
Abstract
Objective
To investigate the effects of salidroside on cognitive dysfunction induced by chronic cerebral hypoperfusion in rats.
Methods
Male Sprague–Dawley rats (n = 36) were divided into three groups (n = 12 per group): sham operation; bilateral permanent occlusion of the common carotid arteries (2-VO); 2-VO + salidroside. Rats received 20 mg/kg per day salidroside or vehicle intraperitoneal injection beginning the day before surgery and continuing until 34 days postoperatively. Cognitive function was evaluated by Morris water maze test and hippocampal long-term potentiation (LTP) measurement. Hippocampal neuronal apoptosis was evaluated via immunofluorescence.
Results
Chronic cerebral hypoperfusion caused marked cognitive deficit and LTP inhibition. These effects were largely ameliorated by salidroside administration. Salidroside prevented caspase-3 activation, increased the ratio of Bax/Bcl-2, and reversed hippocampal neuronal loss induced by chronic cerebral hypoperfusion.
Conclusions
Salidroside prevents cognitive deficits caused by chronic cerebral hypoperfusion in rats, and alleviates apoptosis in the hippocampal CA1 area.
Introduction
Senile dementia is a progressive neurodegenerative disorder, characterized by severe memory loss and cognitive impairment; vascular dementia is the second most common form of dementia in the ageing population. 1 To date, there are no specific drugs to cure, delay or prevent vascular dementia. Bilateral permanent occlusion of the common carotid arteries (2-VO) in rats results in a substantial reduction in cerebral blood flow, and is a well-characterized animal model used to investigate the cognitive consequences of chronic cerebral hypoperfusion. 2 These animals exhibit learning and memory impairments, accompanied by neuronal degeneration and microvascular abnormalities, resembling those found in human subjects with vascular dementia. 3 Studies based on this rat model have tested potentially beneficial strategies to prevent, delay or reverse the progression of dementia associated with reduced cerebral blood flow. 4
Rhodiola rosea is a popular medicinal plant found at high altitudes in the mountainous regions of Tibet, and has been used in traditional Tibetan medicine for centuries. It has antiradiation and antifatigue effects, improves cognitive deficit, delays the effects of ageing on the neural system and resists anoxia.
5
The main bioactive component of R. rosea is salidroside (p-hydroxyphenethyl-β-
Material and methods
Experimental animals
A total of 36 male Sprague–Dawley rats (mean weight, 240 ± 20 g; supplied by the Experimental Animal Centre, The Fourth Military Medical University, Xi’an, China) were assigned to one of three groups (n = 12 per group): (i) sham group (sham operated rats treated with normal saline); (ii) 2-VO group (2-VO rats treated with normal saline); (iii) 2-VO salidroside group (2-VO rats treated with salidroside [Yi-Le Pharmaceutical Co. Ltd., Shaanxi, China]). Rats were housed (n = 6 per cage) at a temperature of 22 ± 2℃, with a 12-h light/12-h dark cycle and free access to food and water. Daily 20 mg/ml salidroside intraperitoneal injection (1 ml/kg per day) or vehicle (normal saline) was started on the day before surgery and continued until the termination of the experiment (34 days postoperatively). Salidroside dose was determined based on an earlier study
13
and our preliminary experiments. During the last 7 days of the experiment (days 28–34 postoperatively), spatial learning and memory were assessed 1 h after drug administration. The experimental procedure is shown in Figure 1. The research protocol was approved by the Institutional Animal Ethics Committee of The Fourth Military Medical University, Xi’an, China and the study protocol complies with international rules and policies.
Experimental design and timeline (in days) for a study investigating the effects of salidroside on cognitive dysfunction induced by chronic cerebral hypoperfusion in rats.
Surgical procedures
Surgery was carried out as described. 14 Briefly, under 320 mg/kg chloral hydrate intraperitoneal anaesthesia, a ventral midline incision was made. The bilateral common carotid arteries were exposed and gently separated from the carotid sheath and vagus nerve. In rats assigned to the 2-VO and 2-VO salidroside groups, the two arteries were ligated with a 5-0 silk suture. Rats in the sham-operated groups received the same operation but the carotid arteries were not ligated. During the surgery, the body temperature of the rats was maintained at 37.5 ± 0.5℃ until the rats recovered to thermal homeostasis.
Morris water maze test
The Morris water maze test was carried out daily between day 28 and 34 after surgery, using described methods 15 with slight modifications. The apparatus comprised a circular tank (diameter 120 cm, height 50 cm) filled with water (temperature 22 ± 1℃, depth 25 cm), which was made opaque with milk so that the rats were unable to see the underwater platform. The tank was divided into four quadrants and was surrounded by several prominent cues to spatial co-ordinates, including items such as a shelf, several posters and illumination lights. Rats were trained to locate a hidden platform (diameter 10 cm), which was centred at one of the quadrants (target quadrant) and submerged ∼2 cm below the surface of the water. The swimming paths of the rats were monitored by a computerized video tracking system and swimming velocity was measured during each test.
During the training period (5 consecutive days) rats were given eight trials per day to find the hidden platform using four different starting points (equally spaced around the perimeter of the tank). All four starting points were used once in a clockwise sequence. Rats were given 60 s to find the platform and sit on it for 30 s. Rats that failed to find the location within the given time were guided to the platform and were allowed to stay on it for 30 s. Then they were picked up and placed in the water for the next trial. The time taken to find the platform (escape latency) was measured, and the mean over the eight trials was calculated. After the eighth trial, all rats were towel-dried and placed in their home cages. Investigators who conducted the Morris water test were blind to the treatment groups.
A probe test, in which the hidden platform was removed, was conducted on the day before the first training and the day after the last training. Rats were placed in the water at the opposite point to the target quadrant and were allowed to swim freely for 120 s in the tank. The percentage of time in the target quadrant was recorded as an assessment of spatial memory.
Electrophysiological recordings
The right cerebral hemisphere of six rats from each group was used for this analysis. Rats were sacrificed via isoflurane anaesthesia, the brain was rapidly removed, and 2 transverse hippocampal slices (400 -µm thick) were prepared from each rat. Slices were cut with a vibratome in ice-cold artificial cerebrospinal fluid (ACSF) solution (126 mM sodium chloride, 2.5 mM potassium chloride, 2.4 mM calcium chloride, 1.2 mM magnesium chloride, 1.2 mM monosodium phosphate, 19 mM sodium bicarbonate and 11 mM glucose, gassed with 95% oxygen and 5% carbon dioxide (pH 7.35–7.45). The slices were incubated in 95% oxygen/5% carbon dioxide-equilibrated ACSF at 34℃ for ≥1 h. Each slice was then placed on the recording chamber and submerged in continuously flowing ACSF (2 ml/min) gassed with 95% oxygen and 5% carbon dioxide (pH 7.35–7.45), and heated to 34℃.
Field excitatory postsynaptic potentials (fEPSPs) were recorded in the CA1 stratum radiatum using glass micropipettes with 2–4 MΩ electrical resistance and filled with 3 M sodium chloride. Schaffer collateral fibres were stimulated with bipolar stimulation electrodes (tip separation 300 µm). Stimulations were generated by a Master-8 pulse generator and a stimulus isolator (A.M.P.I, Jerusalem, Israel). Recorded electrical signals were amplified with an Axopatch™ 700B amplifier (Molecular Devices Corporation, Sunnyvale, CA, USA) filtered at 3 kHz. Data were acquired using a computer equipped with the Digidata® 1440A acquisition system (Molecular Devices Corporation) and analysed using pCLAMP™ version 10.2 software (Molecular Devices Corporation).
The fEPSP was evoked by a square-wave pulse (100 μs duration, 0.05 Hz). Prior to the experiment, an input/output curve was made to determine the stimulus intensity required to produce a half-maximal response. All subsequent stimuli were delivered at this intensity. Baseline responses were recorded for 30 min after stable baseline responses were obtained, and the mean of 12 consecutive fEPSPs was calculated. Long-term potentiation (LTP) was induced by the application of high-frequency stimulation (HFS) (twice for 1 s at 100 Hz, applied with a 20-s interval). After HFS, stimulation resumed at 0.05 Hz and recordings were taken for another hour. The slope of evoked fEPSP was normalized to the mean fEPSP measured during the baseline recording period. Results were expressed as mean ± SD of percentages relative to control baseline.
Western blotting
The left cerebral hemisphere of six rats from each group was used for this analysis. Hippocampus homogenates were prepared in radio-immunoprecipitation assay lysis buffer, containing 50 mM Tris (pH 7.4), 150 mM sodium chloride, 1% Triton X™-100, 1% sodium deoxycholate, 0.1% SDS, 2 mM sodium fluoride, 5 mM ethylenediaminetetra-acetic acid (pH 8.0), 1 mM sodium orthovanadate (Beyotime Inc., Jiangsu, China), and the protease inhibitor, phenylmethylsulphonyl fluoride (1 mM). Protein concentrations were determined using the Pierce™ BCA® Protein Assay Kit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Equal amounts of total protein (20 µg per lane) were separated by sodium dodecyl sulphate–polyacrylamide gel electrophoresis followed by transfer to polyvinylidene difluoride membranes. Nonspecific binding was blocked by incubation with 3% bovine serum albumin in Tris-buffered saline containing Tween 20 (TBST) for 1 h. Next, the membranes were incubated at 4℃ overnight with rabbit anti-rat Bcl-2 (1 : 400 dilution), rabbit anti-rat Bax (1 : 1000 dilution), or rabbit anti-rat active caspase-3 (1 : 500 dilution) antibodies (Abcam, Cambridge, UK). After washing three times with TBST, the membranes were incubated with goat anti-rabbit IgG-horseradish peroxidase (1 : 2000 dilution; Zhongshan Goldenbridge Ltd., Beijing, China) for 2 h at room temperature. Mouse anti-rat monoclonal antibody against β-actin (Sigma–Aldrich, St Louis, MO, USA) was used as the loading control. Enhanced chemiluminescence (Thermo Scientific) was used to develop the membrane. Relative intensities of the bands were quantified by densitometric analysis.
Histology and immunofluorescence
Morphological analysis was performed after the probe trials as described.
16
A total of six rats from each group were anaesthetized with 320 mg/kg chloral hydrate intraperitoneal injection and perfused intracardially with 0.9% saline, followed by 4% paraformaldehyde, in 0.1 M phosphate buffer (pH 7.4). The brains were immediately removed, immersed in 4% paraformaldehyde for 24 h, and cryoprotected in a 30% sucrose solution. Coronal sections were cut at 25 -µm thickness by a frozen-stage microtome. Finally, the sections were rinsed in 0.01 M phosphate-buffered saline (PBS) and mounted onto gelatin-coated glass slides. Sections were labelled using immunofluorescence. After blocking for 1 h at 37℃ with 10% goat serum and 0.3% Triton X
Statistical analyses
Data were presented as mean ± SD. Morris water maze escape latencies and fEPSP slopes were analysed using two-way analysis of variance (ANOVA) with repeated measures, followed by the Tukey–Kramer test. Other data were analysed using one-way ANOVA followed by Duncan multiple group comparison. Statistical analysis was performed using SPSS® version 13.0 (SPSS Inc., Chicago, IL, USA) for Windows®. P-values <0.05 were considered statistically significant.
Results
Data regarding Morris water maze performance are shown in Figure 2. In acquisition trials, rats gradually learned the location of the hidden platform, indicated by decreased escape latency duration over the 5-day training period (Figure 2A). Rats in the 2-VO group had significantly impaired learning compared with animals in both other groups (P < 0.05 for both comparisons; Figure 2A). In the probe trial, there were no significant between-group differences in time spent in the target quadrant before training (Figure 2B). After training, rats in the 2-VO group spent significantly less time in the target quadrant compared with animals in both other groups (P < 0.05 for both comparisons). There were no significant between-group differences in mean swimming velocity (Figure 2C).
Morris water maze performance deficit in rats with chronic cerebral hypoperfusion, induced by bilateral permanent occlusion of the common carotid arteries (2-VO) with or without salidroside treatment, or sham-operated rats. (A) Mean daily escape latency time (from test starting point onto a hidden platform). (B) Percentage of time spent in the target quadrant in the probe trial (no platform). (C) Swimming velocity. Data are mean ± SD (n = 12). *P < 0.05 vs sham group. **P < 0.05 vs 2-VO group.
High frequency stimulation of Schaffer collateral fibres led to an initial increase in fEPSP slope in all groups. LTP was significantly impaired in the 2-VO group compared with the sham and 2-VO salidroside groups (P < 0.05; Figure 3). There was no significant difference in LTP between the 2-VO salidroside and sham groups.
Schaffer collateral-CA1 LTP inhibition in rats with chronic cerebral hypoperfusion induced by bilateral permanent occlusion of the common carotid arteries (2-VO), with or without salidroside treatment, or sham-operated rats. Upper trace represents typical recordings of field excitatory postsynaptic potentials (fEPSP) at 15 min pre-high frequency stimulation (HFS) (a,c,e) and 60 min post-HFS (b,d,f). Long-term potentiation was significantly impaired in the 2-VO group compared to the sham and 2-VO salidroside groups. Data are mean ± SD (n = 12). *P < 0.05 vs sham group. **P < 0.05 vs 2-VO group.
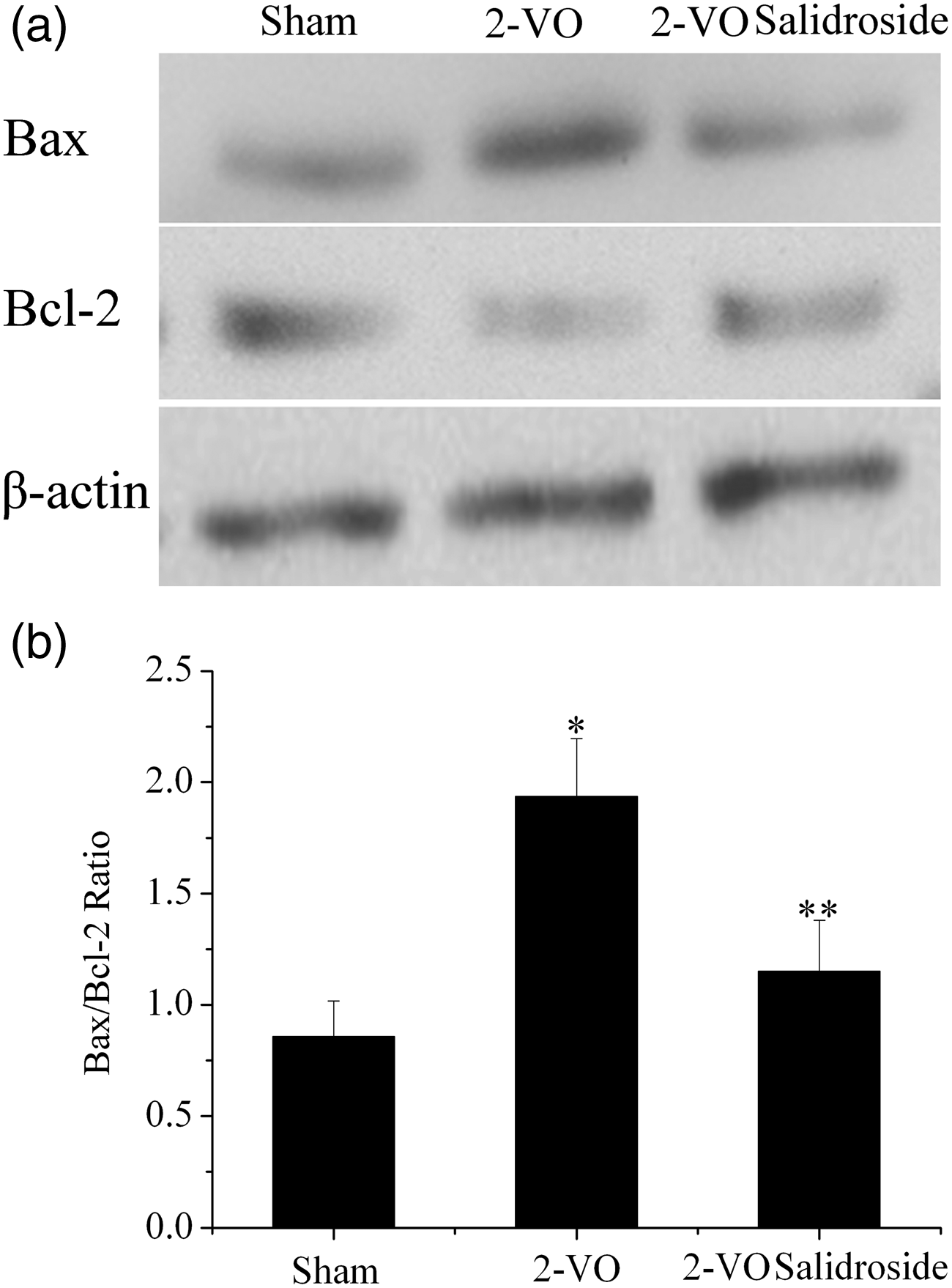
Western blotting analysis of active caspase-3 in the hippocampus is shown in Figure 4; Bax and Bcl-2 are shown in Figure 5. The hippocampal concentration of active caspase-3 was significantly higher in the 2-VO group compared with both other groups (P < 0.05 for each comparison; Figure 4). The hippocampal Bax/Bcl-2 ratio was significantly higher in the 2-VO group compared with both other groups (P < 0.05 for each comparison; Figure 5).
Hippocampal concentrations of active caspase-3 in rats with chronic cerebral hypoperfusion, induced by bilateral permanent occlusion of the common carotid arteries (2-VO), with or without salidroside treatment, or sham-operated rats. (a) Representative Western blot of active caspase-3 and β-actin (control). (b) Densitometric quantification of active caspase-3 protein normalized to β-actin. Data are mean ± SD (n = 6). *P < 0.05 vs sham group. **P < 0.05 vs 2-VO group. Hippocampal concentrations of Bax and Bcl-2 in rats with chronic cerebral hypoperfusion induced by bilateral permanent occlusion of the common carotid arteries (2-VO), with or without salidroside treatment, or sham-operated rats. (a) Representative Western blot of Bax, Bcl-2, and β-actin. (b) Densitometric quantification of Bax and Bcl-2, expressed as Bax/Bcl-2 ratio. Data are mean ± SD (n = 6). *P < 0.05 vs sham group. **P < 0.05 vs 2-VO group.
The findings of NeuN immunofluorescence are shown in Figure 6. Staining was visibly reduced in hippocampal CA1 regions from 2-VO animals compared with both other study groups. Staining in the sham and 2-VO salidroside groups appeared to be of similar intensity.
Representative fluorescence confocal photomicrographs of hippocampal CA1 sections stained with anti-NeuN in rats with chronic cerebral hypoperfusion induced by bilateral permanent occlusion of the common carotid arteries (2-VO) with or without salidroside treatment, or sham-operated rats. (a) Sham group. (b) 2-VO group. (c) 2-VO salidroside group. Original magnification × 200; scale bar, 10 µm. The colour version of this figure is available at: http://imr.sagepub.com.
Discussion
Animal experiments demonstrate that chronic cerebral hypoperfusion can induce memory deficits. 2 In addition, the 2-VO rat is recognized as a popular model for studying the pathophysiology of learning and memory deficits in vascular dementia, as well as for assessing therapeutic potential and mechanism of putative antidementia drugs.2,17 Cerebral blood flow decreases by 25–50% at 2.5 h after the 2-VO operation, reducing progressively for 1 week, with the effect and lasting for several months. 18
To investigate the effects of salidroside on cognitive dysfunction in 2-VO rats, in the present study we evaluated cognitive function using the Morris water maze test and hippocampal LTP. Next, we evaluated neuronal apoptosis of the hippocampus to explore the underlying mechanism by which salidroside prevents cognitive impairment.
The water maze has been consistently used to identify deficits in spatial learning and memory in global cerebral ischaemia, in rats. 19 In accordance with the findings of others, 2 spatial learning and memory of rats in the 2-VO group were significantly impaired in the present study, as demonstrated by longer escape latency and a shorter time spent in the target quadrant in the Morris water maze, compared with sham-operated rats. Salidroside administration effectively ameliorated this memory impairment, in accordance with a study using streptozotocin intracerebroventricular injection. 11 Since there were no between-group differences in swimming velocity in the current study, the longer escape latency seen was not a result of motor injury; rather it was the result of an impairment of learning and spatial memory capacity.
Although the correlation between LTP and cognitive function is debatable, 20 LTP may be a cellular model of learning involved in initial encoding of information, at least in the hippocampus, where this information is maintained for a substatntial period of time before being potentially transferred to the cortex.21,22 HFS-evoked LTP in the CA1 region of rat hippocampus is thought to be related to memory.21,23 In the hippocampus, the CA1 region is the most vulnerable to cerebral hypoperfusion. 24 We therefore investigated the effect of salidroside on Schaffer collateral-CA1 LTP inhibition induced by chronic cerebral hypoperfusion. Our findings suggest that LTP in the Schaffer collateral-CA1 synapses was significantly inhibited by chronic cerebral hypoperfusion, and that salidroside counteracted this LTP inhibition. This paralleled the preventive effects of salidroside on impairment of learning and spatial memory. These results provided possible electrophysiological evidence that salidroside prevents cognitive impairment.
The mechanism underlying 2-VO-induced cognitive deficit is complex. Neuronal apoptosis of the hippocampus is related to cognitive deficit in rats with induced chronic cerebral hypoperfusion. 25 Caspase-3 is a principal effector caspase in the apoptotic cascades that lead to neuronal apoptosis: caspase-3-null mice lose developmental neuronal apoptosis, whereas other effector caspase knockout models (such as caspase-6) do not.26,27 Active caspase-3 is therefore considered to be the final common path in apoptosis. Salidroside effectively inhibited the activation of caspase-3 induced by chronic cerebral hypoperfusion in the present study. Bax and Bcl-2, two members of the Bcl-2 family, are crucial regulators of apoptosis. Bax is a proapoptotic protein, whereas Bcl-2 is an antiapoptotic protein, and the interaction between these proteins determines cell fate. As such, the Bax/Bcl-2 ratio may better predict the apoptotic status of a cell than the absolute concentration of either protein. 28 Salidroside significantly counteracted the chronic cerebral hypoperfusion-induced upregulation of Bax and down regulation of Bcl-2 in the present study, reducing the Bax/Bcl-2 ratio in the hippocampus. Immunofluorescence revealed significant neuronal loss in the hippocampal CA1 area after 2-VO in our study, consistent with the findings of others 29 , and salidroside administration reversed this neuronal loss. These results were consistent with our biochemical findings. In accordance with our findings, salidroside pretreatment has been shown to attenuate hydrogen peroxide-stimulated apoptotic cell death in a dose-dependent manner, in a primary culture of hippocampal neurons. Thus, the antiapoptotic action of salidroside might be mediated by inhibiting caspase-3 activity. 30
The present study has some limitations. Although neuronal loss was observed in 2-VO rats, the neuronal types were unclear. The preventive effect of salidroside on vascular dementia was confirmed, but the therapeutic effect of salidroside requires further investigation.
In conclusion, salidroside prevents cognitive deficits caused by chronic cerebral hypoperfusion in rats, and alleviates apoptosis in the rat hippocampal CA1 area. The antiapoptotic effects of salidroside might contribute to the improvement in cognitive function in rats that received this agent, in the present study.
Footnotes
Declaration of conflicting interest
The authors declare that there are no conflicts of interest.
Funding
This work was supported by a research grant from the Natural Science Foundation of China (81201000, 81100816).
Acknowledgements
We thank Medjaden Bioscience Limited, Hong Kong, China, for assisting in the preparation of this manuscript.