
Abstract
The role of the mitochondrial calcium uniporter (MCU) gene (Mcu) in cellular energy homeostasis and generation of electrical brain rhythms is widely unknown. We investigated this issue in mice and rats using Mcu-knockout and -knockdown strategies in vivo and in situ and determined the effects of these genetic manipulations on hippocampal gamma oscillations (30–70 Hz) and sharp wave-ripples. These physiological network states require precise neurotransmission between pyramidal cells and inhibitory interneurons, support spike-timing and synaptic plasticity and are associated with perception, attention and memory. Absence of the MCU resulted in (i) gamma oscillations with decreased power (by >40%) and lower synchrony, including less precise neural action potential generation (‘spiking'), (ii) sharp waves with decreased incidence (by about 22%) and decreased fast ripple frequency (by about 3%) and (iii) lack of activity-dependent pyruvate dehydrogenase dephosphorylation. However, compensatory adaptation in gene expression related to mitochondrial function and glucose metabolism was not detected. These data suggest that the neuronal MCU is crucial for the generation of network rhythms, most likely by influences on oxidative phosphorylation and perhaps by controlling cytoplasmic Ca2+ homeostasis. This work contributes to an increased understanding of mitochondrial Ca2+ uptake in cortical information processing underlying cognition and behaviour.
Introduction
The integrative role of mitochondrial Ca2+ uptake and cycling in central neurons has been established on molecular, cellular and system levels.1–3 This role comprises a variety of physiological processes, such as maintenance of intracellular Ca2+ homeostasis, synaptic activity, vesicle exocytosis and energy metabolism.4–10 During neurometabolic coupling, mitochondrial Ca2+ can act as a feed-forward regulator of oxidative phosphorylation and ATP production in mitochondria by affecting the activity of pyruvate dehydrogenase (PDH), several TCA cycle enzymes and F1FO-ATP synthase.1,11,12 Moreover, the detrimental consequences of excessive mitochondrial Ca2+ uptake in neurons have been studied under various pathophysiological conditions.1,13,14 Excessive mitochondrial Ca2+ uptake can trigger several harmful events, including opening of the mitochondrial permeability transition pore and loss of mitochondrial membrane potential, which potentially lead to bioenergetic failure and neuronal death.15–19 Both initiation and cellular consequences of excessive mitochondrial Ca2+ uptake likely differ in ischemia, epilepsy and age-related neurodegenerative diseases, however.20–23
During neuronal activity, cytoplasmic Ca2+ transients are associated with the uptake of Ca2+ into mitochondria and increased mitochondrial matrix Ca2+ concentration.2,6,8,24,25 Importantly, mitochondrial Ca2+ uptake requires the recently identified mitochondrial Ca2+ uniporter (MCU),26,27 which is part of a large protein complex.2,28 Alternative routes for mitochondrial Ca2+ uptake, such as ion exchangers and the rapid uptake mode, have been also described.2,29,30
The role of the MCU in the cellular homeostasis underlying the generation of fast neuronal network rhythms that associate with cortical information processing in vivo is unknown. This role is of particular interest because the awake human brain features per se a high metabolic rate that further increases during functional activation of primary cortical regions, with mean values of 27 ± 10% and 20 ± 13% for ΔCMRglc and ΔCMRO2, respectively.31–33 Therefore, active neurons critically depend on efficient neurometabolic coupling and high performance of mitochondrial oxidative phosphorylation.22,31 In theory, such neurometabolic coupling can be achieved both by cytosolic Ca2+ that stimulates mitochondrial substrate carriers and by mitochondrial matrix Ca2+ that stimulates TCA cycle dehydrogenases. However, whether mitochondrial Ca2+-uptake via the MCU complex is required to support physiological neuronal network activity by regulating cellular energy metabolism and/or Ca2+ homeostasis is widely unknown. 2
We started to address this by studying gamma oscillations (30–70 Hz) and sharp wave-ripples in hippocampal slice preparations.34–37 These physiological network rhythms require precise synaptic transmission between excitatory pyramidal cells and inhibitory interneurons and occur during wakefulness and sleep.38,39 They support, for example, spike-timing and synaptic plasticity on the cellular level and are thus thought to contribute to the emergence of higher brain functions, such as perception, attention and memory.38–40 We used various knockout (KO) and knockdown (KD) strategies of the Mcu in vivo and in situ.
Materials and methods
Mcu-knockout and Mcu-knockdown strategies
All animal experiments conformed to national and institutional guidelines including the Animals [Scientific Procedures Act] 1986 (UK), and the Council Directive 2010/63/EU of the European Parliament, and had full Home Office ethical approval (University of Edinburgh Animal Welfare and Ethical Review Body; University of Heidelberg Animal Welfare Office and Regierungspraesidium Karlsruhe, licenses T46/14 and T96/15). All animals were maintained in pathogen-free and light- (12-h light/12-h dark) and temperature-controlled (22°C ± 2°C) conditions. Food (LasVendi Rod 16 or Rod 18) and water were available ad libitum. Animals were group-housed in conventional cages with ABBEDD LT-E-001 bedding material and were provided with environmental enrichment. Animal welfare was assessed daily by staff of the animal facility. Experiments were performed and reported in accordance with the ARRIVE guidelines. The investigator was blinded for the genotype.
To obtain mice with brain-specific loss of Mcu expression, Mcufl/fl mice (colony name: B6N-Atm1Brd-Mcutm1c(EUCOMM)Hmgu/H MGI Allele name: Mcutm1c(EUCOMM)Hmgu, MGI ID: 5692853, EMMA ID: EM:10448) were crossed with C57BL/6 Emx1-Cre mice
41
in which the neo cassette had been removed by crossing with the ROSA26Fki Flp deleter mice.
42
Mcufl/fl mice were generated by MRC Harwell from targeted ES cells made by The European Conditional Mouse Mutagenesis Program, as part of the International Mouse Phenotyping Program. Intercrosses of Mcu/Cre+ double heterozygotes with heterozygous Mcu/Cre− mice were carried out to generate experimental Mcu knockout (Mcu/Cre+) and wildtype controls (Wt/Cre+). Animals were genotyped by PCR using the following primers (5ʹ-3ʹ): A:
Recombinant adeno-associated viral vectors for the expression of short hairpin RNAs (shRNAs) contain a ubiquitous U6 promoter for shRNA expression and a CaMKII promoter driving mCherry expression. Note that due to the use of a CaMKII promoter, the fluorescent infection marker is expressed in glutamatergic neurons only, while shRNAs are expressed in all infected cells. To verify that serotype 1/2 efficiently targets parvalbumin-expressing GABAergic basket cells, we infected slice cultures with a rAAV that expresses mCherry under the control of a ubiquitous CMV promoter. Immunostaining for parvalbumin confirmed reliable infection of basket cells (Suppl. Figure 2). The following shRNA target sequences were used in this study (5′–3′): control shRNA targeting LacZ:
Slice preparations
Wild type and Mcu-KO male mice (6–12 weeks old) were anaesthetised with CO2 and decapitated. The brain was quickly removed and maintained in cooled (4°C) artificial cerebrospinal fluid (ACSF), saturated with 95% O2 and 5% CO2. After removal of frontal brain structures and the cerebellum, horizontal entorhinal-hippocampal slices (400 µm) were prepared using a vibratome (VT 1000 s, Leica, Bensheim, Germany). These acute slices were stored in a Haas-type interface chamber at 34 ± 1°C for recovery of at least 2 h as well as for experimental recordings. Slices with incomplete entorhinal-hippocampal structures were excluded.
Slice cultures were prepared as described.36,44 In brief, hippocampal slices (400 µm) were cut with a McIlwain tissue chopper (Mickle Laboratory Engineering Company Ltd., Guildford, UK) from 10-day-old male Wistar rats under sterile conditions. Slices with intact hippocampal structures were maintained on Biopore™ membranes (Millicell standing inserts, Merck Millipore, Darmstadt, Germany) between culture medium, which consisted of 50% minimal essential medium, 25% Hank’s balanced salt solution (Sigma-Aldrich, Taufkirchen, Germany), 25% heat-inactivated horse serum (Life Technologies, Darmstadt, Germany), and 2 mM L-glutamine (Life Technologies) at pH 7.3 titrated with Trisbase, and humidified normal atmosphere (5% CO2, 36.5°C) in an incubator (Heracell, Thermoscientific, Dreieich, Germany). Slices were randomly assigned to experimental groups. The culture medium (1 ml) was replaced three times per week. Slice cultures were used after 7–14 days in vitro (DIV), when the tissue had recovered from slice preparation and damaged cut surfaces had been re-organized.1,45
Recording solutions and drugs
Acute slices and slice cultures were stored in a Haas-type interface chamber and constantly supplied with heated (34 ± 1°C) recording solution, i.e. ACSF containing: 129 mM NaCl, 3 mM KCl, 1.25 mM NaH2PO4, 1.8 mM MgSO4, 1.6 mM CaCl2, 26 mM NaHCO3, and 10 mM glucose.36,44 The pH was 7.3 when the recording solution was saturated with 95% O2 and 5% CO2. The gas supply to the interface chamber was 1.5 l/min (95% O2 and 5% CO2).
Gamma oscillations were induced by bath application of acetylcholine (2 µM) and physostigmine (400 nM) in rat slice cultures or the muscarinic receptor agonist carbachol (5 µM) in mouse acute slices.34,36,45,46
Carbachol and physostigmine were purchased from Tocris (R&D Systems GmbH, Wiesbaden-Nordenstadt, Germany). Standard salts and acetylcholine were from Sigma-Aldrich (Taufkirchen, Germany).
Recordings of local field potentials
In rat slice cultures, the local field potential was recorded with glass electrodes (tip diameter 3–5 µm) that were pulled from GB150F-8P borosilicate filaments (Science Products GmbH, Hofheim, Germany) with a PC-10 vertical micropipette puller (Narishige International Ltd., London, UK) and backfilled with ACSF. In mouse acute slices, carbon fibre electrodes (Kation Scientific, Minneapolis, MN, USA) were used. The microelectrodes were positioned in stratum pyramidale of the CA3 region with a mechanical micromanipulator (MX-4, Narishige). Extracellular field potentials were low-pass filtered at 3 kHz, and digitised at 10 kHz using CED 1401 interface and processed with Spike2 software (Cambridge Electronic Design, Cambridge, UK) for offline analysis.
Offline signal analysis of 5-min data segments from local field potential recordings was performed using custom-made scripts in MATLAB 2015a (The MathWorks, Inc., Natick, MA, USA). Recordings of gamma oscillations were low-pass filtered with a digital Butterworth algorithm at 200 Hz cutoff frequency and processed with Welch’s algorithm and fast Fourier transform with a Hamming window size of 32,768 points for calculation of the power spectral density (power) (bin size of 0.3052 Hz). Gamma oscillations were analysed for peak power, frequency and full width at half-maximum (FWHM). Recordings of sharp wave-ripples were band-pass filtered with a Butterworth algorithm between 5 and 60 Hz corner frequencies to detect the transient sharp wave component. Amplitude and incidence of sharp waves were measured and averaged. Ripples superimposed on the sharp wave were analysed using continuous wavelet transformation. Multi-unit activity was assessed by high-pass filtering of the local field potential with a Butterworth algorithm at 700 Hz corner frequency. The threshold for unit detection was set to 4.5 standard deviations of the local field potential during spontaneous asynchronous activity and the correctness of unit detection was checked visually.
Toluidine blue staining and immunohistochemistry
For toluidine blue staining (Sigma-Aldrich), acute hippocampal slices were fixed overnight with 4% paraformaldehyde in phosphate-buffered salt solution (PBS), incubated for 5 h in 30% sucrose (AppliChem GmbH, Darmstadt, Germany) and cut in thin sections (40 µm) with a CM1850 cryostat (Leica Microsystems GmbH, Nussloch, Germany). Sections were mounted on slides, exposed to descending ethanol series, briefly rinsed in double-distilled water and then incubated in 0.1% toluidine blue working solution (pH 2.3) for 1–5 min. Thereafter, the sections were briefly rinsed in double-distilled water; 95% ethanol with traces of glacial acetic acid was used for colour differentiation of the staining. The sections were then exposed to ascending ethanol series, a 1:1 mixture of 100% ethanol and xylene, and finally xylene for 6–10 min. Afterwards the sections were embedded with Entellan®Neu (Merck Millipore, Schwalbach, Germany).
For eGFP, parvalbumin and glial fibrillary acidic protein (GFAP) staining, mice were anaesthetized by intraperitoneal injection of sodium pentobarbital and transcardially perfused with 20 ml of phosphate buffer (PB) followed by 20 ml of ice cold 4% paraformaldehyde. A vibratome (Leica, UK) was used to cut 50 µm thick sections, which were stored free-floating in PB at 4°C until use. Sections were washed in PB, blocked with 5% donkey serum for 1 h and incubated in the following antibodies overnight: anti-GFP conjugated to FITC (1:500 Abcam ab6662), mouse anti-parvalbumin (1:5000, Swant PV235) and rabbit anti-GFAP (1:1000, Dako Z0334). Sections were then washed in PB and incubated in donkey anti-mouse Alexa Fluor 546 antibody (1:500) and donkey anti-rabbit Alexa Fluor 546 antibody (1:500) for 1 h before being washed and mounted onto slides. Sections were imaged using a Leica AF6000LX immunofluorescence microscope.
Western blotting
Wild type, Mcu/Cre− and Mcu-KO mice were anaesthetized with isoflurane and decapitated. Brains were quickly removed, the hippocampus dissected and the CA3 region microdissected using a scalpel blade. The tissue was snap frozen and stored at −80°C until required. Samples were homogenized in RIPA buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 0.5% sodium deoxycholate) containing protease and phosphatase inhibitors (Roche, UK). The samples were then lysed in 1.5× LDS sample buffer (NuPage, Life Technologies) and boiled at 100°C for 10 min. Approximately 10 µg of protein was loaded onto a precast gradient gel (4–16%; Invitrogen) and subjected to electrophoresis. Western blotting onto a PVDF membrane was then performed using the Xcell Surelock system (Invitrogen) according to the manufacturer's instructions. Following the protein transfer, the PVDF membranes were blocked for 1 h at room temperature with 5% (w/v) non-fat dried milk in TBS with 0.1% Tween-20. The membranes were incubated at 4°C overnight with the primary antibodies diluted in blocking solution: Rabbit anti-MCU (1:4000, Sigma HPA016480) and rabbit anti-beta actin (1:50,000, Abcam ab8227). For visualisation of Western blots, HRP-based secondary antibodies were used followed by chemiluminescent detection on Kodak X-Omat film. Western blots were digitally scanned and densitometric analysis was performed using Image J.
Gene expression analysis
Wild type and Mcu-KO mice (6–12 weeks old) were anaesthetised with CO2 and decapitated. Hippocampal CA3 region was rapidly dissected and flash frozen in liquid nitrogen. RNA was isolated from frozen tissue using an RNeasy Microarray Tissue Mini kit (Qiagen, Hilden, Germany) with additional on-column DNase I digestion according to the manufacturer’s instructions; 460 ng RNA per sample were used to prepare cDNA using a RT2 First Strand Kit (Qiagen) according to the manufacturer’s instructions. Real-time PCR was performed on an ABI StepOnePlus system using RT2 SYBR® Green ROX™ qPCR Mastermix and RT2 Profiler PCR Arrays PAMM-087ZA (mouse mitochondria) and PAMM-006ZA (mouse glucose metabolism). Expression of target genes was normalized against the geometric mean of Actb, Gapdh, and Gusb as endogenous control genes. Heatmaps (Figure 7) were generated with Heatmapper. 47
pPDH analysis
Acute slices were recorded in standard ACSF in a recording chamber, or were treated with carbachol in a recording chamber for 40 min. Slices were then fixed overnight with 4% paraformaldehyde at 4°C. After keeping the slices in 30% sucrose (AppliChem, Darmstadt, Germany) they were cut into 50 µm sections on a cryostat (CM1850, Leica Microsystems, Wetzlar, Germany). Free floating sections were blocked and permeabilised with 5% normal goat serum and 0.3% Triton X-100 for 90 min at RT, then labelled overnight at 4°C with a mixture of primary antibodies (mouse anti-PDH E1-alpha, Life Technologies 456600, lot 456600/G0529, 1:500; rabbit anti-pPDH Ser293, Millipore ABS204, lot 2315725, 1:2000), and then labelled over night at 4°C with a mixture of secondary antibodies (Alexa 594-labelled goat anti-mouse IgG, Thermo Fisher Scientific A-11005, and Alexa 488-labelled donkey anti-rabbit IgG, Thermo Fisher Scientific R37118; both 1:1000). Sections were counterstained with Hoechst 33258 (Sigma-Aldrich, 1 µg/ml), and mounted in Mowiol 4-88 (Calbiochem).
Images were recorded in sequential mode on a Leica TCS SP8 confocal microscope equipped with a Leica HC PL APO CS2 63× NA 1.4 oil-immersion objective (pixel size 240 nm × 240 nm). Images were imported into FIJI 48 and converted to 32-bit; 488 nm and 552 nm excitation channels were split and a threshold was set in both channels to contrast mitochondria. The 488 nm excitation channel (pPDH) was divided by the 552 nm excitation channel (total PDH). The resulting 488 nm/552 nm ratio image was displayed in fire lookup table. The 488 nm/552 nm ratio was measured in stratum pyramidale (as identified in the Hoechst channel) and data were exported to Microsoft Excel. Image acquisition and data analysis were done blinded to genotype.
Calculations and statistics
Electrophysiological data are presented as median with 25th and 75th percentile derived from ‘n' acute slices (cultured slices) from ‘N' mice (rats), unless stated otherwise. Statistical significance, P < 0.05 or confidence interval (CI) was determined using SigmaPlot 12.5 (Systat Software, San Jose, CA) and GraphPad Prism (GraphPad Software Inc., La Jolla, USA). Data distribution was checked for normality with Shapiro–Wilk test. Comparisons among paired data were made with Wilcoxon signed-rank test. Comparisons among unpaired data were made with Kruskal–Wallis ANOVA on ranks followed by Dunn's post hoc test. Figures were generated with MATLAB 2015a (MathWorks), and CorelDRAW (Corel Corporation, Ottawa, Ontario, Canada). Quantitative immunofluorescence data are presented as mean + 95% CI. Data distribution was checked for normality with Shapiro–Wilk test and P value was determined by unpaired two-tailed Student’s t-test using GraphPad Prism (GraphPad Software Inc., La Jolla, USA).
Results
We used a combined experimental approach to determine the role of the MCU in the generation of fast neuronal network rhythms in the hippocampus of mice and rats.
First, we used acute hippocampal slices from conditional Mcu-KO mice. These mice were generated by crossing a floxed Mcu mouse line to Emx1-Cre mice that express Cre-recombinase in cortical glutamatergic neurons. 41 Notably, in these mice, the MCU is still present in GABAergic interneurons, such as parvalbumin-positive, fast-spiking inhibitory interneurons important for the generation of gamma oscillations and sharp wave-ripples.34,41,49,50 The Mcu-KO mice showed no gross differences in the hippocampal cytoarchitecture (Figure 1(a) and (b)). Western blotting of extracts from the hippocampal CA3 region confirmed significant knockout of MCU (Figure 1(c) and (d)) as compared to wild type and Mcu/Cre− mice. To confirm the distribution and identity of Emx1-Cre expressing cells, Emx1-Cre mice were crossed with B6;129S4-Gt(ROSA)26Sortm9(EGFP/Rpl10a)Amc/J mice, in which Cre-mediated excision results in eGFP expression. This revealed expression in neurons throughout the cortex and hippocampus, consistent with previous studies 41 (Suppl. Figure 1(a) and (b)). Immunohistochemistry confirmed the lack of GFAP (astrocyte marker) and parvalbumin (interneuron marker) co-expression with eGFP (Suppl. Figure 1(c) to (j)).
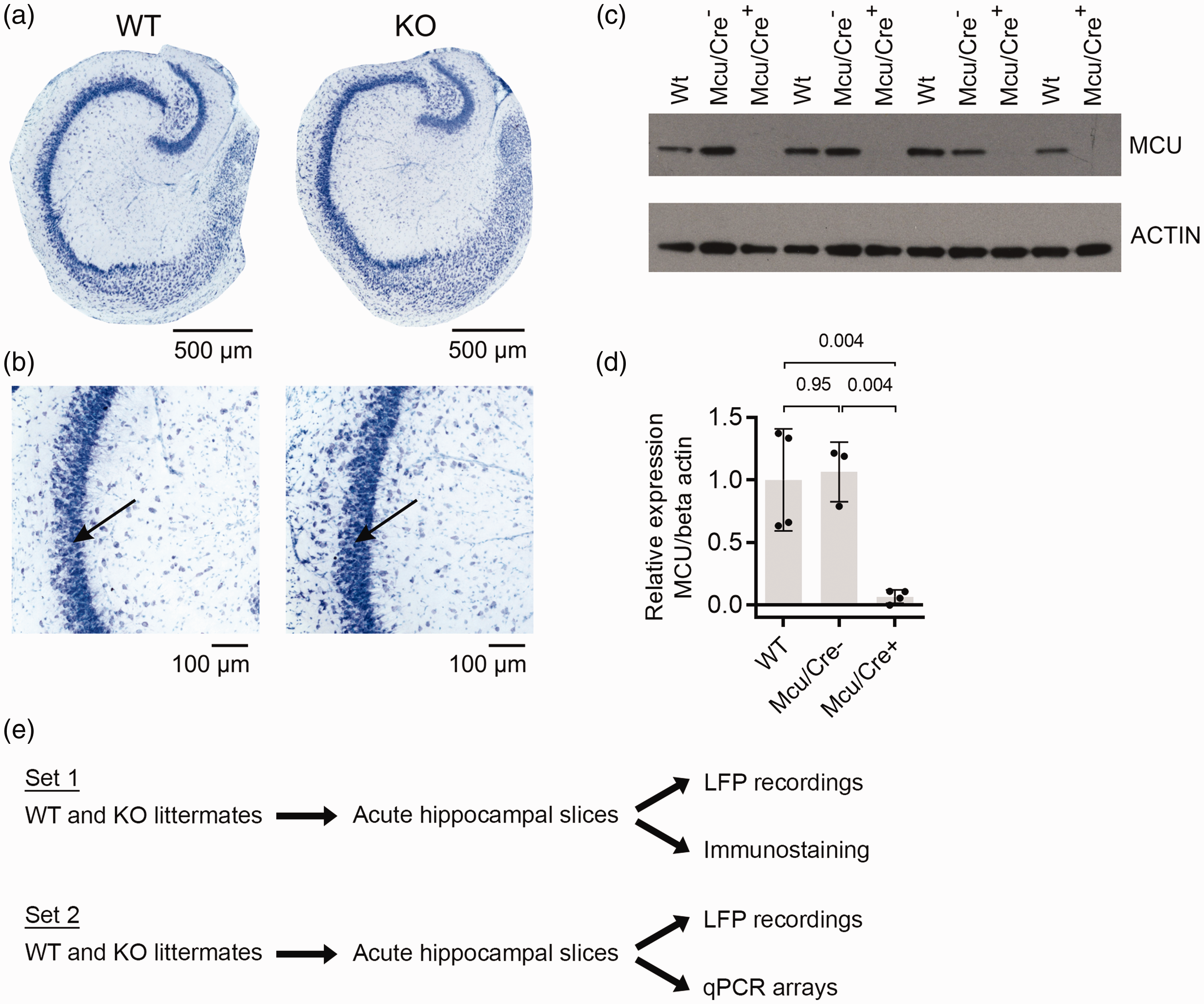
Cytoarchitecture of the hippocampus in wild type (WT) and Mcu-KO (KO) mice. (a) Acute hippocampal slices were stained with toluidine blue (TB) in WT and KO mice. (b) The CA3 region is shown at higher magnification. Note that the cytoarchitecture of the hippocampus is well preserved. The extracellular local field potential electrode was positioned in stratum pyramidale (strongly stained layer with densely packed neuronal somas; black arrows). (c) Immunoblot analysis of microdissected hippocampal CA3 region confirms lack of MCU protein expression in Mcu/Cre+ mice. Blots have been cropped for conciseness. (d) Quantification of immunoblot experiments. Round symbols represent individual animals, bars represent mean, error bars indicate standard deviation. P-values are indicated in the graph and were determined by one-way ANOVA followed by Tukey’s multiple comparisons test. (e) Scheme of the experimental design in mice. WT (Wt/Cre+) and Mcu-KO (Mcu/Cre+) littermates were used in each experiment. Acute (ex vivo) hippocampal slices were used for electrophysiological local field potential (LFP) recordings, immunostaining or qPCR arrays to obtain various readouts. The experimental design for WT and Mcu-KD slice cultures of the rat was similar.
As a second approach, we used organotypic hippocamal slice cultures from rats.36,44 In these slice cultures, Mcu was knocked down by recombinant adeno-associated virus-mediated expression of Mcu-directed short hairpin RNAs (shRNAs) in excitatory and inhibitory neurons 18 (Suppl. Figure 2).
In these slice preparations (Figure 1(e)), we performed local field potential recordings, including fast Fourier transform, to characterize the properties of cholinergically induced, persistent gamma oscillations and spontaneously occurring, recurrent sharp wave-ripples, both of which share many features with their counterparts in vivo.36,37,46,51 We focussed on the CA3 region, which is an intrinsic generator of gamma oscillations and sharp wave-ripples in the hippocampus,34,50,51 and shows a higher Mcu-expression of about 2-fold compared with the CA1 region. 52
Disturbances of gamma oscillations
Gamma oscillations with a frequency of around 32 Hz were present in the CA3 region of wild type and Mcu-KO mice (Figure 2(a), (b) and (d)). However, Mcu-KO mice showed a strong decrease in the power of gamma oscillations by about 87% (Figure 2(c)) as well as an increase in full width at half maximum (FWHM), reflecting less precision (Figure 2(e)). We note that the oscillation power increases with number and synchrony of postsynaptic currents at gamma-frequency, whereas FWHM increases with jitter in the timing of postsynaptic currents. 37
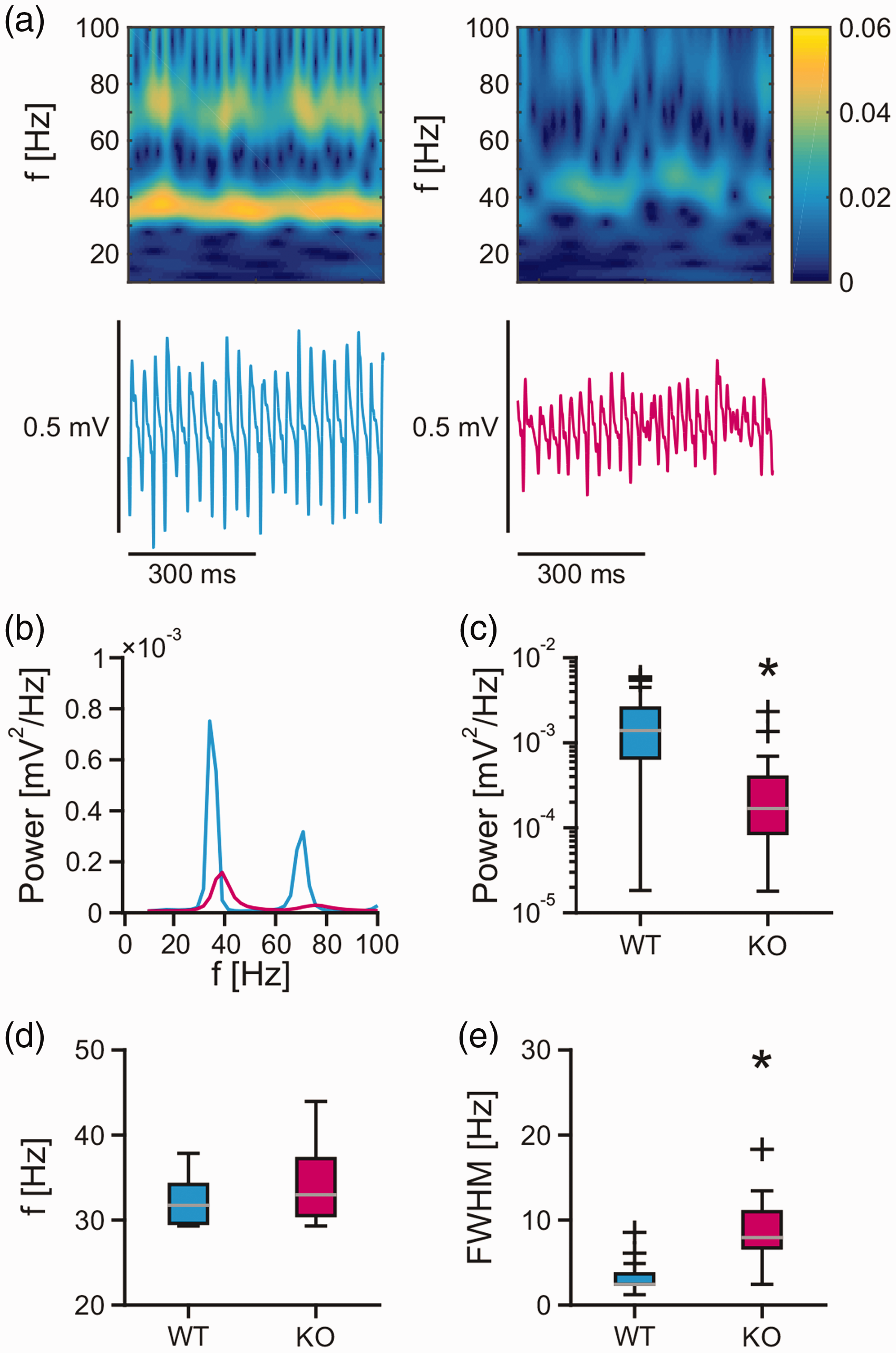
Gamma oscillations in mouse acute slices. Persistent gamma oscillations were recorded in stratum pyramidale of the CA3 region in acute hippocampal slices of the mouse. (a) Sample traces of local field potential recordings and corresponding wavelet transform of gamma oscillations in wild type (blue trace) and Mcu-KO (magenta trace) mice. (b) Sample power spectra of gamma oscillations calculated from 5 min intervals in slices from wild type (blue) and Mcu-KO (magenta) mice. Power spectra in (b) correspond to gamma oscillations in (a). Gamma oscillations in hippocampal slices from wild type (WT) (n = 23, N = 6) and Mcu-KO (n = 20, N = 6) mice were analysed for (c) the peak of the power spectrum (Power), (d) the peak frequency (f) and (e) the full width at half maximum (FWHM). Mann–Whitney rank sum test was applied for statistical analysis and Shapiro–Wilk test for normality. Statistical significance is marked by asterisks (P < 0.05).
Similar disturbances in the power of gamma oscillations were present in the CA3 region of rat slice cultures expressing shRNAs against Mcu. To obtain robust results, we used three different shRNA sequences. Expression of sh1 and sh2 resulted in a decrease in the power of gamma oscillations by about 40.9% and 65.9%, respectively (Figure 3(a) to (g)). The increase in FWHM was only present in slice cultures expressing sh2 and was accompanied by an increase in gamma oscillation frequency (Figure 3(h) and (i)). By contrast, expression of sh3 resulted in a decrease in FWHM (Figure 3(g) to (i)). Thus, we broadly confirm our findings in Mcu-KO mice with two out of three variants of shRNAs in rat slice cultures.
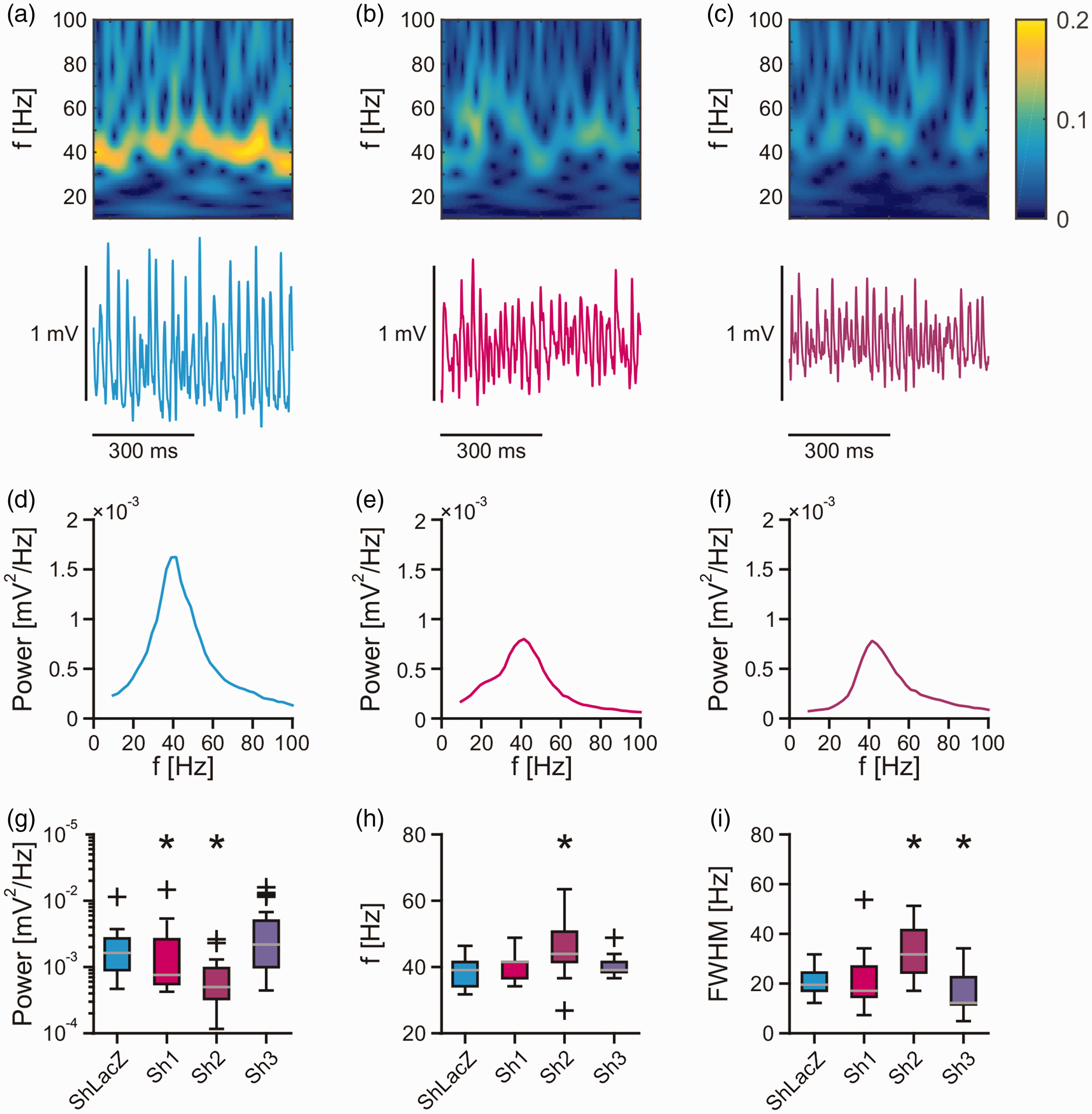
Gamma oscillations in rat slice cultures. Persistent gamma oscillations were recorded in stratum pyramidale of the CA3 region in hippocampal slice cultures of the rat. (a–c) Sample traces of local field potential recordings and corresponding wavelet transform of gamma oscillations in control (a, ShLacZ) and Mcu-knockdown (KD) slice cultures (b, Sh1; c, Sh2). (d–f) Sample power spectra of gamma oscillations calculated from 5 min intervals in control (d) and KD slice cultures (e, Sh1; f, Sh2). Gamma oscillations in control (ShLacZ, n = 26, N = 6) and KD slice cultures (Sh1, n = 25, N = 7; Sh2, n = 15, N = 4; Sh3, n = 25, N = 8) were analysed for (g) the peak of the power spectrum (Power), (h) the peak frequency (f) and (i) the full width at half maximum (FWHM) of the gamma power. (g–i) Mann–Whitney rank sum test was applied for statistical analysis and Shapiro–Wilk test for normality. Statistical significance is marked by asterisks (P < 0.05).
These findings suggest that the MCU is required for proper generation of gamma oscillations.
Disturbances of neuronal spiking synchronization
We further characterized the synchronization of neuronal action potential generation (‘spiking') during gamma oscillations. For this purpose, multi-unit activity was extracted from local field potential recordings (Figure 4(a)). In wild type mice, there was a second peak of the multi-unit intervals present at 20–30 ms, reflecting synchronized neuronal spiking at the gamma-band rhythm (Figure 4(b)). Notably, this second peak of multi-unit intervals was absent in Mcu-KO mice. In addition, the timing of multi-unit activity was less concentrated at the negative peak of the gamma-band cycle in Mcu-KO mice and thus less precise (Figure 4(c)). The frequency of multi-unit activity was similar in wild type and Mcu-KO mice (Figure 4(d)), excluding an overall decrease in neuronal excitability in the mutant.
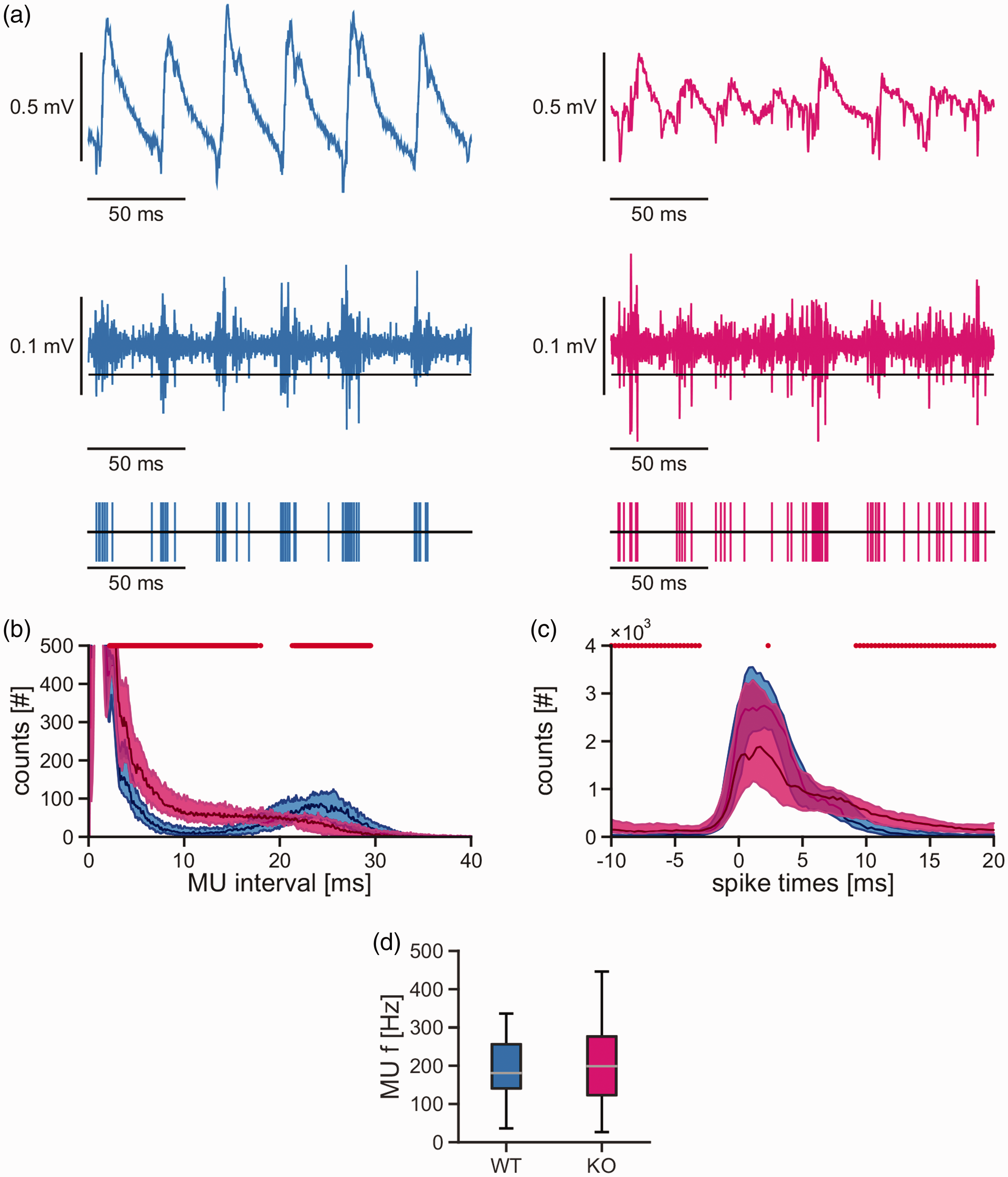
Spiking synchronization during gamma oscillations in mouse acute slices. Persistent gamma oscillations were recorded in stratum pyramidale of the CA3 region in acute hippocampal slices of the mouse. (a) Sample traces of the local field potential (top), with 700 Hz high-pass filter (middle) and extracted multi-unit (‘spiking') activity (bottom) during gamma oscillations in slices from wild type (blue traces) and Mcu-KO (magenta traces) mice. (b) Distribution of multi-unit intervals calculated from 5 min intervals in wild type (blue, 58,380 ± 3930 events, n = 23, N = 6) and Mcu-KO (magenta, 61,500 ± 7710 events, n = 20, N = 6) mice. Red dots denote significant differences. Note that the second peak of multi-unit intervals at 20–30 ms is absent in Mcu-KO (magenta). (c) Distributions of the timing of multi-unit activity relative to the negative peak of the gamma-band cycle (0 ms). Red dots denote significant differences. Note that the timing of multi-unit activity is less precise in Mcu-KO mice. (d) Frequency of multi-unit activity in wild type (WT) and Mcu-KO mice. Mann–Whitney rank sum test (b, c) and Student’s t-test (d) were applied for statistical analysis and Shapiro–Wilk test for normality. Statistical significance (P < 0.05).
These findings suggest that the MCU is required for proper synchronization of neuronal spiking during gamma oscillations.
Disturbances of sharp wave-ripples
To test whether the disturbances on the network and cellular level described above are specific for gamma oscillations, we additionally characterized the properties of sharp wave-ripples. Recurrent sharp wave-ripples were present in the CA3 region of wild type and Mcu-KO mice (Figure 5(a)). Whereas the amplitude of sharp waves was similar in wild type and Mcu-KO mice (Figure 5(b)), the incidence of sharp waves decreased by about 22% in Mcu-KO mice (Figure 5(c)). The frequency of fast ripples (Figure 5(d)) decreased by about 3%, and also the number of ripples per sharp wave (Figure 5(e)) was lower in Mcu-KO mice.
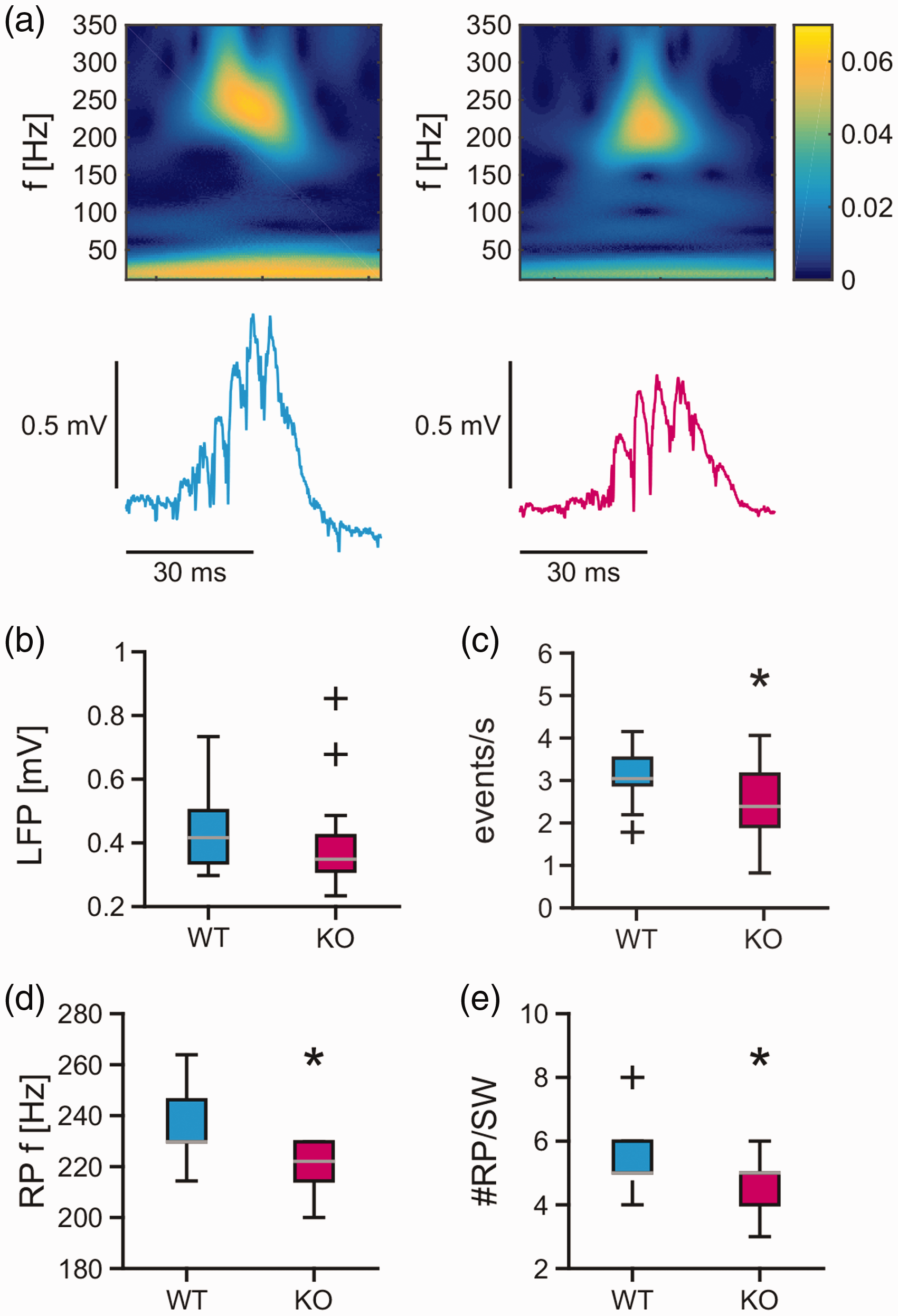
Sharp wave-ripples in mouse acute slices. Spontaneously occurring, recurrent sharp wave-ripples were recorded for 5 min in stratum pyramidale of the CA3 region in acute hippocampal slices of the mouse. (a) Sample traces of local field potential recordings and corresponding wavelet transform of single sharp wave-ripples in wild type (blue trace) and Mcu-KO (magenta trace) mice. Sharp wave-ripples in wild type (WT) (922 ± 183 events, n = 17, N = 6) and Mcu-KO (717 ± 283 events, n = 16, N = 5) mice were analysed for (b) amplitude of the sharp wave (local field potential [LFP]) and (c) incidence (events/s) of sharp waves as well as (d) the frequency of ripples (RP f) and (e) the number of ripples per sharp wave (#RP/SW). Mann–Whitney rank sum test and t-test were applied for statistical analysis and Shapiro–Wilk test for normality. Statistical significance is marked by asterisks (P < 0.05).
These findings suggest that the MCU is also required for proper generation of sharp wave-ripples.
Alterations of neurometabolic coupling
Cortical information processing depends on ATP supply by neuronal mitochondria. 22 We therefore reasoned that impaired Ca2+-mediated activation of mitochondrial oxidative metabolism might be one mechanism that underlies the disturbances in neuronal network oscillations in Mcu-KO mice. To assess neurometabolic coupling in wild type and Mcu-KO mice, we measured activity-dependent PDH dephosphorylation, which is an indicator of mitochondrial TCA cycle stimulation, 11 after 40 min of gamma oscillations in acute hippocampal slices (Figure 6). In slices from wild type mice, PDH phosphorylation was markedly reduced. In contrast, PDH phosphorylation was unchanged in slices from Mcu-KO mice.
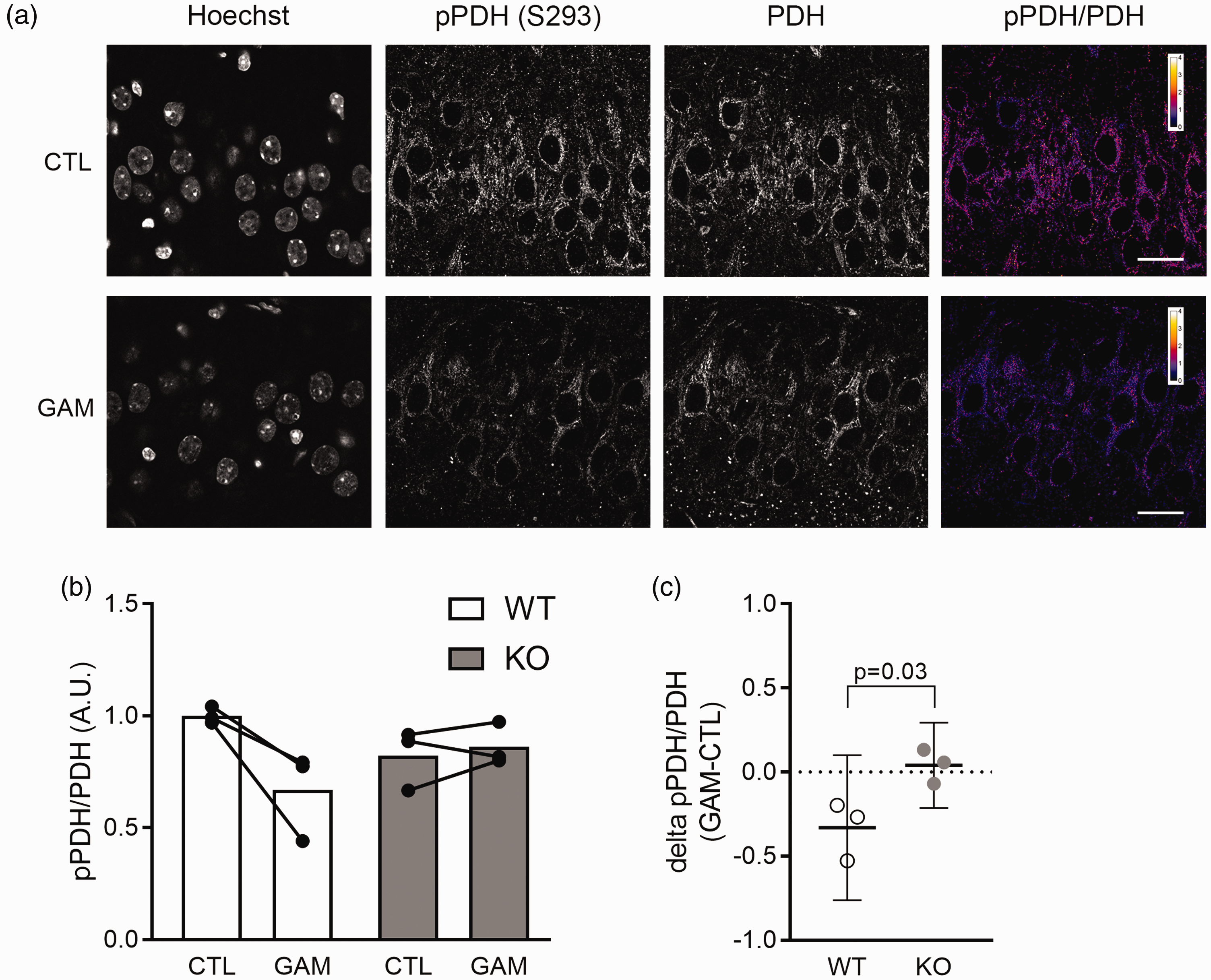
Gamma oscillation-mediated dephosphorylation of PDH in mouse acute slices. (a) Immunofluorescence labelling of acute mouse hippocampal slices. Anti-phospho PDH labelling, anti-total PDH labelling, and a pPDH/PDH ratio image are shown for a control slice (CTL) and for a slice that underwent 40 min of gamma oscillations (GAM). Both slices are from the same wild type mouse. Scale bars represent 20 µm. (b) Quantification of pPDH/PDH ratio in control slices (CTL) and slices that underwent 40 min of gamma oscillations (GAM). Slices from three animals were analysed for both conditions in each genotype. Round symbols represent individual animals; bars represent mean ratio. Values were normalized to average ratio in wild type control slices. (c) Quantification of gamma oscillation-mediated PDH dephosphorylation in wild type and Mcu-KO acute slices. Round symbols represent individual animals, horizontal lines represent mean difference between control and gamma oscillations, error bars indicate 95% CI. P value was determined by unpaired two-tailed Student’s t-test.
These findings indicate that during gamma oscillations the Ca2+-dependent stimulation of the mitochondrial TCA cycle is dysfunctional in Mcu-KO mice.
Absence of adaptations in gene expression
We next asked if the reduced capability to activate mitochondrial oxidative phosphorylation in Mcu-KO mice results in compensatory changes in the expression of metabolic genes. For this purpose, we isolated the CA3 region from hippocampus of control and Mcu-KO mice, and analysed mRNA expression of a panel of 168 genes that are related to mitochondrial function or glucose metabolism using RT2 Profiler PCR Arrays (Figure 7, Suppl. Tables 1 and 2). Using a threshold of 2-fold up- or downregulation to identify differentially expressed genes, we found no difference between WT and KO mice in any of the analysed genes.
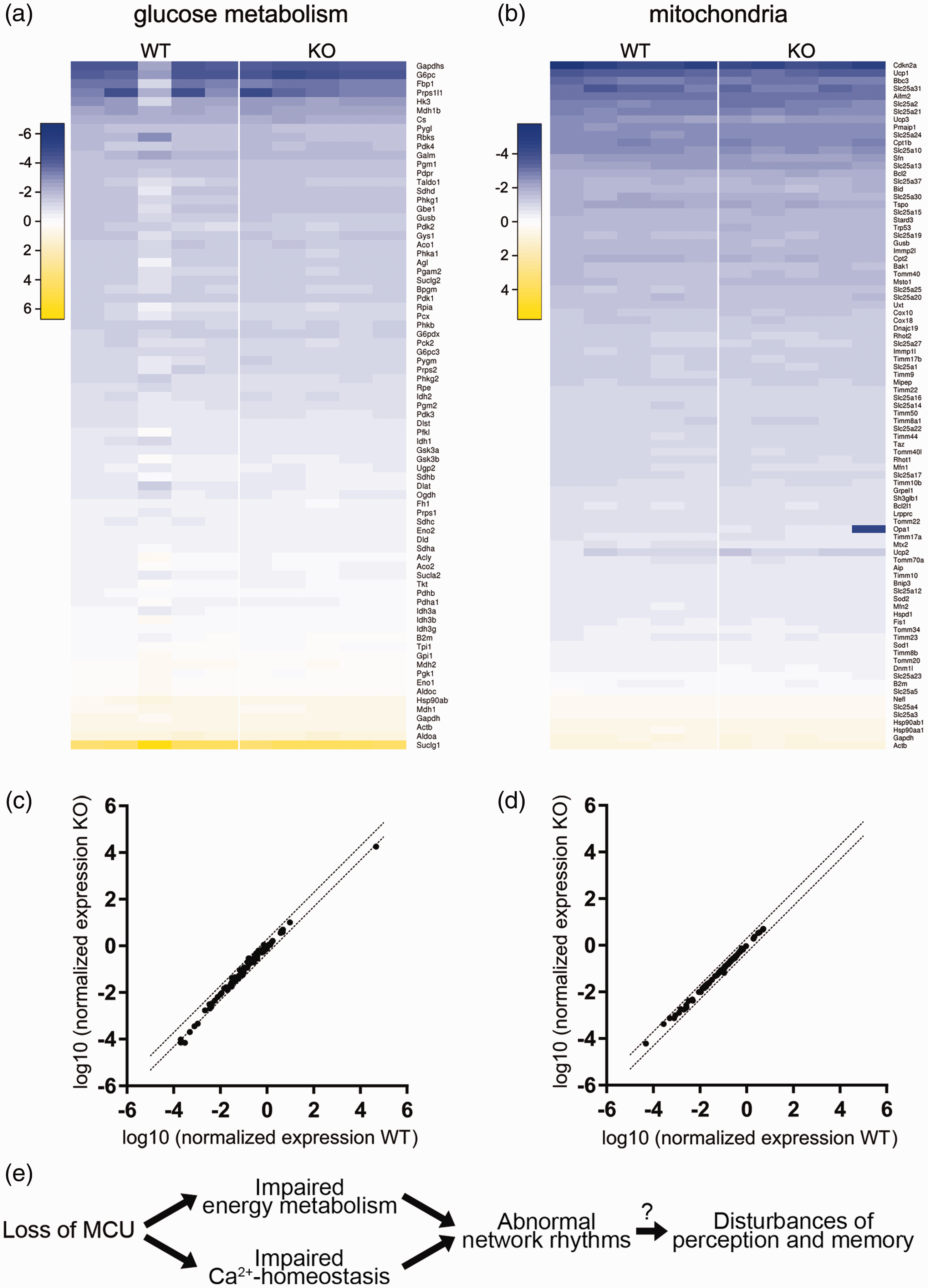
Expression of genes related to glucose metabolism and mitochondria in the mouse hippocampal region CA3 and summary scheme. (a, b) Heatmaps illustrating the results of RT2 Profiler PCR Array gene expression analyses of glucose metabolism-related (a) and mitochondria-related (b) genes. N = 5 mice per genotype. Colour scale represents log10 of normalized expression (2−dCT). (c, d) Comparison of average expression per gene in wild type versus Mcu-KO mice for glucose metabolism-related (c) and mitochondria-related (d) genes. Dotted lines indicate 2-fold up- or down-regulation. (e) Summary of the main findings and potential pathophysiological mechanisms triggered by loss of the neuronal MCU. The relative contributions of impaired oxidative energy metabolism and impaired intracellular Ca2+ homeostasis to different abnormal neuronal network rhythms as well as the disturbances of higher brain functions need to be investigated in future studies. For details, see main text.
These findings suggest that under basal conditions there are no compensatory adaptations in energy metabolism in Mcu-KO mice, at least at the transcriptional level.
Discussion
Mcu-KO and Mcu-KD strategies
In this study, we used two complementary approaches to investigate the role of the MCU during fast neuronal network rhythms. First, we generated a brain-specific Mcu-KO mouse line that lacks expression of the MCU in excitatory cortical and hippocampal neurons. The specific advantage of this approach is the complete lack of MCU expression in targeted neurons, and normal expression elsewhere. However, Emx1 promoter-driven expression of Cre recombinase starts early during development, which potentially allows for compensatory adaptations to occur. In addition, the Emx1 promoter does not drive expression of Cre recombinase in parvalbumin-positive GABAergic interneurons that are important for the generation of fast network oscillations. In a second approach, we therefore used recombinant adeno-associated viruses of serotype 1/2 to express shRNAs against Mcu in both excitatory and inhibitory neurons in rat organotypic hippocampal slice cultures (Suppl. Figure 2). Compared to acute hippocampal slices, the use of organotypic cultures provides the advantage of better tissue recovery from brain isolation and slice preparation. In addition, the transient knock-down of Mcu, albeit less efficient than the genetic knock-out, provides less time for compensatory cellular adaptions to develop. Although all three shRNAs were able to reduce MCU protein levels in rat neurons (Suppl. Figure 3), only two shRNAs were able to phenocopy the mouse knock-out. The reason for the different results obtained with the third shRNA is not clear, but might involve unidentified off-target effects of the third shRNA. However, this observation is not uncommon and, in fact, underscores the importance of using multiple shRNA sequences and additional complementary approaches. 53 Together, using ex vivo slices and slice cultures from the mouse and the rat hippocampus, we conclude that the MCU is required for neurometabolic coupling and generation of proper fast neuronal network rhythms.
Disturbances of gamma oscillations and sharp wave-ripples
The role of the MCU complex in neurometabolic coupling has been addressed in a number of previous experimental studies. These studies, however, focused on widely undefined activation stages of cultured neurons using artificial stimuli 54 or on intense metabolic stress during hypoxic-ischemic brain injury. 55
Here, we used gamma oscillations and sharp wave-ripples as models of physiological neuronal network rhythms. Gamma oscillations emerge in many cortical areas in awake mammals, including humans, and have been associated with sensory perception, attentional selection, motor activity and memory formation.22,39 Sharp wave-ripples arise in the hippocampus during waking immobility, consummatory behaviour and slow-wave sleep. They assist in transferring compressed hippocampal information to distributed neocortical circuits to support memory consolidation.38,39 In addition, sharp wave-ripples have been implicated in erasure of hippocampal memory traces and certain aspects of active spatial navigation.38–40
Gamma oscillations and sharp wave-ripples rely on the precise synaptic transmission between excitatory pyramidal cells and GABAergic interneurons, in particular parvalbumin-positive, fast-spiking interneurons, such as basket cells.34,35,50,51 In generating these patterns of activity, pyramidal cells excite fast-spiking inhibitory interneurons that in turn transiently inhibit the perisomatic region of pyramidal cells through rhythmic GABA release. During gamma oscillations, for example, individual pyramidal cells generate action potentials at 1–3 Hz, whereas fast-spiking inhibitory interneurons show much higher firing rates (>20 Hz).22,34 We note that fast-spiking inhibitory interneurons were not targeted with our Mcu-KO strategy, but we nevertheless see a strong phenotype.
We demonstrate disturbances in the generation of gamma oscillations and the underlying neuronal spiking. In addition, we report the lack of activity-dependent PDH dephosphorylation in Mcu-KO mice. This indicates that the Ca2+-dependent stimulation of the mitochondrial TCA cycle is impaired, in line with other reports30,56 but see Nichols et al. 55 Therefore, our data might primarily reflect an energy deficit in excitatory pyramidal during gamma oscillations that feature high energy expenditure.22,37 Similar results have been reported for the skeletal muscle during strenuous work in a different Mcu–/– mouse mutant. 30 Since the overall spiking rates were similar in our wild type and Mcu-KO mice, the energy deficit might arise in specific subcellular compartments with high energy expenditure, such as glutamatergic presynaptic terminals.57,58 This might, for example, result in less precise excitation of fast-spiking inhibitory interneurons through alterations of the synaptic vesicle cycle.10,22,59
However, we also demonstrate disturbances in the generation of sharp wave-ripples that are thought to involve less energy expenditure compared with gamma oscillations. 37 Therefore, the functions of the MCU in excitatory pyramidal cells may not be restricted to neurometabolic coupling and energy metabolism (Figure 7(e)). Similar to other excitable cells, the MCU also has an important role in Ca2+ homeostasis in neurons.2,60,61 During neuronal activity, mitochondrial Ca2+ uptake significantly shapes cytoplasmic Ca2+ transients and thereby modulates neural excitability9,62,63 and the release of neurotransmitters,4,7,59,64,65 including nitric oxide.45,66,67 Thus, loss or dysfunction of the MCU might affect neuronal signalling on different levels.
Disturbances of gamma oscillations and sharp wave-ripples might affect spike-timing and synaptic plasticity in local cortical networks as well as information transfer between remote cortical networks.38–40
Our data suggest that dysfunction of the MCU might result in cognitive impairment, with putative relevance to several neurological and psychiatric disorders.2,13,22,68,69
Absence of adaptations in gene expression
Similar to previous studies in different MCU loss-of-function models,30,55,56 we found no evidence for impaired cellular function under basal conditions in our newly generated Mcu-KO mouse line. As suggested previously, 2 these moderate phenotypes might be due to compensatory mechanisms. However, we found no adaptations in gene expression related to mitochondrial functions and glucose metabolism. Therefore, our data support the notion that stimulation of mitochondrial metabolism by matrix Ca2+ is dispensable for basal neuronal function. 2 In fact, recent work has shown that modest stimulation of neurons leading to moderate workloads and moderate increases in cytosolic Ca2+ concentration can stimulate mitochondrial ATP production via Ca2+-regulated mitochondrial substrate carriers, such as the Aralar/AGC1 malate-aspartate-shuttle system and SCaMCs (small calcium-binding mitochondrial carriers). 54 Moreover, mitochondrial oxidative phosphorylation is further regulated by a variety of Ca2+-independent mechanisms, such as the ratios of ADP/ATP, NAD/NADH or CoA/acetyl CoA.70,71 Consistent with its high Ca2+ concentration threshold for Ca2+-uptake,72,73 recruitment of the MCU complex for neurometabolic coupling might therefore be especially relevant during neuronal network activity states that feature large increases in intracellular Ca2+ and high energy expenditure, such as gamma oscillations.
Conclusions
We provide substantial evidence that the neuronal MCU is crucial for the generation of fast cortical network rhythms, most likely by adapting oxidative phosphorylation, and perhaps by controlling cytoplasmic Ca2+ homeostasis. Our study might contribute an increased understanding of the functions of the MCU during neuronal information processing. Future studies on neuronal Ca2+ homeostasis and synaptic transmission as well as on the behavioural level will further clarify these relationships.
Supplemental Material
JCB887777 Supplementary material - Supplemental material for The mitochondrial calcium uniporter is crucial for the generation of fast cortical network rhythms
Supplemental material, JCB887777 Supplementary material for The mitochondrial calcium uniporter is crucial for the generation of fast cortical network rhythms by Carlos Bas-Orth, Justus Schneider, Andrea Lewen, Jamie McQueen, Kerstin Hasenpusch-Theil, Thomas Theil, Giles E Hardingham, Hilmar Bading and Oliver Kann in Journal of Cerebral Blood Flow & Metabolism
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Deutsche Forschungsgemeinschaft (Collaborative Research Center 1134, projects B02 and C03; FOR2289, BA1007/9-1 and BA3679/4-2) and also by the UK Medical Research Council, Alzheimer’s Research UK and Alzheimer’s Society.
Acknowledgements
The authors thank Yan-Wei Tan for providing shRNA against LacZ.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors’ contributions
Carlos Bas-Orth: Conceptualization, Funding acquisition, Investigation, Formal analysis, Visualization, Writing – original draft, Writing – review and editing
Justus Schneider: Investigation, Formal analysis, Visualization, Writing – review and editing.
Andrea Lewen: Investigation, Formal analysis, Visualization, Writing – review and editing.
Jamie McQueen: Resources, Investigation, Formal analysis, Writing – review and editing.
Kerstin Hasenpusch-Theil: Resources, Interpretation, Writing – review and editing.
Thomas Theil: Resources, Interpretation, Writing – review and editing.
Giles E Hardingham: Conceptualization, Resources, Project administration, Writing – review and editing.
Hilmar Bading: Conceptualization, Funding acquisition, Project administration, Writing – review and editing.
Oliver Kann: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review and editing.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.