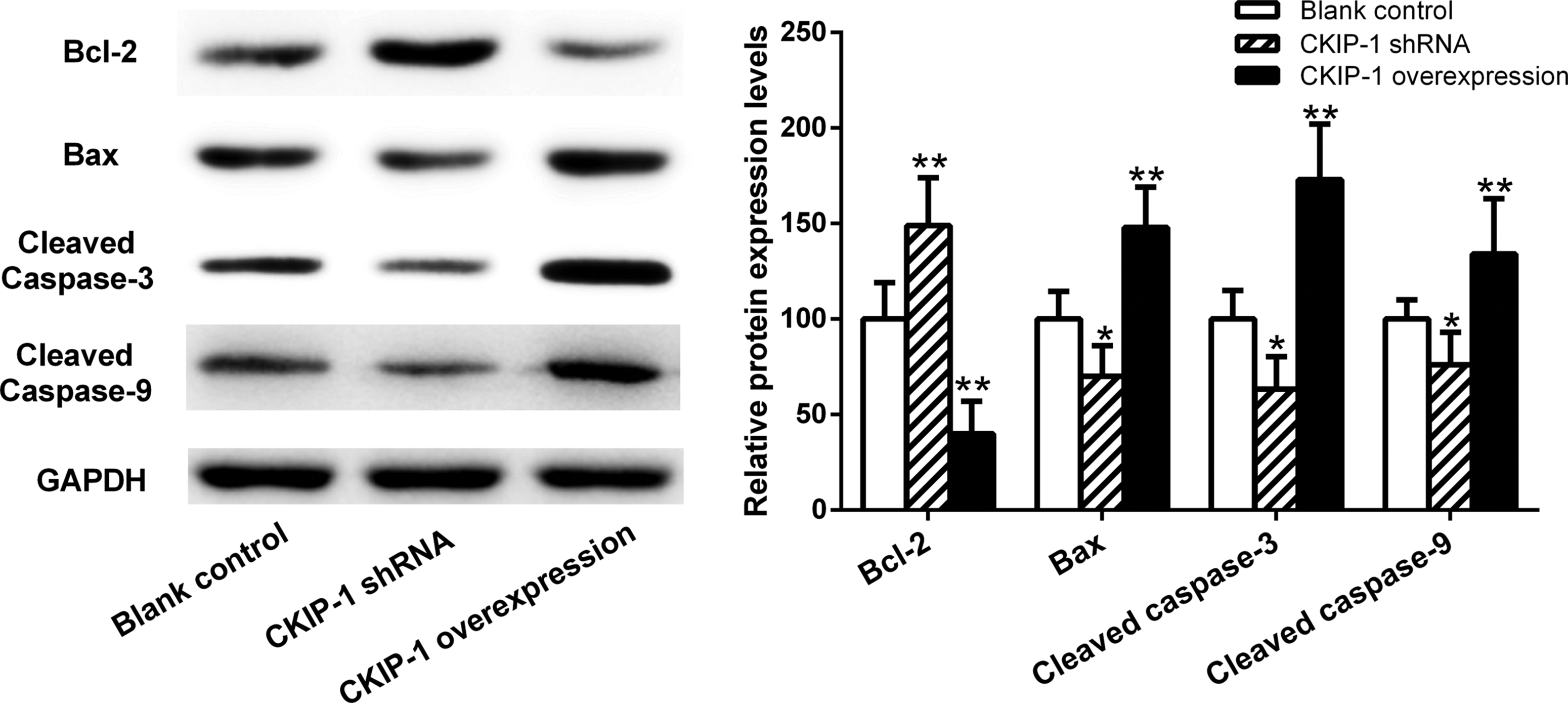
The protein levels of B cell lymphoma-2 (Bcl-2), Bcl-2 associated X protein (Bax), cleaved cysteinyl aspartate specific protease 3 (cleaved caspase-3) and cleaved caspase-9 as measured by Western blot analysis in casein kinase 2 interacting protein 1 (CKIP-1) short hairpin RNA (shRNA) SGC7901 cells and CKIP-1 overexpression SGC7901 cells. Data presented as mean ± SD. *P < 0.05 and **P < 0.01 compared with the control cells; data were compared using one-way analysis of variance. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
To evaluate whether the Ras/ERK signalling pathway could suppress apoptosis in the intestinal type of GC cells, the study measured the levels of the proteins involved in the Ras/ERK signalling pathway using Western blot analysis. The data showed that the levels of Ras and the ratio of p-ERK/ERK were significantly increased in the CKIP-1 shRNA group, while significantly decreased in the CKIP-1 overexpressed group, compared with the control group (P < 0.05 for all comparisons) (Figure 6).
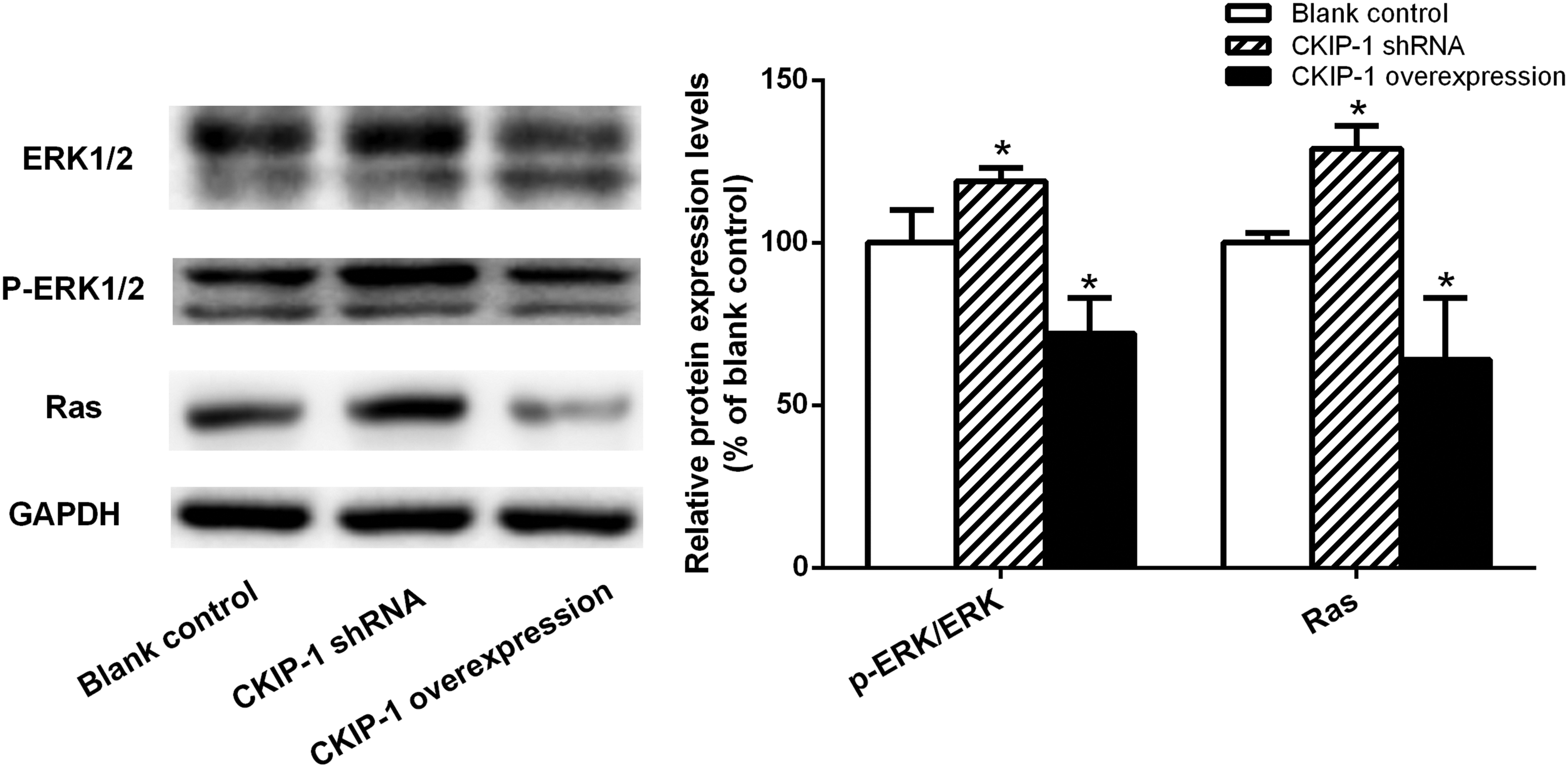
The protein levels of extracellular signal-regulated kinase 1 and 2 (ERK1/2), phosphorylated extracellular signal-regulated kinase 1 and 2 (p-ERK1/2) and rat sarcoma (Ras) as measured by Western blot analysis in casein kinase 2 interacting protein 1 (CKIP-1) short hairpin RNA (shRNA) SGC7901 cells and CKIP-1 overexpression SGC7901 cells. Data presented as mean ± SD. *P < 0.05 compared with the control cells; data were compared using one-way analysis of variance. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Discussion
At present, research on CKIP-1 mainly focuses on bone diseases.14–16 Recently, the role of CKIP-1 in malignancies has attracted more attention. For example, studies have shown that CKIP-1 is a scaffold protein that may be involved in the tumorigenesis and progression of colorectal cancer, 5 non-Hodgkin’s lymphoma 8 and glioma. 9 CKIP-1 might function as a potential tumour suppressor gene in these malignancies.5,8,9 In contrast to these previous reports, another study demonstrated that CKIP-1 mRNA was highly expressed in non-small cell lung cancer tissue and that CKIP-1 knockdown could promote cell apoptosis and inhibit tumour cell growth. 7 Although CKIP-1 has been studied in several types of tumour, the expression levels and the possible role of CKIP-1 in the intestinal type of GC remains unclear. To explore the possible role of CKIP-1 in the intestinal type of GC, this current study first undertook the immunohistochemical staining of CKIP-1 in healthy tissues and intestinal type of GC tissues from patients. These current results showed that there was only weak CKIP-1 staining in the gastric mucosal epithelial cells adjacent to cancer tissues, but CKIP-1 staining was increased in areas of IM. When compared with the levels of staining in the areas of IM, the CKIP-1 staining declined in the intestinal type of GC. These current results suggest that the levels of CKIP-1 protein were normally low in gastric mucosal epithelium, whereas when IM occurs, the gastric mucosal epithelium obtains an intestinal phenotype and higher levels of CKIP-1, similar to the levels of CKIP-1 observed in the intestinal mucosa. When IM progresses to intestinal type of GC along Correa’s cascade, the levels of CKIP-1 protein staining decreases, which suggests that CKIP-1 acts as a tumour suppressor in the development of the intestinal type of GC. This current study also investigated the correlations between CKIP-1 levels and the clinicopathological characteristics of patients with GC. These current data showed that the levels of CKIP-1 were associated with the degree of differentiation of the intestinal type of GC and the age of the patient. The levels of CKIP-1 decreased gradually with decreasing differentiation level of cancer cells of patients, which was consistent with a previous study by this same research group. 10 The current study also observed the same results in cell lines of the intestinal type of GC with various differentiation states. Poor differentiation of tumour cells means that there is greater cell atypia and a higher the degree of malignancy, with the tumour cells being more prone to invasion and metastasis, leading to patients being diagnosed at a later clinical stage. 17 These current data suggest that CKIP-1 may serve as a negative regulator in the intestinal type of GC. However, these current data did not demonstrate that the levels of CKIP-1 were significantly associated with the depth of invasion, lymph mode metastasis, TNM stage and vascular or nerve invasion. A previous in vitro study showed that upregulation of CKIP-1 can inhibit GC cell proliferation, migration and invasion; and that downregulation of CKIP-1 can promote GC cell proliferation, migration and invasion. 10 Due to the small number of patients in this current study, it will be necessary to increase the number of patients in future studies to clarify the relationship between CKIP-1 and invasion and metastasis of cancer.
Apoptosis is one of the basic pathways in cells and it plays an important role in the physiological and pathological processes of the body. 12 Inhibition of apoptosis is an important process in tumorigenesis and the promotion of apoptosis may provide new methods for tumour treatment.12,18 Recent studies reported that CKIP-1 could promote apoptosis. 11 CKIP-1 can be cleaved by caspase-3, releasing C-terminal fragments and travelling to the nucleus, thereby inhibiting the activity of c-jun in promoting the apoptosis of tumour cells during tumour necrosis factor-induced apoptosis. 11 In addition, CKIP-1 can form a positive feedback loop with caspase-3 to enhance tumour cell apoptosis. 11 To explore the relationship between CKIP-1 and apoptosis in the intestinal type of GC, this current study first detected the levels of apoptosis in tissue samples of the intestinal type of GC from patients and in cell lines in vitro. The levels of apoptosis gradually decreased with decreasing cell differentiation in both human tissue samples and in cell lines, which was similar to the changes observed for CKIP-1 levels. The levels of CKIP-1 protein were positively correlated with the AI in human intestinal type of GC tissues from patients (r = 0.761; P < 0.05) and the levels of CKIP-1 protein were positively correlated with the rate of apoptosis in intestinal type of GC cell lines of various differentiation states (r = 0.967; P < 0.01). These results suggest that CKIP-1 might act as a tumour suppressor gene in the intestinal type of GC by promoting cancer cell apoptosis.
To explore whether CKIP-1 could influence cell apoptosis in the intestinal type of GC cells, this current study selected the intestinal type of GC cell line SGC7901 and used it to establish CKIP-1 knockdown and overexpression cell lines for further studies on the effect of CKIP-1 on apoptosis. The current results demonstrated that the rate of apoptosis of cancer cells in the CKIP-1 shRNA group was inhibited, and the rate of apoptosis in CKIP-1 overexpression group was increased. These current findings suggest that low levels of CKIP-1 may inhibit apoptosis in SGC7901 cells, while high levels of CKIP-1 may promote apoptosis in SGC7901 cells. It has been reported that a variety of proteins are closely related to apoptotic pathways, which jointly affect tumour growth and malignant transformation. 19 For example, proteins of the Bcl-2 and caspase families are known to modulate apoptosis through the regulation of the mitochondrial apoptosis pathway. 20 This present study analysed the relationship between the levels of CKIP-1 protein and the levels of Bcl-2, Bax, cleaved caspase-3 and cleaved caspase-9 proteins. These current results showed that the level of Bcl-2 protein was significantly increased and the levels of Bax, cleaved caspase-3 and cleaved caspase-9 proteins were significantly decreased in the CKIP-1 shRNA group compared with the control group. In the CKIP-1 overexpression group, the opposite results were observed. These current findings suggest that high levels of CKIP-1 protein in SGC7901 cells may promote apoptosis via the downregulation of Bcl-2 and the upregulation of Bax, cleaved caspase-3 and cleaved caspase-9.
Various signalling pathways mediate GC cell apoptosis, including phosphatidylinositol-3-kinase (PI3K), nuclear factor-ƙB and Ras/ERK pathway. 21 The Ras/ERK pathway is frequently activated and plays a vital role in the development and maintenance of cancers. 22 The levels of ERK1/2 in the tumour tissues of GC were significantly increased compared with paracarcinoma tissues. 23 The Ras/ERK signalling pathway promoted the growth and metastasis of the GC cell lines SNU-16 and MKN1. 22 Activation of the Ras/ERK signalling pathway promotes survival by repressing the expression of proapoptotic proteins and elevating antiapoptotic proteins. 24 Silencing of Ras in vitro blocks the Ras signal transduction pathway and increases apoptosis in carcinoma cell lines. 25 It was reported that CKIP-1 negatively regulates several important pathways, such as the transforming growth factor-β/bone morphogenetic protein, Wnt/β-catenin and PI3K/protein kinase B signalling pathways.16,26–28 However it is not clear whether CKIP-1 is related to the Ras/ERK signalling pathway. To clarify this, the current study measured the protein levels of Ras and ERK1/2 in CKIP-1 shRNA and overexpressed SGC7901 cells. The current results showed that the levels of Ras and the ratio of p-ERK/ERK were both significantly increased in CKIP-1 shRNA SGC7901 cells, while they were significantly decreased in CKIP-1 overexpressed SGC7901 cells compared with the control group. On the basis of these current findings, the high levels of CKIP-1 appear to promote apoptosis in the intestinal type of GC by downregulating the Ras/ERK signalling pathway.
In conclusion, these current results indicate that high levels of CKIP-1 may promote apoptosis in the intestinal type of GC, which might involve the downregulation of the Ras/ERK signalling pathway. These studies provide support for the suggestion that CKIP-1 acts as a tumour suppressor gene in the development of intestinal type of GC.