
Abstract
Advances in stem cell technology have provided three approaches to address the demanding issue of the treatment of intractable neurological disease. One of the approaches is the screening of compounds attenuating pathological phenotypes in stem-cell based models. A second approach consists of exogenous-targeted cell supplementation to the lesion with stem cell-derived differentiated cells. A third approach involves in vivo direct programming to transdifferentiate endogenous somatic cells and to boost CNS tissue remodeling. In this review, we outline research advances in stem cell technology of direct reprogramming in vitro and in vivo and discuss the future challenge of tissue remodeling by neural transdifferentiation.
Introduction
Stem cell technology opened a new road for advancement of the clinical approach of regenerative medicine for incurable central nervous system (CNS) diseases.1,2 The availability of induced pluripotent stem cells (iPSCs)-derived human neurons is now leading the way toward the accessibility of novel drugs for incurable neurodegenerative diseases. 3 Another regenerative medical approach with iPSCs technology is that of cell supplementation into affected CNS regions, contributing to new autologous transplantation therapy for incurable diseases.4,5
Stem cell technology has resulted in the novel research field of transcriptional factors (TFs)-mediated cell fate conversion in vitro. 6 Furthermore, previous studies have demonstrated that this reprogramming technique using defined TFs can be applicable to not only in vitro but also in vivo direct reprogramming to transdifferentiate somatic cells across different cell lineages in both acute CNS injury and neurodegenerative disease model rodents. 7 This should offer the next promising milestone for future regenerative medicines for CNS diseases, since it is a less invasive approach than cell transplantation and does not need to depend upon immunosuppressive therapy (Figure 1).
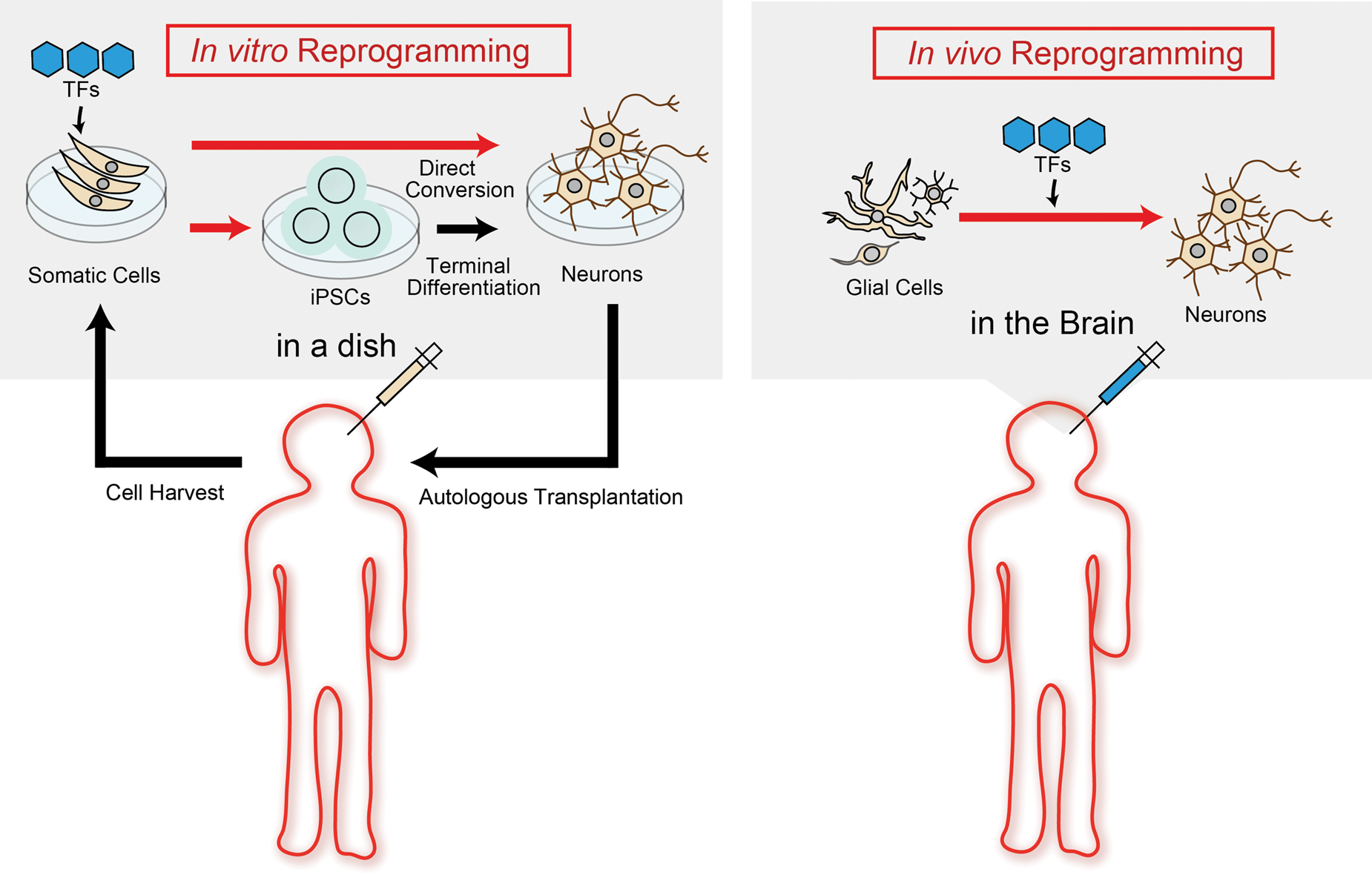
In vitro and in vivo reprogramming for neural transdifferentiation. Left panel (In vitro reprogramming): Cells derived from patients are reprogrammed into iPSCs or targeted cells using defined transcriptional factors (TFs) transfer in a dish; Right panel (In vivo reprogramming): Glial cells are directly reprogrammed into targeted cells using defined TFs transfer in the brain.
In the acute phase of CNS diseases such as ischemia and traumatic injury, gliosis accompanying neuronal cell death is a common pathological change, where glial cells including astrocytes, oligodendrocyte precursor cells (OPC) and microglia proliferate or/and migrate to be activated for the clearance of debris or filling-out sparse space and focal tissue remodeling of injured lesion. 8 In general, neurological symptoms depend on the severity of the injury to mature neurons, implying that glial cells surrounding injured core lesions could be a main cell source for in vivo direct conversion into functional neurons. In healthy CNS, an operational cellular assembly between endothelial cells, glial cells and neurons is called a neuro-vascular unit (NVU), whose crosstalk is critical for the integration of each functional activity. 9 Therefore, acute neuronal loss could disrupt this NVU homeostasis, the cell-cell interaction, and also cause further delays in NVU and neural network remodeling. This suggests that early therapeutic intervention aimed at facilitating the NVU remodeling can improve the prognosis of patients with acute CNS injury.
Gliosis is also a common pathological change associated with chronic cell death in neurodegenerative diseases and might precipitate hazardous effects on disease progression and even determine the disease onset in some diseases.10,11 In contrast, gliosis may also have a protective role in pathogenesis by the clearance of pathogenic protein accumulated in the brain in other neurodegenerative diseases.12,13 Therefore, appropriate selection of the original cell source and target cells would be important for the future clinical application of in vivo reprogramming in CNS diseases.
Here, we outline the advances of stem cell technology for reprogramming and discuss the future prospects of in vivo direct reprogramming to transdifferentiate neural cells for tissue remodeling in acute CNS injury and for disease modifying for chronic neurodegenerative diseases.
In vitro reprogramming to determine cell fate
Forced ectopic expression of genes that are developmentally silenced in closed chromatin is critical for the reprogramming process. The discovery of ‘Yamanaka-TFs 14 enabled us to recognize the existence of pioneer TFs that are critical chromatin modifiers engaging in targeted sites on nucleosomal DNA and opening chromatin, and they differ from conventional TFs that simply regulate gene expression by binding the promoter region of DNA. 15 Pioneer TFs have cis-acting elements to access tightly packed chromatin and induce nucleosome remodeling, whereas specifier TFs have DNA-binding motifs that bind exposed trans-acting elements of promoter regions to regulate gene expression. Epigenetic regulators including histone modifiers (e.g. histone deacetylases: HDACs, histone acetyltransferases: HATs) also play cis-acting regulatory roles in gene expression that allows passive access of several TFs to the promoter region of a set of genes.16,17 Therefore, the in vitro reprogramming process to determine the cell fate of somatic cells can be divided into two operations – ‘Cis-reprogramming’ by pioneer TFs or epigenetic regulators and ‘Trans-reprogramming’ by specifier TFs. For example, among ‘Yamanaka-TFs’, Oct4, Sox2 and Klf4 are known to act by cis-reprogramming as pioneer TFs, whereas c-Myc only binds to opened chromatin to act by trans-reprogramming as secondary enhancer, which is not necessary for generating iPSCs. 18 Pioneer TFs drive somatic cells to be multi-potent cells that can have the potential to differentiate into subtypes of neurons that could be determined by defined specifier TFs as additional maturation steps. Cell fate diversity can be dependent on transition via a progenitor cell (Figure 2).
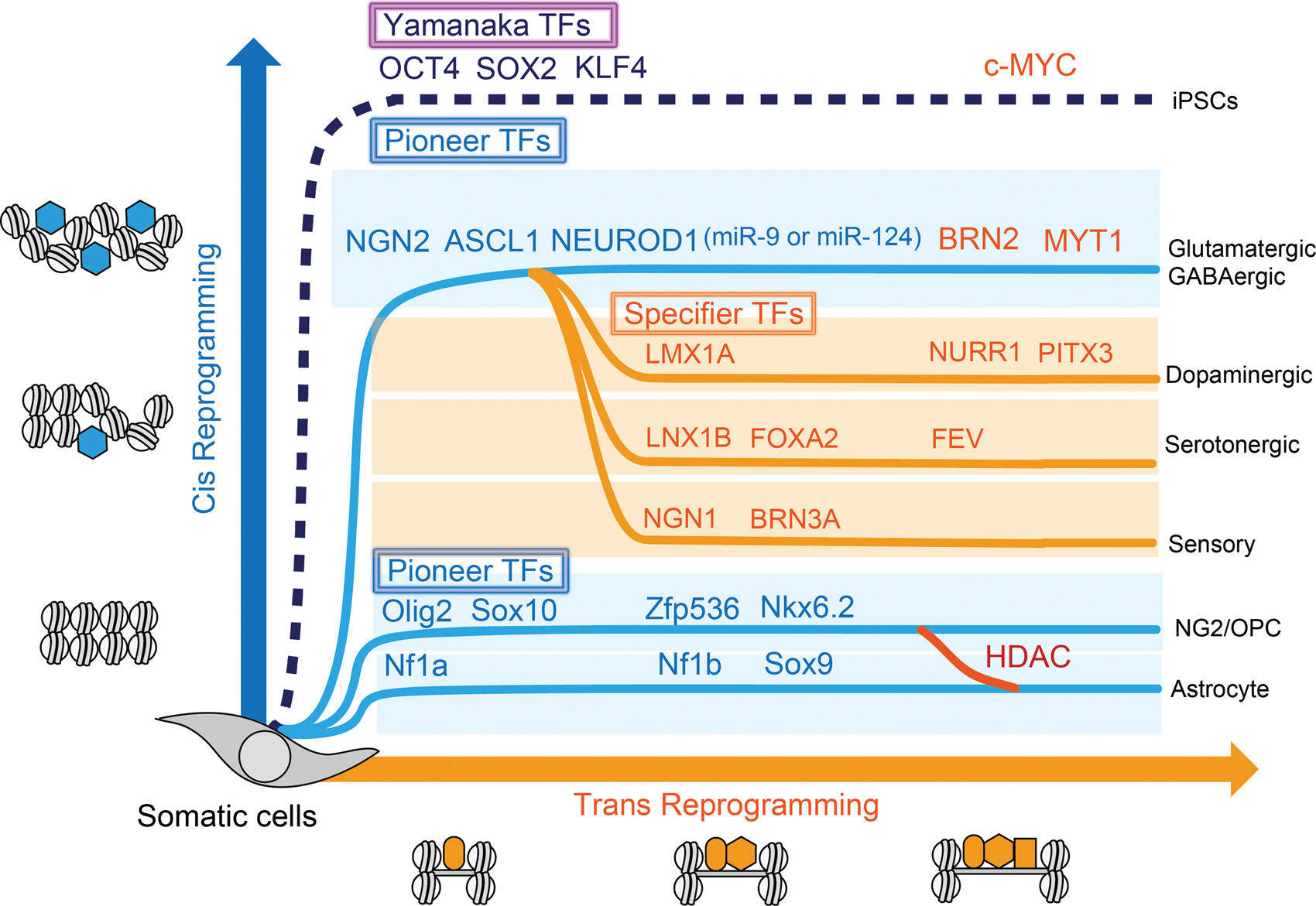
Two operations of in vitro direct reprogramming for somatic cells. Pioneer transcriptional factors (TFs) (blue hexagons) open chromatin and involve in cis-reprogramming events and specifier TFs (orange squares) involve in trans-reprogramming events as additional cooperative factors to activate gene regulatory network to terminal differentiation. Histone deacetylase (HDAC) could modulate glial cell fate between NG2/OPC and astrocytes.
After primary de-condensing of chromatin and exposing multiple gene promoters by the use of ectopic pioneer TFs or histone modifiers, in vitro reprogramming can be achieved by either of the following processes: (1) direct binding by ectopic specifier TFs: direct reprogramming, (2) passive/non-specific binding by endogenous specifier TFs: passive/indirect reprogramming.
In vitro direct reprogramming to determine cell fate
Stem cell technology has promoted the next step of TFs-mediated conversion of somatic cells into targeted terminated cells 19 (Table 1). Vierbuchen et al. 6 demonstrated the TF-initiated direct conversion into neurons by defined factors from fibroblasts in vitro and opened a new avenue into the research field related to direct conversion. They used a combination of only three TFs – achaete-scute homologue 1 (Ascl1), Brn2, and myelin transcription factor 1-like protein (Myt1l) (referred to as BAM) to generate synapse-forming functional neurons from mouse fibroblasts and human fibroblasts (iNs),20–22 where Ascl1 acts as a pioneer TF, Brn2 binds to the site of Ascl1, and Myt1l later binds to the specific site for neuron maturation 23 (Figure 2). The addition of pioneer TF, neurodifferentiation D1 (NEUROD1) to BAM, is necessary for efficient conversion from human fibroblasts into functional iN. 20 Neurogenin-2 (Ngn2) is also a pioneer TF that can convert astrocytes into neurons in vivo and in vitro.24,25 The combination of pioneer TFs, ASCL1 and NGN2 strongly convert human fibroblasts, even if derived from aging humans, into functional iNs.26,27 BAM plus NGN2 (termed BAMN) can convert T-cell lymphocytes in peripheral blood into neurons. 28 MicroRNAs (miRNAs) can convert fibroblasts into iN through NeuroD2 activation. 29
In vitro/in vivo direct reprogramming into target cell with differential transcriptional factors.
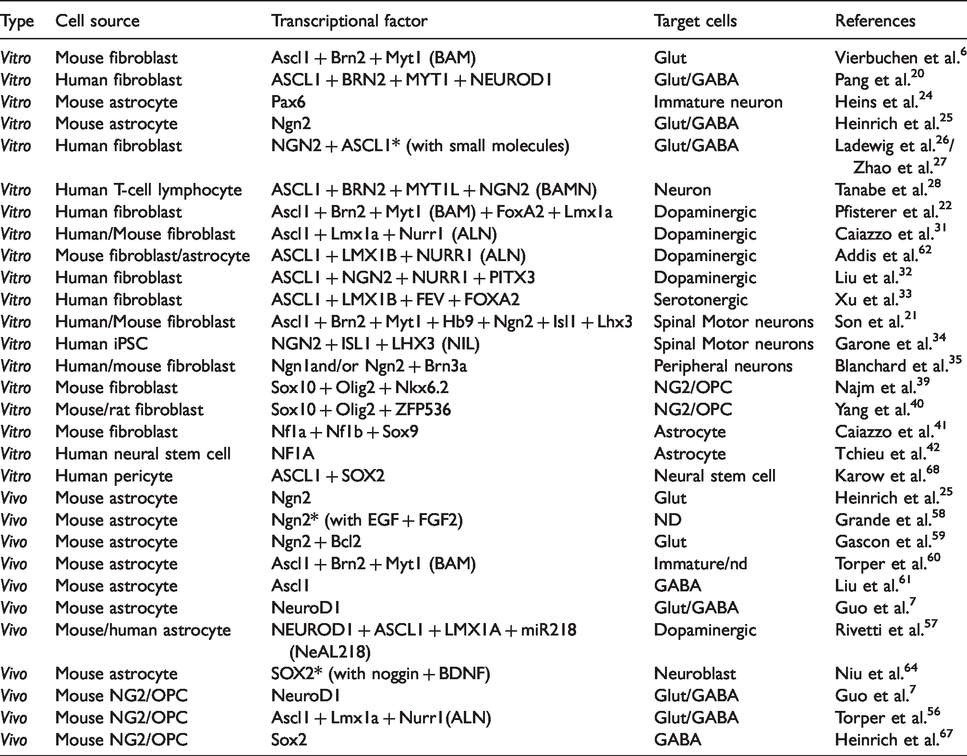
These iNs are prone to be characterized as glutamatergic neurons and/or a minor group of GABAergic neurons,20,26,29 suggesting that a set of endogenous specifier TFs can easily direct to these types of cells under open-chromatin condition as default iNs. To target a specific subtype of iNs such as dopaminergic neurons (iDAN),22,30–32 serotonergic neurons, 33 spinal motor neurons (MNs)21,34 or peripheral neurons, 35 defined sets of exogenous specifier TFs might be needed as shown in Table 1.
Other types of neuronal cells besides neurons can also be generated by a similar direct reprogramming technique. NG2 cells express NG2 chondroitin sulfate proteoglycan (CSPG4), 36 and a multipotent stem cell that can proliferate itself and can differentiate mainly into oligodendrocytes which were originally purified as bipotential oligodendrocyte-type2 astrocyte progenitor cells (O-2A cells). 37 Therefore, NG2 cells are also referred to as OPC or polydendrocytes. 38 Induced NG2/OPC (iOPC) can be directly generated from fibroblasts by Sox10 and oligodendrocyte transcriptional factor 2 (Olig2) with either zinc-finger protein 536 (ZFP536) or Nkx6.2. 39 , 40 Induced astrocytes (iAs) can also be generated from mouse fibroblasts through exogenous TFs of Nuclear factor 1A (Nf1a), Nf1b and Sox9. 41 A previous study has demonstrated that NF1A is enough for generating iAs from human neural stem cells. 42
Finally, emerging data suggest that neural crest-derived sources may also be a potential source for an endogenous transdifferentiation response. The neural crest is a unique and transient embryonic cell population that originates in the ectoderm within margins of the neural tube. After a phase of epithelial-mesenchymal transition and extensive migration, neural crest-derived stem cells (NCSCs) settle down in different parts of the body to contribute to the formation of a plethora of different organs and tissues via differentiation into neurons, glia (or Schwann cells), and mesenchymal derivatives. 43 Intriguingly, a subset of NCSCs may be present in bone marrow, meninges, dental tissue, gut, heart, and skin in adults.44,45 Neural crest cells may be directly induced from human fibroblasts by reprogramming with SOX10 along with a WNT activator 46 or with gene transfer of FOXD3 together with a chitosan substrate. 47 Applications of patient-specific NCSCs with the reprogramming technique may provide a better understanding of the regeneration of mesodermal and ectodermal cells and tissues in various disease conditions.
In vitro passive/indirect reprogramming to determine cell fate
Combinations of methylated DNA with acetylated/deacetylated histones are involved in “open” or “closed” chromatin configuration for gene expression, implying that nucleosomal modification could work as a cis-acting element and allow passive access of endogenous specifier TFs to the specific promoter region, that is, the so-called passive/indirect conversion.
Past studies demonstrated that NG2/OPC are not only restricted to generating oligodendrocytes but they also produce astrocytes and neurons under certain conditions. 48 The cell fate of NG2/OPC is regulated by DNA chromatin modifications as epigenetic regulation by epigenetic modifiers such as histone modifier, HDACs, HATs and DNA methyltransferases (DNMTs), suggesting that indirect reprogramming through epigenetic regulators could have the potential to reprogram NG2/OPC into neurons. HDAC1/2 double-knockdown suppressed terminal differentiation of NG2/OPC into oligodendrocytes.49,50 HDAC3 ablation led to an increase in astrocytes together with a loss of oligodendrocytes. 51 The authors showed that HDAC3 interacted with p300 HAT to open the chromatin configuration to activate oligodendrocyte lineage-specific genes. 51 These studies indicated that passive/indirect reprogramming through HDACs regulation could have the potential to transdifferentiate oligodendrocytes into NG2/OPC.
DNMTs consist of DNMT1, DNMT3A, and DNMT3B. DNMT1 is necessary for the maintenance of DNA methylation, and DNMT3A/3B are necessary for de novo DNA methylation. 52 DNMT1 is upregulated in NG2/OPC differentiation during the prenatal stage in vivo and in vitro, and its activity decreases with age. 53 DNMT1 supports NG2/OPC survival in vitro. Global DNA methylation by DNMT3A is required for efficient BAM-mediated in vitro reprogramming into iN, 54 suggesting that the activity of DNMTs would support cellular plasticity and upregulate the neural transdifferentiation potential.
In vivo reprogramming to determine cell fate
Advances of in vivo reprogramming studies have demonstrated that glial cells could be a cell source of neurogenesis after acute/chronic neuronal loss in disease progression. 55 TFs involved in brain development are expected to be important pioneer TFs that determine cell fate not only in vitro but also in terminally differentiated cells in vivo. Among them, major pioneer TFs common in in vivo direct reprogramming from glial cells are Ngn2, Ascl1 and NeuroD1 (Figure 3), and all these TFs are potent converters by cis-reprogramming action in vitro as mentioned above. It should be noted that specifier TFs in vivo direct reprogramming does not function the same as in vitro. For example, Ascl1, Lmx1a, and Nurr1 (referred to as ALN) are originally used for obtaining iDAN from fibroblasts in vitro, 31 but in vivo conversion using the ALN combination cannot direct iDAN, but other types of neurons, glutamatergic or GABAergic neurons. 56 An additional pioneer TF, NEUROD1, is necessary to achieve in vivo conversion from glial cells into iDAN. 57 These proposals suggest that a combination of potent pioneer TFs and additional specifier miRNAs, suppressors of the original TFs specifically expressed in the terminally differentiated cells, will be effective for in vivo direct reprogramming.
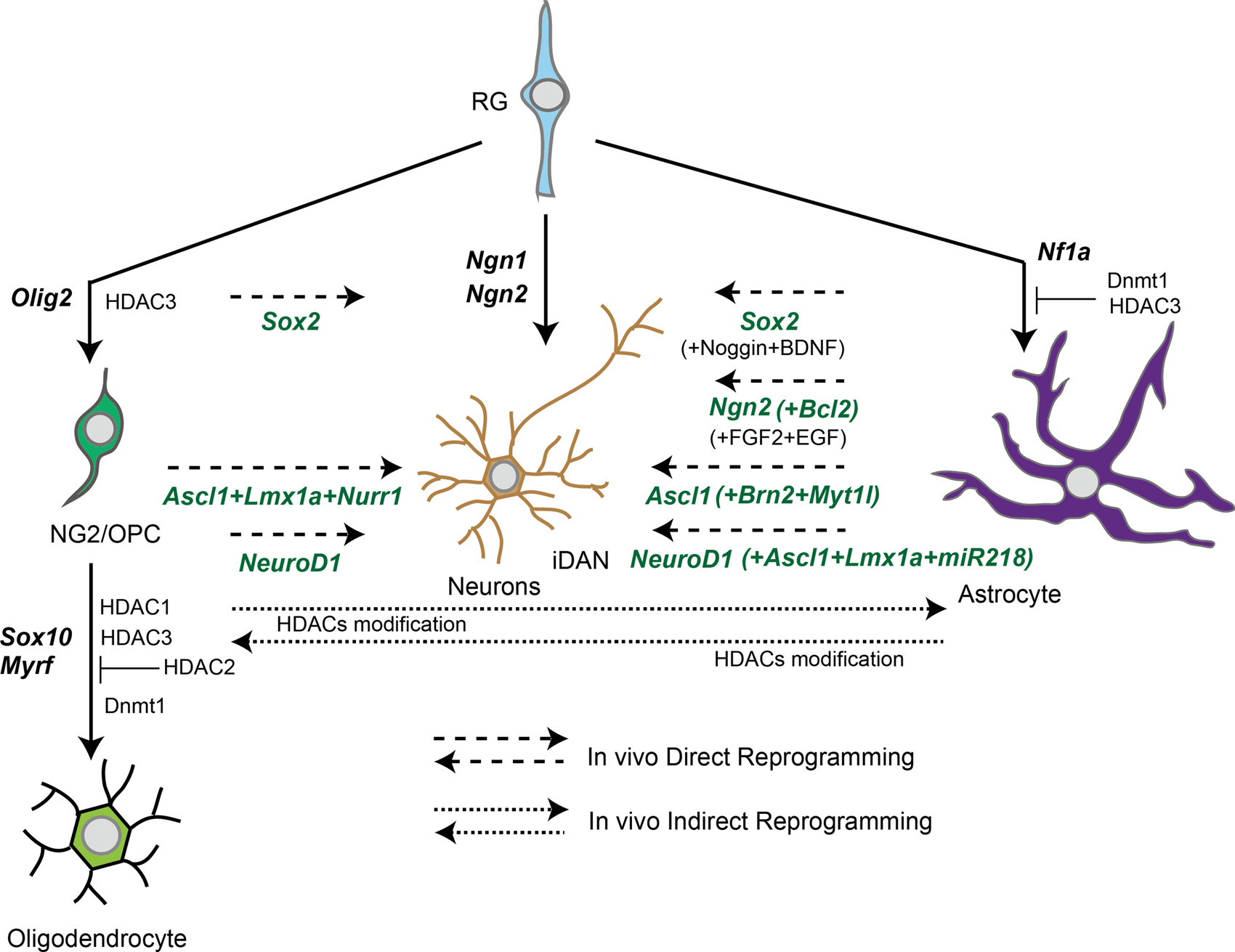
In vivo reprogramming between neurons and glial cells via pioneer transcriptional factors (TFs).Long dash lines indicate in vivo direct conversion pathway with pioneer TFs and specifier TFs. Short dash lines indicate in vivo indirect conversion pathway with epigenetic regulators. Solid lines indicate terminal differentiation with essential TFs for brain development. RG: radial glia; HDAC: histone deacetylase.
In vivo direct reprogramming from astrocytes to neurons (in vivo A-to-N)
Pax6 was the first identified TF to have the potential to reprogram glial cells into neuronal cells such as GABAergic or glutamatergic cells in vitro as the transcriptional factor expressed in radial glia and which positively regulated the expression of Ngn2. 24
Ngn2 has been identified as a pioneer transcript factor in in vivo A-to-N, which converted terminally differentiated astroglia into synapse-forming glutamatergic neurons. 25 A combination of the neurotrophic factors FGF2 and EGF could reprogram astrocytes into functional neurons in the neocortex and striatum even after traumatic injury. 58 The co-expression of Bcl2 with Ngn2 boosts the efficiency of conversion from astrocytes to glutamatergic neurons after stab wound injury. 59
Ascl1 is likely another potent pioneer TF in in vivo A-to-N. A combination of transcriptional factors of Ascl1, Brn2 and Myt1l converted mouse astrocytes into immature neuron-like cells in the mouse brain with low efficiency (∼6%). 60 Another group revealed that only Ascl1 could induce astrocytes in the midbrain into functional GABAergic neurons in postnatal mouse. 61 Efficient reprogramming of astrocytes to dopaminergic neurons can be achieved by a single polycistronic lentiviral vector carrying three transcription factors – ASCL1, LMX1B, and NURR1. The process was efficient, with about 18% of cells expressing markers of dopaminergic neurons after two weeks of in vitro study. 62
A third pioneer transcriptional factor is NeuroD1, which plays a role in neurogenesis 63 and can be used for reprogramming from reactive astrocytes into functional glutamatergic neurons with high effectivity after traumatic injury. 7 A previous study demonstrated that NEUROD1 with ASCL1, LMX1A, miRNA, miR218 (termed NeAL218) directly converted striatal astrocytes into iDAN. 57 They also succeeded in correcting the motor functional phenotype in a Parkinson’s disease (PD) model using iDAN.
Sox2 has also been identified as having the potential, as transcriptional factor, to reprogram glial cells into neuroblast cells. The subsequent addition of noggin and BDNF results in functionally mature neurons. 64
In addition to exogenous inducers of reprogramming, the endogenous responses of trans-differentiation may also be worth further investigation. Transcription factor-mediated in vivo reprogramming in the brain is mostly performed via stereotaxic injection of viruses for over-expression in the CNS. 55 Chemical approaches have also been described. Human astrocytes were sequentially treated to a cocktail of small molecules that inhibit glial but activate neuronal signaling pathways.65,66 This method successfully reprograms astrocytes into neurons in 8–10 days through epigenetic regulation and transcriptional activation.65,66 It has been reported that human astrocytes-derived neurons can survive for more than five to seven months in vitro and form functional synaptic networks with synchronous burst activities.65,66 Chemically reprogrammed human neurons can also survive for more than one month in mouse brain in vivo and integrate into local circuits. 66 When administered in vivo through intracranial or intraperitoneal injection, combinations of various small molecules significantly increase hippocampal neurogenesis in mice. 65 Chemical reprogramming of human astrocytes into functional neurons may therefore provide another translationally relevant approach to regenerate functional neurons from patients’ endogenous glial cells for brain repair.
In vivo direct reprogramming from NG2/OPC to neurons (in vivo O-to-N)
Guo et al. 7 found that NeuroD1, which is a potent TF in A-to-N, can convert NG2/OPC into GABAergic neurons and glutamatergic neurons. Torper et al. 56 demonstrated that improved delivery of the three TFs Ascl1, Lmx1a and Nurr1 (generally referred as ALN) using AAV could reprogram NG2/OPC into functional GABAergic neurons. In addition, Sox2 has also been reported to convert NG2/OPC into GABAergic neurons after stab-injury. 67 Interestingly, SOX2 can convert human pericytes into functional neurons with ASCL1 (in vitro study),68,69 suggesting that SOX2 could convert cells into ‘fractional’ neural stem-like cells as weak pioneer TFs. 70
In vivo reprogramming strategy for CNS diseases
In this section, we will introduce a potential therapeutic approach using an in vivo reprogramming technique to convert residential glial cells into functional CNS cells directly or to boost endogenous cellular plasticity to transdifferentiate into other types of neural cells after acute CNS injury and chronic neurodegenerative diseases. Blood–brain barrier (BBB) breakdown after disruption of NVU is a pathological feature common to both acute CNS injury and some chronic neurodegenerative diseases, creating an attractive delivery site of ectopic TFs for in vivo direct reprogramming.
In vivo reprogramming strategy for acute CNS injury
Representative acute focal injuries in CNS are traumatic injury and ischemic stroke. Both trauma and stroke result in vascular damage and the following BBB breakdown. There are three phases in tissue remodeling after acute CNS injury, as follows. 8 The first phase of acute CNS injury (within 1 day after injury) is cell death of parenchymal cells (i.e. neurons and glial cells) and platelet aggregation to serve the recruitment of inflammatory cells and residual/circulating immune cells through a leaky BBB. Neuronal cell death and apoptosis result from an insufficiency of energy such as lactate supplied by damaged glial cells and from massive Ca2+ influx into neurons. Infiltrated immune cells/microglia phagocytose cellular debris and damaged cells release danger-associated molecular patterns (DAMPs) and alarmins that promote non-infectious inflammation. Efficient removal of damaged glial cells protects neurons from myelin-associated inhibitory proteins for axonal regeneration secreted by damaged glial cells. 71
The second phase of acute CNS injury (2–10 days after injury) is tissue replacement by proliferation of intrinsic CNS cells such as endothelial progenitors, fibroblast-lineage cells and various glial cells. 72 Finally, the third phase of CNS injury (over 10 days after injury) is tissue remodeling under scar formation by functional astrocytes to surround fibroblast-lineage cells and pericytes and promote angiogenesis. Astrocyte scar formation serves as a border that protects healthy neurons in peri-lesions from further migration of inflammatory cells and cytotoxic cytokines and molecules. 73
An in vivo direct reprogramming approach may be effective during the second and third phases of active cellular proliferation for replacement in acute CNS injury. Endothelial progenitors, fibroblast-lineage cells and various types of glial cells and pericytes would be the source of cells of direct reprogramming by delivery of TFs. Conventional delivery methods consist of direct injection of virus vectors such as retrovirus and lentivirus. Previous studies have highlighted the potential application of exosome, which is a nanosized vesicle, as miRNA delivery.74–76 Delivery of engineered exosome, the half-life of which is increased by CD47 expression, 77 suppressed pancreatic cancer by silencing oncogenic KRAS in vivo. 78 Delivery of such engineered exosome packing TFs through leaky BBB in vivo might be a safe and promising approach for direct reprogramming into target neurons during the second and third phases after acute CNS injury.
Besides in vivo direct reprogramming, in vivo passive/indirect reprogramming using epigenetic modifiers may also be effective for early tissue remodeling in all phases after acute CNS injury. HATs bind to transformation-related protein (TRP53) to form a transcriptional complex, which enhances the accessibility of the promoter of regeneration-associated genes (RAGs), such as RAB13, CORO1B and growth-associated protein (GAP43). Increasing histone acetylation by HDAC inhibitors induced GAP43 expression as well as the improvement of axon regeneration after injury.79–81 HDAC5 may play an important role in axonal regeneration after injury via regulating microtubule dynamics.82,83 In addition, HDAC6 also contributes to axon regeneration, as its inhibition enhanced tubulin acetylation levels to promote axon regeneration of DRG neurons in the presence of myelin-associated glycoprotein. 84 Therefore, the epigenetic regulation of gene expression related to axon regeneration could offer a promising approach toward increasing the neuronal network after CNS injury.
BBB modeling using iPSC-derived cells has been intensively studied to mimic a neurovascular unit. A combination of endothelial cells derived from iPSCs with human primary pericytes and astrocytes in Transwell® or a microfluidic chip setting could mimic a BBB co-culture model with formation of a tight junction, suggesting that physiological contact between heterogenous cells derived from iPSCs could achieve self-BBB remodeling.85,86 BBB modeling using iPSC-derived cells can provide us with a novel platform where the delivery of TFs toward in vivo reprogramming can be examined.
Looking at cell-based regenerative medicine via cell-reprogramming in the clinical field of stroke, three transplantation clinical trials have largely succeeded in lab-to-clinic translational application of reprogrammed cells. The pioneering study by Layton Bioscience Inc. used a cocktail of growth factors and mitotic inhibitors that turned teratocarcinoma cells into post-mitotic-like neurons called NT2N.87–89 Subsequently, SanBio Inc. initiated clinical trials of intracerebral implantation of SB623® into patients in chronic stage of stroke. SB623® are bone marrow-derived cells that exhibit a neuronal phenotype by reprogramming with Notch-1 gene transfer.90–92 Reneuron Inc. started trials of transplanting immortalized fetal cortical cells called CTX0E03® via control of c-mycERTAM.93,94 These stem cell trials were intended to boost CNS tissue repair that supports the survival of intrinsic neurons, and combination with reprogramming technology could enhance the CNS tissue remodeling during the second and third phases after acute CNS injury.
Reprogramming strategy for neurodegenerative diseases
Tissue damage in neurodegenerative diseases accumulates gradually during chronic periods. Multifocal pathogenic processes evoke multifocal gliosis and multicellular response similar to those caused by acute injury with a breakdown of BBB and recruitment of neural cells and leukocytes, but on a smaller scale.
Pathogenic molecules such as beta-amyloid (Aβ) produced in Alzheimer’s disease (AD) can also act just as alarmins to trigger microglial activation to release IL-1β, TNF-α and IL-6. 95 Cytokine signaling links to activation of kinase that phosphorylates tau, another pathogenic protein. 96 However, microglia and astrocytes have a biphasic role in the AD pathological process, as they are known to serve as Aβ scavengers to clear out Aβ,12,13 a burden that precedes cognitive decline in AD patients by about 20 years. Attenuation of reactive astrocytes increased the Aβ load in AD model mice. 97 Therefore, direct programming of microglia and astrocytes into functional neurons in AD might not be effective for the prevention of disease progression.
Dysfunctional glial cells may trigger the onset, or facilitate progression of amyotrophic lateral sclerosis (ALS). Oligodendrocytes, microglia and astrocytes with mutation of responsive gene SOD1 for familial ALS mutation accelerate disease progression. 10 However, astrocytic NF-kB activation drives microglial proliferation that is rather protective against ALS disease progression in the pre-symptomatic phase. 98 Actually, exogenous glial-rich cells derived from iPSCs extended the survival of mutant SOD1 ALS mice, 99 implying that stage-dependent glial replacement would be effective for disease-modifying therapy.
Parkinson’s disease (PD) is characterized by a loss of dopaminergic neurons that innervate the striatum, where astrocytes are chronically activated. Microglia activate to convert normal astrocytes into neurotoxic A1 astrocytes, which kill neurons and other glial cells. 100 Pharmacological inhibition of A1 astrocyte conversion was neuroprotective for dopaminergic neurons in a PD rodent model, 101 suggesting that reprogramming astrocytes could be beneficial against the pathogenic process in PD. In vivo direct reprogramming from striatal astrocytes into iDAN in PD model mice reverted the motor dysfunction phenotype via generation of functional iDAN. 57
Collectively, the role of glia in the pathogenetic process is pivotal in neurodegenerative diseases, and selection of subtypes of glial cells depending on the disease stage is essential for future in vivo direct programming therapy for each neurodegenerative disease. In vivo direct reprogramming therapy towards gaining pericytes would be attractive for the neurodegenerative disease. Since about 50% of pericytes in BBB are lost in ALS, 102 the blood-spinal cord barrier in ALS would be permeable, suggesting that intravenous delivery of TFs might achieve in vivo direct conversion of toxic glial cells into target motor neurons in the spinal cord. Pericyte transplantation extended the survival of SOD1 mice and iPSC-derived neuronal cells by expression of anti-oxidant enzymes, 103 implying that cell replacement could revert the protective effects on motor neurons. Pericyte degeneration precedes the disease progression not only in ALS and but also in AD. 104 Pericyte loss slows the blood flow and affects the accumulation of Aβ, 105 which in turn induces pericyte loss, further burdening the Aβ load, 106 suggesting that in vivo direct conversion of adjunct astrocytes beside the BBB into functional pericytes could contribute to efficient remodeling of NVU to prevent AD pathological progression.
A three-dimensional (3D) cultured system of the CNS microenvironment could provide an attractive research platform for reprogramming technology.107,108 A scaffold-based 3D system of neuronal cells derived from iPSCs has promoted the AD pathological recapitulation of Aβ. 109 In a scaffold-free system, a 3D spheroid structure derived from AD-iPSCs can produce extracellular Aβ, replicating AD phenotypes of the cellular niche in vivo. 110 These evolving in vitro modellings using a 3D cultured system could contribute to the development of in vivo direct reprogramming technology applicable for neurodegenerative diseases.
Conclusion and future direction
Collectively, rapid advances and growing evidence of stem cell technology for cell regeneration and CNS tissue repair have been accumulating. TFs-mediated direct reprogramming into functional neurons and other types of neural cells could be a promising approach for early remodeling of NVU in both acute CNS injury and neurodegenerative diseases.
However, there are several limitations. First, the delivery method for TFs must be considered for targeted cell conversion into intended functional cells. Careful selection of target cells as a source of reprogramming is needed because locally delivered TFs can affect the cells surrounding the target cells and rather ruin the tissue remodeling by functional endogenous CNS cells. Therefore, cell heterogeneity in the intended tissue remodeling would be a primary hurdle for in vivo direct reprogramming. One resolution is a local injection of viral vectors carrying the promoter specifically expressed in target cells, which would be effective for minimizing the off-target effect. Adeno-associated virus vector has been reported to be relatively safe and applicable to gene therapy for CNS diseases. 111 Intravenous delivery of miRNA/mRNA enwrapped by exosome via leaky BBB reprogramming would be another attractive approach aimed at remodeling the micro-circulating system including pericyte assembly in acute CNS injury and neurodegeneration. 112
Second, the safety and functional role of reprogrammed cells should be clarified and their tracking and monitoring must be addressed. Reprogramming is mainly aimed at (1) compensation for loss of function of neurons secreting neurotransmitters to receiver cells by cell replacement, (2) rebuilding the neuronal circuit to convey bioelectrical signaling via synapse formation, (3) and remodeling early assembly among neurons, glial cells and endothelial cells by improving the microcirculation and metabolic homeostasis. Ideally, reprogrammed cells should maintain cellular specialization, survive for a long period, and form a synaptic connection with the correct target, but these ‘three S (Specialization, Survival, Synapse)’ are all challenging. To address the therapeutic effect of in vivo direct reprogramming, we need to define appropriate biomarkers for neurotransmitters, functional imaging for axonal connectivity, and cerebral blood/CSF flow for cerebral perfusion in the patient after in vivo direct reprogramming.
Third, direct reprogramming is insufficient for the radical treatment of diseases resulting from genetic mutation. Although replenishment by regenerative neurons could be transiently effective for the loss of function in neurodegenerative diseases, the disease-causing mutation carried in reprogrammed cells will cause gradual neuronal cell loss by its gain of toxic function. One promising solution may be the combinational delivery of both additional genes for functional restoration and TFs for direct reprogramming in vivo. 113 Technological progress in the drug-delivery system, imaging and gene editing will boost in vivo direct reprogramming to become future promising regenerative medicine.
Fourth, reprogramming could induce immature iNs that have the potential to transdifferentiate into other types of cells than the aimed-for cells. For example, Ascl1 is sufficient to induce a heterogenic cell population including neuron progenitor cells. 23 Further, elucidation of hierarchical mechanisms for both in vivo and in vitro reprogramming would be necessary for efficient and safe regenerative medicine. 70 In vivo direct conversion into GABAergic, glutamatergic and dopaminergic neurons can be achieved, although the precise mechanism of conversion into other types of neurons (i.e. cholinergic, serotoninergic, sensory) has not yet been elucidated. Utilization of computational strategy for predicting lineage specifiers with single cell analysis might facilitate our understanding of the complex transcriptional network to manipulate cell identity for in vivo direct reprogramming.114,115
Footnotes
Acknowledgements
We would like to express our sincere gratitude to all our coworkers and collaborators: Kayoko Tsukita, Takako Enami, Ayako Nagahashi, Ikuyo Inoue, Ran Shibukawa, Yukako Sagara, Yasue Okanishi, and Kazuma Kamata for technical assistance; Mikie Iijima, Nozomi Kawabata, Ayumi Suzuki, Miwa Fujita, Mayu Okuda, and Makiko Yasui for valuable administrative support.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded in part by a grant for Core Center for iPS Cell Research of Research Center Network for Realization of Regenerative Medicine from AMED to H.I. and from KAKENHI (18K15347, 20K07883) to N.E. and KAKENHI (18K18452, 18H02717) to H.I. and supported in part by NIH to K.A.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.