
Abstract
Stroke is a leading cause of long-term disability, with around three-quarters of stroke survivors experiencing motor problems. Intensive physiotherapy is currently the most effective treatment for post-stroke motor deficits, but much recent research has been targeted at increasing the effects of the intervention by pairing it with a wide variety of adjunct therapies, all of which aim to increase cortical plasticity, and thereby hope to maximize functional outcome. Here, we review the literature describing neurochemical changes underlying plasticity induction following stroke. We discuss methods of assessing neurochemicals in humans, and how these measurements change post-stroke. Motor learning in healthy individuals has been suggested as a model for stroke plasticity, and we discuss the support for this model, and what evidence it provides for neurochemical changes. One converging hypothesis from animal, healthy and stroke studies is the importance of the regulation of the inhibitory neurotransmitter GABA for the induction of cortical plasticity. We discuss the evidence supporting this hypothesis, before finally summarizing the literature surrounding the use of adjunct therapies such as non-invasive brain stimulation and SSRIs in post-stroke motor recovery, both of which have been show to influence the GABAergic system.
Introduction
Chronic stroke is one of the commonest causes of long-term neurological disability worldwide. There are approximately 17 million first-time strokes worldwide each year, 1 leaving over 5 million people living with moderate to severe disabilities as a result of stroke. 2 Over recent years, there have been significant improvements in treatments administered during the acute stroke phase, but little is currently available to aid long-term recovery. Recovery of motor skills is of high importance for stroke survivors,3,4 but intensive physiotherapy, the current “gold-standard” intervention, is expensive, in short supply, and inherently limited by the activity of the residual cortex. In current practice, half of all people who survive a stroke are left permanently disabled, and a third are completely reliant on others for normal activities of daily living, significantly impacting on their independence. 5 Given the increasing number of stroke survivors, this long-term morbidity presents a significant, urgent, unmet clinical burden, meaning there is an increasing need to develop plasticity-inducing adjunct interventions to extend the potential benefits of therapy in a cost-effective manner.
The rationale for adjunctive therapies stems from the hypothesis that the beneficial effects of treatments, such as physiotherapy, are mediated via a combination of structural and functional plastic brain changes. Interventions that modulate these physiological processes, such as drugs or brain stimulation, might therefore be used to enhance the effects of standard regimes. However, before we can develop successful adjunct therapies, it is vital to understand the neurochemical changes underpinning functional recovery after stroke, and how these potential adjunct therapies may interact with this. Remarkably little is currently understood about the physiological mechanisms underlying naturally occurring plasticity, making optimizing interventions to promote maximal functional recovery difficult. For example, multiple brain regions are engaged when a stroke survivor moves their affected hand, but there is highly conflicting evidence as to whether recruitment of these regions is an adaptive or maladaptive process.6–8
Here we will provide an overview of what is known about the natural history of stroke recovery. We will then go on to discuss the methods for quantifying neurochemicals in humans, what is known about the neurochemical changes following stroke, and how these changes relate to stroke recovery. The majority of this work has examined motor function – in particular upper-limb function – and therefore we will concentrate on this aspect of recovery after stroke. As a comparative model for the phases of stroke recovery, motor learning in healthy individuals will be discussed. We will review the interventions known to modulate neurochemistry, such as brain stimulation and drugs, have been used to enhance both healthy learning and stroke recovery.
The natural history of stroke recovery
Although the presenting symptoms and signs of a stroke differ across patients depending on the site of the injury, stroke is caused by a disruption in normal blood supply to the brain; 85% of strokes are ischaemic, where perfusion is disrupted due to a blockage of a blood vessel from a variety of different causes, and 15% of strokes are haemorrhagic, caused by rupture of a blood vessel within or on the surface of the brain. 5 Haemorrhagic strokes have classically been associated with a significantly higher mortality and morbidity than ischaemic strokes, within the first three months 9 and beyond, 10 though this has not been found in all cohorts. 11
The details of the aetiology and pathophysiology of ischaemic and haemorrhagic strokes are outside the scope of this review (see, for example O'Donnell et al. 12 ). However, stroke type is an important factor to consider in early intervention, particularly, as it is likely that ischaemic and haemorrhagic strokes have a different natural history of recovery13,14 and therefore stroke survivors may respond differently to different interventions. However, very few interventional trials of the types discussed here include significant numbers of stroke survivors with haemorrhagic strokes and therefore it is difficult to fully assess the impact of different stroke aetiologies on recovery.
Spontaneous functional recovery is fastest in the first days to weeks after a stroke, a time known as the acute and sub-acute periods. However, it is essential to note that rehabilitation induced improvements can still be made months to years after a stroke during what is often termed, the chronic phase, which is normally taken to start at six months post event. It is likely that the observable functional improvements in patients are underpinned by at least two partially or entirely dissociable mechanisms, (see section on “Neurochemical changes occurring after stroke” below).
Methods for quantifying neurochemicals in humans
A number of techniques have been used to understand neurochemical activity in humans, including magnetic resonance spectroscopy (MRS), transcranial magnetic stimulation (TMS) and positron emission tomography (PET). Like all techniques, these have relative advantages and limitations, and are differentially sensitive to aspects of neurochemical activity within the brain. We will first review these techniques here, giving an overview of the methodology and most important limitations of the measures derived from them, before going on to discuss what the evidence from all these techniques can tell us about the role of changed neurochemical signaling after stroke in later sections.
MRS
MRS is a non-invasive technique that allows accurate quantification of neurochemicals in vivo. Like all MR approaches, MRS is only sensitive to molecules containing particular nuclei, such as 1H, 13C and 31P. When these nuclei are placed in a strong magnetic field, the net preferred direction of these atoms aligns toward, and precesses around the main magnetic field (B0). In conceptual terms, to produce a measurable signal within the MR scanner, atoms are excited away from the B0 field by applying a short radio frequency (RF) pulse at the known frequency of the precessing nuclear spins. When this RF magnetism is removed, the atoms’ spins begin to rotate around the axis of the B0 field as they return to their preferred position. This rotating magnetism then causes an oscillating current detected by the ‘receive coils’ of the MR scanner, which can be recorded and analysed.
The behaviour of these nuclei within the magnetic field is influenced by interactions both with other nuclei of the molecule within which they are bound, and the environment surrounding these molecules. Shielding of the atoms by electrons from neighbouring atoms affects the magnitude of the magnetic field experienced by the nucleus. This alters the resonant frequency of the signal arising from the nucleus. This difference in signal frequency between otherwise identical nuclei is therefore dependent on the molecular structure in which the nucleus is found, and is termed chemical shift. If the signal arising from a given region of tissue is plotted as a function of frequency, then a series of peaks in the signal can be seen, each corresponding to the nucleus of interest bound in different molecules. The characteristic peaks and frequencies of many neurochemicals are known and can be identified from within the spectrum. The concentration of a given neurochemical within the selected volume is proportional to the amplitude of its representative peak(s) within the spectrum, although accurate quantification often requires complex analysis approaches, with many important confounds to account for. A full description of MRS acquisition and analysis approaches is beyond the scope of this review, but the interested reader is directed to the many reviews of the topic, including Stagg and Rothman. 15
1H-MRS can quantify neurochemicals including the inhibitory neurotransmitter γ-aminobutyric acid (GABA), and the excitatory neurotransmitter glutamate, providing a method for assessing physiological processes across the brain. However, MRS has some technical limitations, which govern the interpretation of findings. MRS has an inherently low signal-to-noise (SNR), requiring the use of relatively large voxel sizes (in the order of a few centimeters) and acquisition times of a few minutes, limiting both its spatial and temporal resolution. In addition, MRS relies on structural differences between neurochemicals to separate their resonances and it can be difficult to accurately separate closely related molecules such as glutamate and glutamine, which are often presented as the composite measures Glx. (see Figure 1 for a representative spectrum, acquired using the MEGA-PRESS sequence at 3T).
Schematic diagram illustrating: (a) a typical in vivo single voxel locaslied MEGA-PRESS spectrum from a control subject at 3 T, highlighting the MEGA-PRESS technique. The primary GABA resonance is found underlying the Creatine resonance at 3 ppm. An edited approach is therefore taken, where an editing pulse at 1.9 ppm modulates the shape of the GABA resonance at 3 ppm. (b) Scans taken without these pulses (OFF) are subtracted from those taken with the editing pulses at 1.9 ppm (ON), revealing a GABA signal in the difference spectrum (DIFF), as Creatine is unaffected. Figure adapted from Mullins et al.,
181
with permission.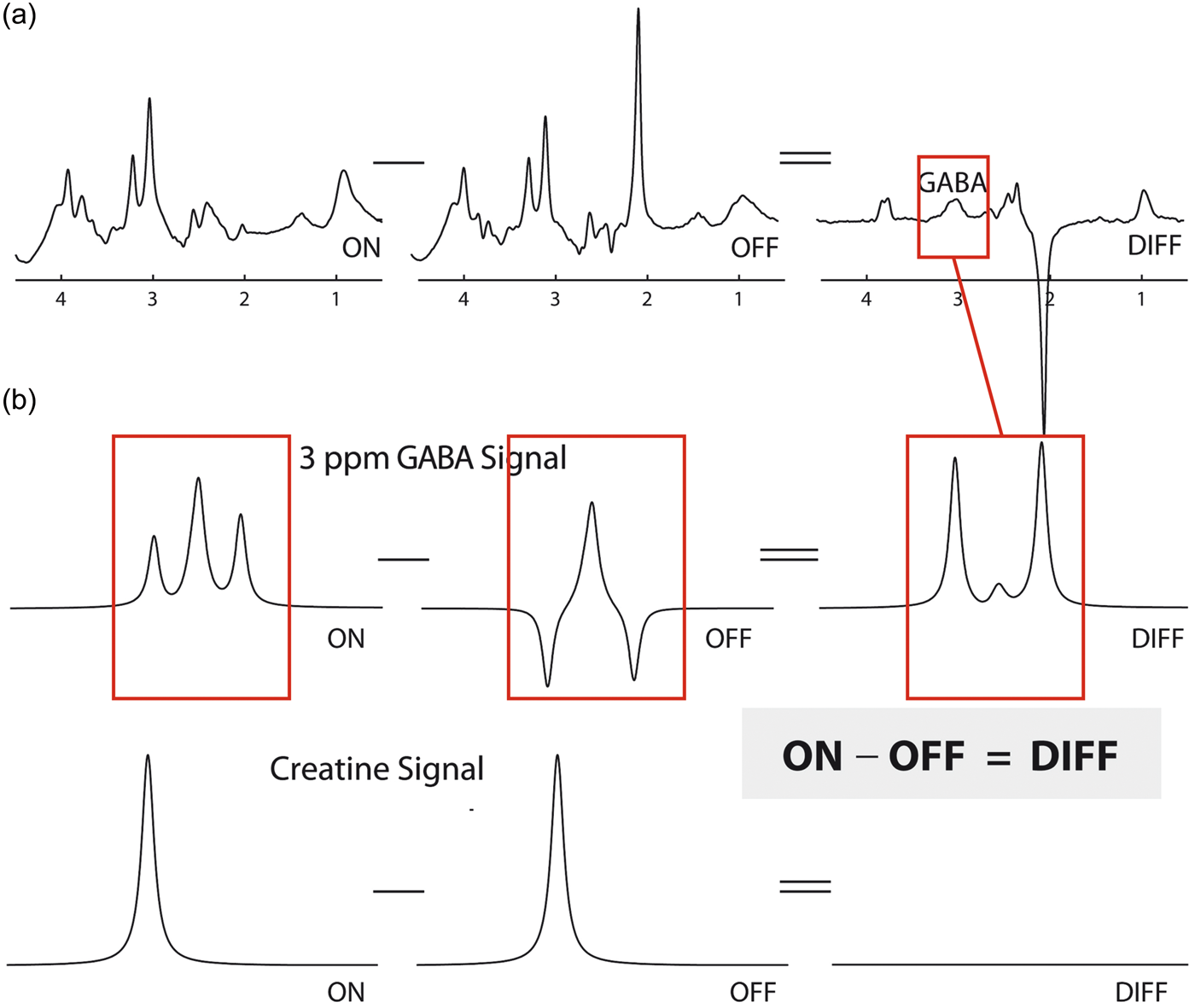
Perhaps most importantly when interpreting MRS-derived results, it is important to highlight that MRS-derived measures of neurotransmitters give a measure of the total concentration of that neurochemical within the voxel of interest. It is therefore not possible to discriminate between different metabolic pools. This is of particular importance for GABA and glutamate, which both have substantial roles in brain metabolism as well as neurotransmission. In addition, MRS is not directly sensitive to changes in synaptic density or function, for example in NMDA receptors underlying plasticity in the motor cortex, meaning that at least some important changes in glutamatergic signalling will not be directly visible to MRS approaches.
Finally, and of particular relevance here, MRS is sensitive to the total amount of a neurotransmitter within a given volume of interest, and is always given in reference to another spectral peak, most commonly water, creatine or N-acetylaspartate (NAA). However, this poses some inherent problems in situations where it is likely that there are some changes in tissue structure, either macroscopic in terms of stroke lesion, or microscopic in terms of diffuse cell loss. Care has to be taken to exclude core lesion-affected anatomy from the volume of interest and to choose a reference peak appropriately. For a full discussion of all these issues, please see Stagg. 16
TMS
TMS can be used as a method to measure the activity of certain neurotransmitter systems within the motor cortex. In TMS, magnetic pulses are generated and delivered to a brain area of interest through a hand-held coil, positioned on the scalp. These pulses then induce an electrical current in the underlying brain area via electromagnetic induction. This electrical current causes depolarization of cortical neurons leading to firing of action potentials. If the coil is held over the primary motor cortex, M1, this neuronal discharge can cause motor-evoked potentials (MEPs), triggered within muscles, which can then be recorded using electromyography (EMG) (Figure 2(a)).
(a) Image showing subject receiving transcranial magnetic stimulation (TMS) to left primary motor cortex in order to induce motor evoked potentials (MEPs) within the right hand. These MEPs within the muscles are recorded using electromyography (EMG), which is also shown. Image reproduced with permission from John Cairns. (b) Showing input-output curves for 1 ms short interval cortical inhibition (SICI), 2.5 ms SICI and intracortical facilitation (ICF). These were acquired with conditioning stimulus (CS) intensities of 60%, 70%, 80% and 90% of aMT and a test stimulus (TS) of the necessary intensity to evoke an MEP of approximately 1 mV peak-to-peak amplitude. MEP amplitude is given as a percentage ratio of the condition divided by the unconditioned stimulus (each point represents the group mean ± SEM). Figure reproduced with permission from. Stagg et al.
45
(c) Schematic of four different repetitive transcranial magnetic stimulation (rTMS) pulse protocols. 1 Hz rTMS and continuous theta burst stimulation (cTBS) protocols have inhibitory effects on cortical excitability, indicated by red arrows; whereas 3 Hz rTMS and intermittent TBS (iTBS) increase cortical excitability. Figure reproduced with permission from Stagg and Johansen-Berg.
182
The number of pulses per second is indicated, and the characteristic total time over which the TMS is applied is stated.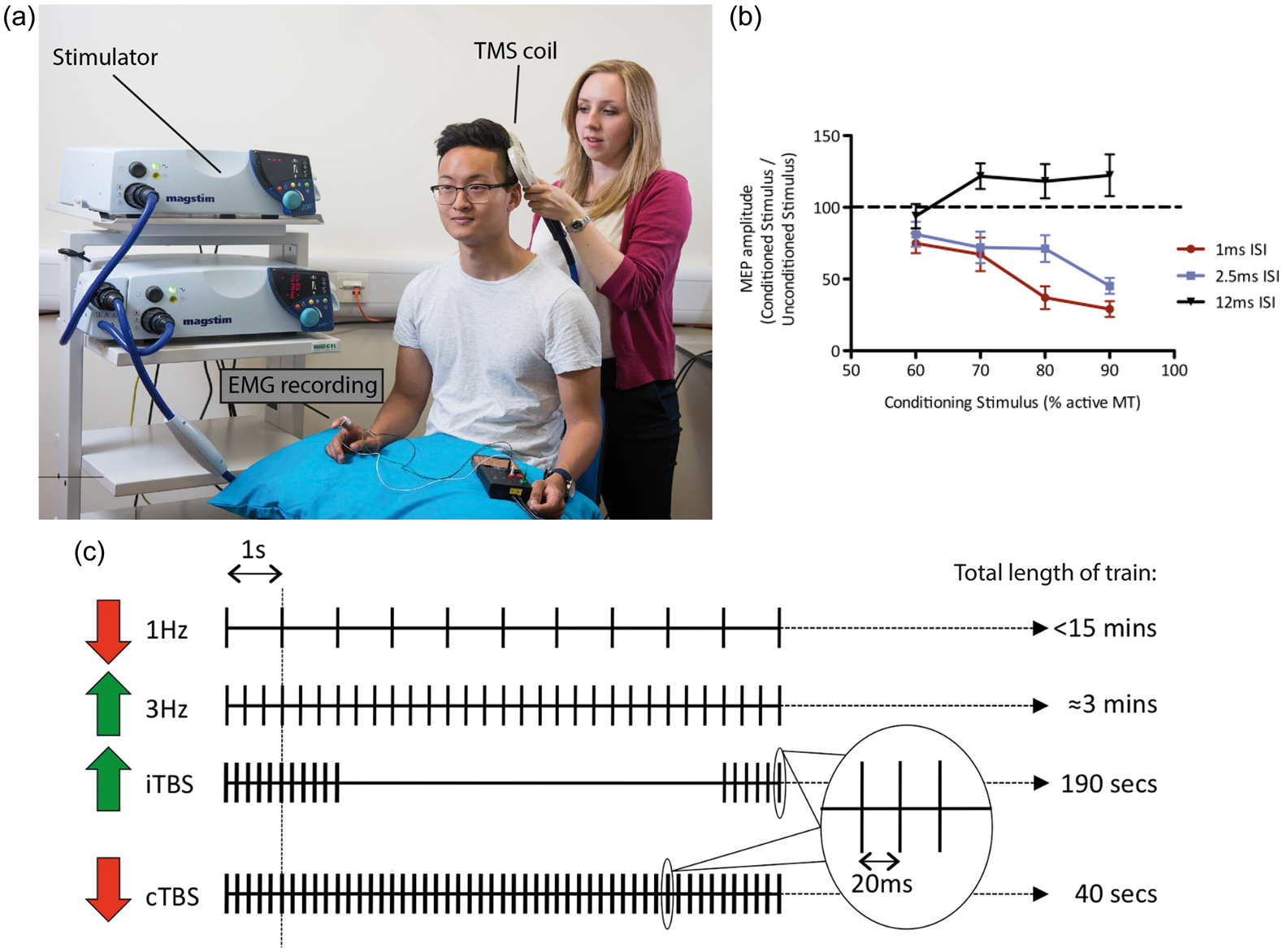
The stimulation intensity required to elicit MEPs from within certain muscles varies highly between individuals. Because of this, most studies will begin by determining the stimulation intensity, at a given coil position, which produces MEP of a predetermined size within the target muscle. This intensity can then be used in subsequent measures, and MEP amplitude can be recorded and examined relative to baseline.
In addition to stroke-affected TMS measures, MEP amplitude in response to single pulses is also influenced by several drugs affecting a variety of neurotransmitter systems, and the exact physiological underpinnings are complex and non-specific. For example, GABAA receptor agonists such as benzodiazapines28–30 and barbituates 31 have been shown to depress high amplitude but not low amplitude MEPs. Similar results have also been shown for the dopamine- (DA) receptor agonist cabergoline 32 and the noradrenaline- (NA) antagonist, guanfacine. 33 Furthermore, drugs such as NA-agonists,34–36 selective serotonin reuptake inhibitors (SSRIs), 37 and muscarinic receptor M1 agonist, scopolamine, 38 have all been seen to increase MEP amplitude.
Paired-pulse TMS
One approach allowing for additional neurochemical specificity is to administer paired-pulse TMS [ppTMS] protocols. In contrast to single-pulse TMS [spTMS], ppTMS can be tailored to assess the activity of specific neurochemical systems. ppTMS involves the application of two pulses to the same scalp location, the first a sub-threshold conditioning pulse, and the second a supra-threshold test pulse. As in spTMS, the supra-threshold test pulse causes an MEP via induction of an action-potential in the pyramidal neurons, whereas the sub-threshold conditioning stimulus stimulates only the cortical interneurons, which have a lower threshold for stimulation. The influence of the interneuronal pool on the output of pyramidal neurons depends on signaling across synapses, the time-course of which varies between different post-synaptic receptors, different synapses, meaning that the influence of interneuron activity on the changes with time. Therefore, by varying the inter-stimulus interval (ISI) between the two pulses, the activity within different neurotransmitter systems can be assessed. The difference in size between the resulting paired-pulse MEP, and an MEP elicited to a single test pulse at the same intensity, is thought to be proportional to activity within the targeted neurotransmitter system (Figure 2(b)).
Short-interval intra-cortical inhibition (SICI) can be achieved using an ISI of 1–5 ms, and is thought to be mediated by ionotropic GABAA receptor activation. Several studies have shown that drugs acting as GABAA receptor agonists, for example the benzodiazapines, enhance SICI with an ISI of 2.5 ms via an increase of GABAA signalling.29,39–41 Long-interval intra-cortical inhibition (LICI) can be achieved using an ISI of 50–200 ms between the conditioning and test stimulus. 42 LICI is thought to reflect metabotropic GABAB receptor-mediated inhibition, with baclofen, a specific GABAB receptor agonist, causing an increase in LICI magnitude. 43 In further support of the distinction between SICI and LICI, use of the GABA reuptake inhibitor tiagabine has differential effect on the two intracortical inhibition types, causing an decrease in SICI magnitude and an increase in LICI magnitude. 44
Despite clear results from drug studies, relating paired-pulse TMS measures of GABA receptor activity to MRS-measured GABA has proved complex. 45 Neither 2.5 ms SICI, 150 ms LICI nor a linear combination of the two can be significantly related to MRS-measured GABA levels; however, a significant relationship has been demonstrated with 1 ms SICI. It has previously been proposed that 1 ms and 2.5 ms SICI reflect different cortical mechanisms,46,47 with the 1 ms SICI perhaps reflecting extra-synaptic GABAergic tone which may be more visible to MRS measurement. 45
As well as assessing GABA receptor activity, ppTMS with an ISI of between 10 and 15 ms [intra-cortical facilitation (ICF)] primarily reflects glutamaterigic receptor activity. One particular glutamatergic receptor, the NMDA receptor, is critically important in long-term potentiation (LTP),48,49 a highly complex cascade by which synapses are strengthened that has been proposed as the physiological mechanism underpinning learning. ICF is decreased by NMDA receptor antagonists.50,51 However, this relationship appears to be more complex than that of SICI and LICI, with one study finding no effect of NMDA receptor agonists, 52 and others finding an increase in ICF with benzodiazapines, which act as positive modulators of the GABAA receptor.30,31,41 These later studies indicate that there is a contribution of GABAA receptor-mediated inhibition to the magnitude of ICF, and therefore ICF cannot be taken to be a pure measure of glutamatergic activity.
TMS approaches are valuable as they are relatively neurochemically and anatomically specific, and are generally well-tolerated. In the context of stroke survivors, standard TMS approaches targeting the motor system rely on the integrity of the corticospinal tracts, as the output measure is an EMG-derived MEP. In the stroke population, this can be difficult to acquire in more impaired patients, meaning that TMS studies in this population are often inherently biased towards patients who are better recovered, who are more likely to have at least relatively intact corticospinal tracts. It is possible to use other metrics as a readout, for example EEG, but although undoubtedly valuable, this is technically challenging and complex to interpret, limiting its use in the stroke population. 53
PET
PET is an imaging method involving the use of radioactive tracers to detect targeted biological processes. The nuclear decay of a radioactive tracer, most commonly, 11C, 15O, 13N and 18F, provides the basis of the signal measured in PET imaging. This tracer is bound to a biologically active molecule, which will, for example, bind to a specific receptor, and therefore allows for the targeting of specific physiological processes in the brain.
Importantly, in PET imaging, radiotracers do not disrupt normal biological function and the radioactive half-lives are relatively short-lived. While the temporal resolution, occurring on a timescale of minutes, and the spatial resolution achieved by PET are not as precise as in other imaging methods (e.g. fMRI, EEG or MEG), the unique advantage of PET imaging is the ability to utilize highly specific radiotracers to observe interactions with selected proteins, receptor binding potential, catecholamine or amino acid transport, protein synthesis, among many other neurochemical concentrations or processes.
In the context of stroke, early PET imaging studies obtaining measures of perfusion and oxygen consumption were able to differentiate core stroke lesion and infarct tissue from that of the penumbra in humans, 54 relate the viability of that penumbral tissue to recovery of function, 55 and examine cerebral blood flow associated with moving the contralesional hand. 56
Neurochemical changes occurring after stroke
MRS, TMS and PET have all been used to study the neurochemical changes occurring post-stroke. Across all of these modalities, measurements of GABA have been shown to be altered post-stoke, both while individuals are at rest and during movement. This literature will be discussed, before highlighting the existing evidence as to how these neurochemical changes relate to functional recovery.
Observed changes in GABA
Changes in GABA at rest
A number of studies have investigated GABA changes after stroke while the subjects are at rest. For example, recent PET approaches to studying stroke and motor recovery have made use of [18F]flumazenil ([18F]FMZ 57 ) and [11C]flumazenil (([11C]FMZ58–60) to infer GABAA concentrations, localize GABAergic activity and as a proxy measure for quantifying neuronal integrity. 2-deoxy-2-18F-D-glucose (FDG 61 ) has also been used in stoke to observe regional metabolic rate of glucose (rCMRglc). In a recent PET study, Kim et al. 57 assessed changes in cerebral GABAA activity using [18F] FMZ in 10 unilateral subcortical ischemic stroke survivors at one and three months post stroke. 57 One month post stroke, patients had significantly increased GABAA-ergic activity compared to 15 age-matched controls, although this had largely normalized by three months post-stroke.
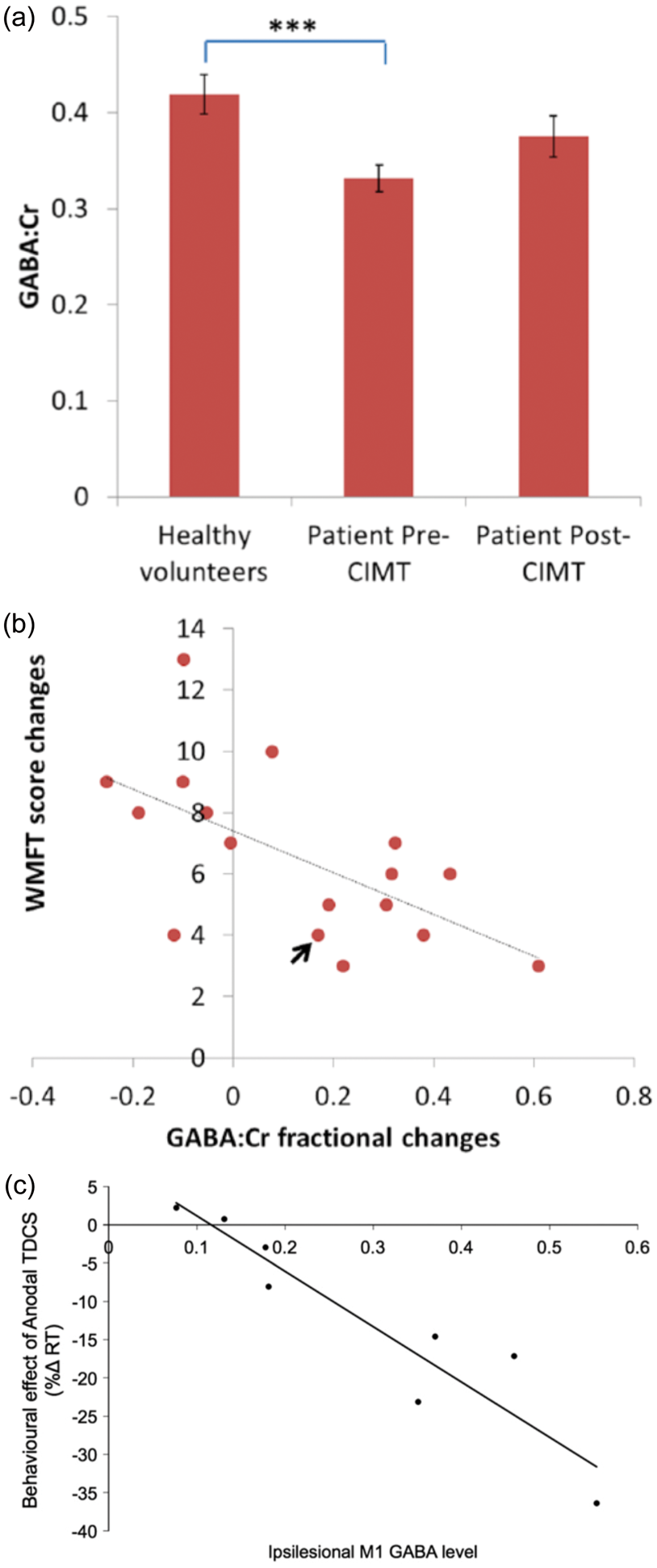
Using MRS, a recent study has shown that GABA levels are significantly decreased in chronic stroke patients compared with age-matched healthy controls
84
(see Figure 3(a)). One study demonstrated decrease in GABA within the primary motor cortex in individuals in the chronic phase of stroke,
85
while another showed this decrease in non-lesioned prefrontal lesions in patients both early after stroke and at three months.
84
(a) Magnetic resonance spectroscopy (MRS) measured GABA:Cr levels in stroke patients either before (n = 19) or after (n = 17) constraint induced movement therapy (CIMT), as well as in 20 age and gender-matched healthy volunteers. Healthy vs. pre-intervention comparison made using independent t test (t(37) = −3.88, P < .001) (identified with *** on figure) and pre vs. post-intervention comparison using paired t test on the 17 patients with both pre and post MRS data. MRS was carried out at 3 T, using a voxel (2 × 2 × 2 cm) placed in ‘hand knob’ region
183
of affected hemisphere for patients and the dominant hemisphere of healthy controls. Error bars represent standard error of the mean. Figure adapted with permission from.
83
(b) Negative correlation between change in score (post training score minus pre score) on the Wolf Motor Function Test (WMFT) and GABA:Cr (continued) concentration fractional change (R = −0.61, P = .009). The WMFT is a quantitative measure of upper extremity motor ability, judged using a mixture of timed and functional tasks. A higher score indicates better function (with a maximum possible score of 75). The single arrow indicates a patient with a lesion in the hand area. (c) Significant relationship (Pearson’s r(8) = −0.936, P = 0.001) between MRS measured GABA:NAA levels in ipsilesional M1 in stroke patients correlated with the behavioral effect of anodal tDCS measured as a change in reaction time (RT) in a simple reaction time task, such that higher GABA:NAA predicted larger RT gain in response to anodal tDCS compared to sham tDCS. Figure reproduced with permission from O'Shea et al.
153
Similar decreases in resting GABA have been demonstrated using ppTMS approaches.85–88 Intracortical inhibition, as measured by SICI at varying delays, appears to be reduced in the ipsilesional M1, as compared to the contralesional M1, or the same M1 in controls. 87 As with the MRS measurements, this decrease in inhibition relative to a control group is also consistently demonstrated across both acute and chronic time points. 88 As discussed above, TMS is technically more difficult in patients than controls, as corticospinal tract damage creates difficulty in acquiring MEPs. Therefore, TMS studies are inherently biased towards the relatively well-recovered patients in whom an MEP can be measured. However, despite the difficulties surrounding TMS measurements, the converging evidence from all the discussed modalities points towards a global reduction in inhibition within the cortex, at least in the chronic stages of post-stroke recovery.
Functional changes in GABA
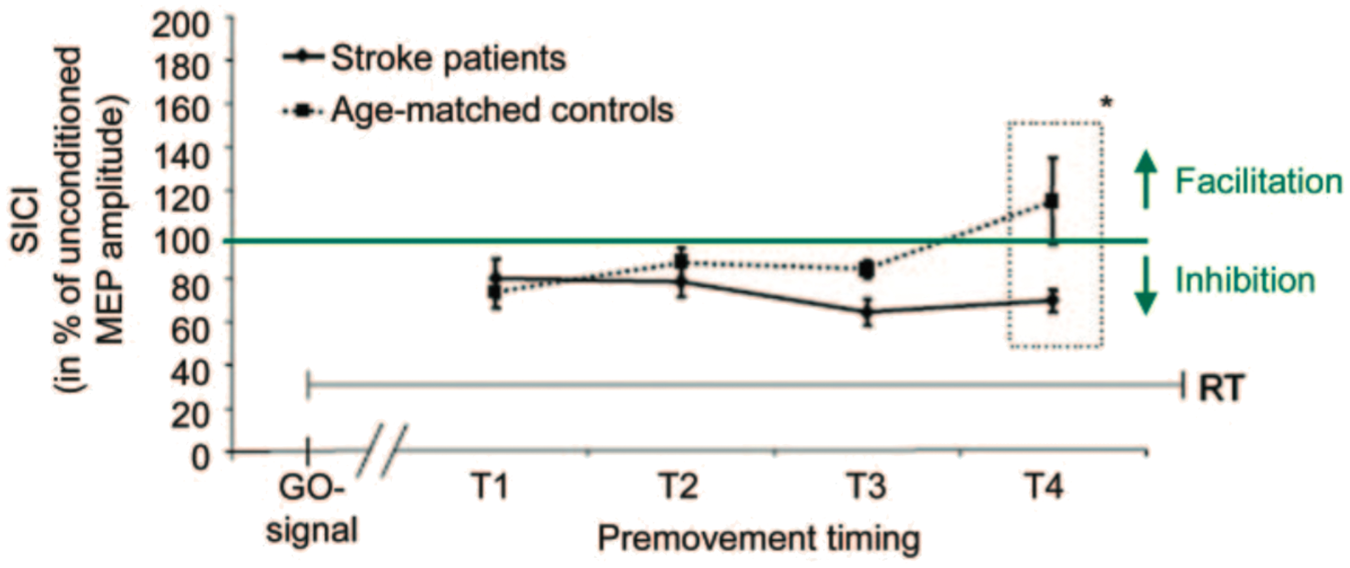
As well as measuring GABA activity within M1 at rest, ppTMS approaches can be used to measure functional changes occurring in local circuits as a subject prepares to move.90–93 During healthy movement preparation, a decrease in GABAA activity has been observed, such that immediately prior to the time of movement, the inhibitory effect of SICI is significantly reduced, or seen even as a facilitation.90,92,94 In stroke patients, this normal decrease in SICI is significantly reduced compared with age-matched healthy controls, i.e. patients are unable to release local inhibition when making a movement
90
(see Figure 4). Taken together with the findings above, it is possible to formulate a hypothesis that while resting levels of inhibition are reduced in chronic stroke patients, possibly as a result of on-going long-term plasticity, local inhibition is less flexible. This would mean that patients are unable to release inhibition sufficiently to enable normal movement. This theory reconciles the somewhat conflicting evidence, discussed above, about changes in inhibitory processing post-stroke, though further work needs to be done to establish whether the functional changes in inhibition seen in patients is pathological, or a compensatory change that promotes function via better coherence of muscle contraction.
Short interval cortical inhibition (SICI) decreases significantly in healthy age-matched controls (n = 14) prior to movement onset, but no such decrease is visible in stroke patients (n = 14). SICI was measured at four timings between cue delivery and movement onset (displayed on X-axis). Y-axis is the SICI MEP measurement as a percentage of the unconditioned MEP (values less than 100% represent inhibition). *2-sample t-test: t(26) = 2.23, p < 0.05. Figure reproduced with permission from Hummel et al.
90
How do changes in GABA relate to post-stroke recovery?
The mechanisms underpinning recovery after stroke are not fully understood, and much work is aimed at elucidating the neurochemical changes, with a view to develop effective adjunct therapies. It has been hypothesized that recovery of function after a stroke can be caused by two separate but complementary mechanisms, that of true recovery, where movement patterns close to those that were used pre-stroke are regained, and compensation, where the patient uses alternative movements or combinations of movements to achieve the same end-goal. 64 It seems likely that a combination of recovery and compensation supports functional improvements in the majority of stroke patients, with spontaneous brain recovery, occurring during the post-ischaemic sensitive period, being underpinned by a neurochemical milieu quite distinct from the neurochemical changes underpinning learning and compensation later in recovery.
Post- ischaemic sensitive period: the role of GABA
Rodent stroke models have suggested that very early after stroke, a post-ischaemic ‘sensitive period’, where the neurochemical milieu strongly encourages neuroplasticity, promotes rapid spontaneous recovery even in the absence of training. In rodent models of stroke, changes in molecular, cellular and physiology occur in the days to weeks following stroke. 95 This distinct neuroplastic milieu is characterised by enhanced synaptic plasticity, increased cortical excitability, and a complex modulation of both phasic and tonic inhibition.64,96 In a recent study, Zeiler et al. 97 demonstrated in a mouse model the ability to re-open the post-ischemic sensitivity period by inducing a second lesion in a functionally related region to the first (i.e. caudal forelimb area and medial premotor area). The deficits observed from the first lesion counter-intuitive were ameliorated following the second lesion, ultimately resulting in motor recovery to pre-stroke proficiency.
Directly reducing pathologically increased GABA-mediated tonic inhibition in the peri-infarct tissue using either pharmacological or genetic manipulations significantly improved recovery early after stroke in a mouse model.
63
In addition, also in a murine model, post-stroke functional recovery could be increased by augmenting phasic GABA early after stroke with zolpidem, a positive allosteric GABA modulator
96
(Figure 5(a)). Increasing serotonin has also been demonstrated to significantly extend the length of the sensitive period in a mouse stroke model via modulation of GABAergic activity.
65
(a) In mice, the delivery of Zolpidem (an FDA-approved positive allosteric GABA modulator; n = 16) three days post occlusion-induced stroke to the somatosensory cortex, facilitated improved functional recovery compared to stroke affected mice treated with a vehicle solution (n = 15; repeated measures ANOVA; P = 0.0072; *P < 0.05, **P < 0.01 for stroke + vehicle versus stroke + zolpidem). Further, administering zolpidem (compared to vehicle) to non-stroke (i.e. healthy) mice in whom no behavioural improvements were seen resulted in no difference in functional performance, strongly suggesting that zolpidem modulates plasticity rather than performance per se. Figure is adapted with permission from Hiu et al.
96
(b) Addition of a GABA agonist can block neocortical LTP in the freely moving rat. Prior to LTP induction, field recordings were obtained on three occasions to establish baseline input/output curves. LTP was then induced using 300 Hz trains per day for 10 days, with saline (n = 7), diazepam (a GABA agonist, n = 6) or picrotoxin (a GABA antagonist, n = 5). Clear potentiation is seen in the groups receiving saline and picrotoxin, but no LTP-like plasticity is observed in the presence of diazepam (repeated measures ANOVA, Group by Session interaction F(48,288) = 11.88, p = 0.001).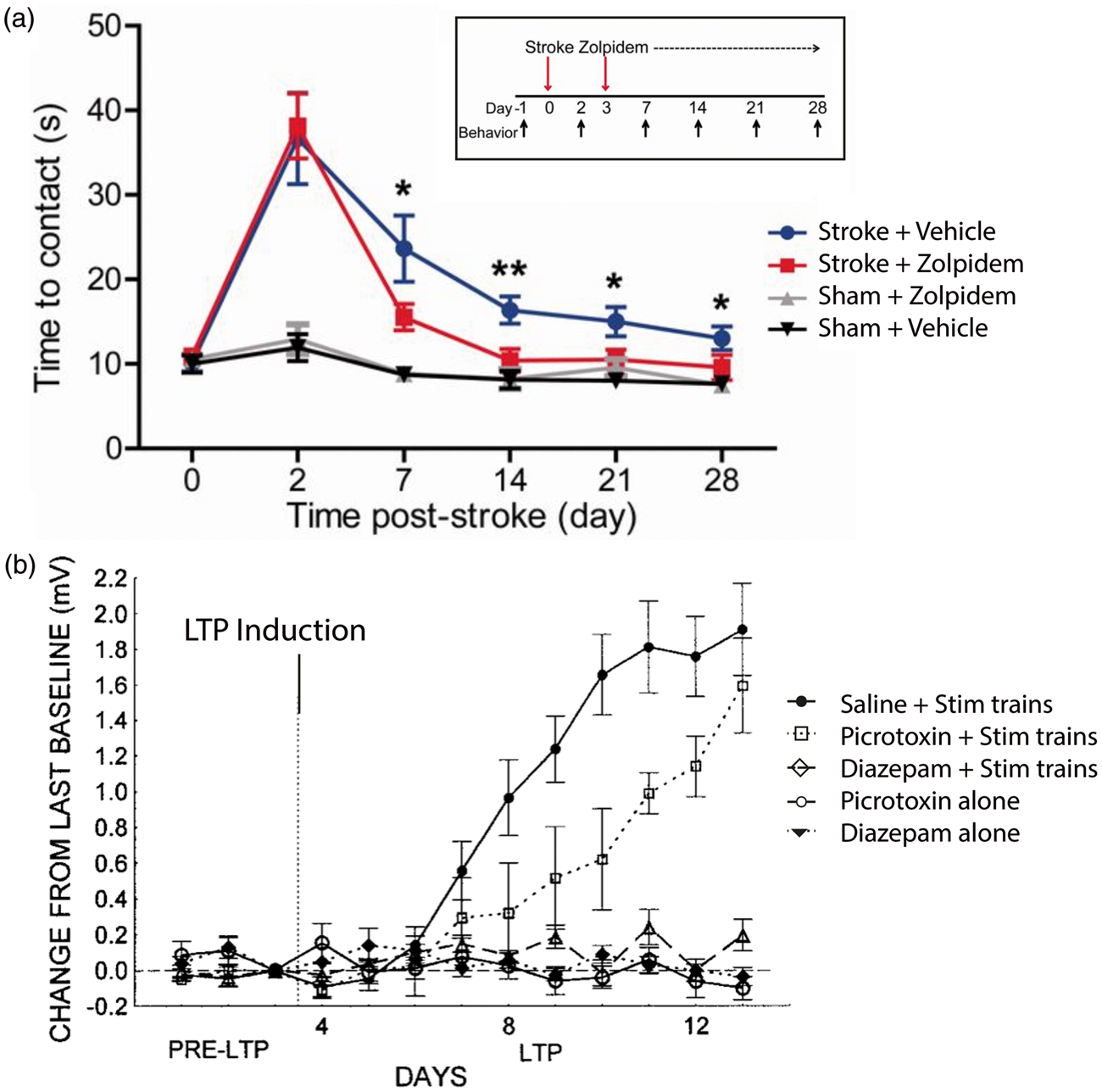
During early stroke recovery in humans, spontaneous recovery of function (including motor, language and other primary neurological impairments) is observed. It is therefore likely that this period is akin to the post-ischaemic sensitive period observed in rodent models; however, it is not clear how long it lasts. Some evidence suggests that a stage with similar properties to the rodent sensitive period might be present for approximately the first three months after a stroke,66,67 though this has yet to be definitively proven. The hypothesis that a post-ischaemic sensitive period exists in humans gives a theoretical framework to develop new treatments, either aimed at directly lengthening this initial period, or developing methods to mimic its pro-plastic milieu.
The immediate post-stroke period has been less studied in humans than in murine models. However, several TMS studies have focused on the changes in cortical excitability occurring during the acute phase after a stroke, the time most likely to be considered as the post-ischemic sensitive period. TMS-induced MEPs tend to be reduced or even eliminated during this time; however, during recovery, MEP size tends to increase,17–20 and TMS threshold tends to decrease.21–25 The levels of TMS measured cortical excitability early after stroke have also been shown to correlate with functional motor recovery.24,26,27 These findings are therefore broadly in line with those from the animal models, and importantly, suggest that neurochemical changes in the acute stages post-stroke can predict functional outcome.
The role of GABA in chronic stroke recovery
As mentioned previously, the neurochemical changes relating to recovery in the chronic phase of stroke are likely to be distinct from those underlying changes in the acute phase.
A recent study by Blicher et al. investigated how motor training modulated GABA levels in chronic stroke. 85 After baseline screening, patients received two weeks of constraint-induced movement therapy (CIMT). As expected, all patients improved on the wolf motor function test (WMFT), a clinical measure of activity. This improvement was not accompanied by a significant overall decrease in GABA, but there was a significant relationship between functional improvement and change in GABA, such that patients who improved the most on WMFT after CMIT were those in whom GABA decreased most 85 (see Figure 3(b)).
These results broadly support the hypothesis that functional improvements after stroke are mediated at least in part by LTP-like plasticity within M1, underpinned by a long-lasting decrease in local GABAergic signaling. The lack of a decrease in M1 GABA in response to CIMT might, at least in part, be explained by the choice of intervention. Although CIMT is a highly effective intervention in terms of clinical improvement, it is not targeted to drive plasticity specifically within the ipsilesional M1. It is possible that training on an intervention more precisely targeted to M1, such as a grip-force modulation task, would lead to a local decrease in GABA, and this should be directly tested.
The majority work focusing on the neurochemical changes post-stroke concentrate on neurochemical changes within the non-lesioned tissue in the ipsilesional cortex, and this review therefore concentrates on this region. However, one of the striking features of neural recovery after stroke is the recruitment of motor regions anatomically distant to the lesion site. In a PET study carried out by Kim et al., 57 improvements in motor function (as indexed by the Fugl-Meyer Assessment; FMA) were shown to correlate with a reduction in GABAA in the contralateral M1. In addition, these authors also identified several time-dependent relationships between GABAA availability at one and three months post stroke and FMA scores, in line with previous findings of time-varying cortical inhibition following stroke.62,63
Studying neurochemical changes in stroke is complex as the clinical population is heterogeneous, and it is not clear how motor learning and true recovery interact in the human brain. In addition, quantification of neurochemicals is necessarily indirect and complicated by the lesion damage to motor regions, making functional imaging and TMS particularly difficult. 89 It is for these reasons that many studies choose to focus on motor learning within healthy individuals as a model for stroke recovery, and it is this that will be discussed in the following section.
Motor learning as a model for stroke plasticity
The physiological processes underpinning compensatory improvements in the chronic stages of stroke recovery are in all likelihood very similar to at least some aspects of motor learning in healthy controls. This allows us to use the learning of motor skills as a model for some aspects of functional recovery after stroke, though this is a model not without complications (for a full discussion of this topic see Krakauer 76 ).
The neurophysiology of motor learning
Hebb
77
clearly described the fundamentals of synaptic plasticity in his seminal 1949 work thus: When an axon of cell A is near enough to excite a cell B and repeatedly or persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells, such that A’s efficiency, as one of the cells firing B, is increased.
The induction of LTP-like plasticity in M1, as in the rest of the brain, is primarily a glutamatergic process, dependent on both pre-synaptic and post-synaptic modulation of glutamatergic synaptic strength (see Feldman 78 for a review). In a manner similar to hippocampal LTP, induction of LTP-like plasticity within M1 relies on the modulation of glutamatergic NMDA receptors. Longer term changes in synaptic strength are likely underpinned by modulation of other glutamatergic synapses, including via AMPA receptor activity.
However, neocortical LTP-like plasticity cannot by induced by the modulation of glutamatergic synaptic strength alone; changes in other neurotransmitters and neuromodulators are also necessary in permitting glutamatergic plasticity to occur. Animal studies suggest that a decrease in GABA is necessary for the successful induction of LTP-like plasticity within M1: GABA agonists block LTP-like plasticity induction 79 (Figure 5(b)), and a reduction of GABA inhibition facilitates LTP-like activity. 80 In addition, it has been postulated that a reduction in GABAergic inhibition may also underlie the “unmasking” of existing horizontal connections, an essential mechanism that underpins the rapid remodelling of motor representations seen in the early stages of plasticity within M1.81,82
As highlighted both here and in the discussion above, both spontaneous brain recovery and motor learning seem to be dependent on GABA modulation, albeit in somewhat complex ways. It is, however, important to note that our focus on the role of inhibition in these processes may be at least in part because GABA is more easily quantifiable in humans than many of the other neurotransmitters and neuromodulators. It is therefore entirely plausible that the GABA changes observed after stroke and motor learning are due at least in part to modulation of the other neurotransmitter and neuromodulator systems, for example serotonin which has been shown to increase motor plasticity in animal models,68–70 as well as increases in cortical excitability and learning in healthy controls.71–73
Developing putative adjunct therapies for stroke recovery
The evidence presented above provides a sound rationale for developing potential adjunct therapies: by understanding the framework of likely neurophysiological changes underpinning natural behavioural improvements, we can drive these exogenously to maximize outcome. The rationale for any of these potential interventions would be that creating a “pro-plastic” neurochemical milieu during motor training would promote superior functional recovery. To this end, both pharmacological approaches and non-invasive brain stimulation (NIBS) approaches have been trialled. Pharmacological approaches are by their nature global interventions, but NIBS approaches have been targeted either to directly increase cortical excitability in the ipsilesional hemisphere or decrease cortical excitability in the contralesional hemisphere.
A number of putative adjunct therapies show promise in this context, with much of their promise coming from evidence of their effectiveness in healthy motor learning. We will therefore discuss these potential therapies here, before going on to summarise the evidence for their use in stroke. There is a wealth of literature from animal models, but the majority of this is outside the scope of this review.
NIBS
NIBS typically refers to both magnetic stimulation in the form of TMS, as discussed above, and electrical stimulation in the form of either transcranial direct- or alternating-current stimulation (tDCS and tACS respectively). All these NIBS techniques can be applied to a number of different anatomical locations in an attempt to enhance motor learning, but for the purposes of this review, we will focus on application to the primary motor cortices (M1).
Repetitive TMS in healthy motor learning
The utility of TMS as a method for quantifying synaptic activity has been discussed above. However, when a train of pulses are applied to the cortex at a set frequency or pattern, known as repetitive TMS (rTMS), it can also be used to induce plasticity (see Figure 2(c) for a schematic of the TMS approaches in common use). Low-frequency rTMS (typically 1 Hz) has been shown to cause a decrease in cortical excitability, 98 whereas high frequency rTMS (5–20 Hz) leads to cortical facilitation. 99 Theta burst stimulation (TBS) is a form of patterned TMS where three TMS pulses are delivered at 50 Hz, repeated every 200 ms (the theta frequency). TBS trains can be applied continuously (cTBS) or intermittently (iTBS) which respectively have inhibitory and excitatory effects on cortical excitability. 100
Attempts to facilitate motor learning using TMS typically aim to increase cortical excitability within the contralateral cortex, either by applying excitatory forms of rTMS locally, or by applying inhibitory rTMS to the ipsilateral cortex. It has been demonstrated that inhibitory 1 Hz rTMS applied to M1 ipsilateral cortex of the training hand, causes increases in excitability in the contralateral M1101–103 as well as decreases in intra-cortical inhibition, 102 and decreases in interhemispheric inhibition from the stimulated to non-stimulated hemisphere. 91 This same protocol, applying 1 Hz rTMS to ipsilateral cortex, also enhances key pressing task performance.104–106 Likewise, excitatory 10 Hz rTMS applied to M1 during a contralateral sequential finger task improved motor sequence learning. 107
Several studies using 1 Hz low frequency rTMS have shown no change in TMS measured intra-cortical inhibition,108–110 but one study found a decrease in 3 ms ISI SICI, 111 and another, using 0.6 Hz rTMS found increases in both 2 ms and 4 ms ISI paired-pulse measurements. 112 Further studies have also found decreases in intra-cortical facilitation, with 1 Hz rTMS.113,114 cTBS has been shown to increase MRS-assessed GABA. 115
Higher frequency rTMS appears to have more reproducible and significant effects on ppTMS measures, with several studies finding decreases in intra-cortical inhibition with 5 Hz rTMS.99,116–121 Fewer studies have examined intra-cortical facilitation, with most finding no change116,118,120,122 but one finding a facilitation. 119
It is not clear why these discrepancies exist, but the near infinite parameter space of rTMS, in terms of frequency, intensity and number of pulses applied, makes direct comparison between studies difficult. It is also important to point out that the literature studying the mechanisms of rTMS improvements in motor learning, rather than rTMS on its own, is to our knowledge sparse.
Repetitive TMS for stroke recovery
Beyond its use to influence motor learning in healthy controls, rTMS and TBS have been widely investigated for their potential role as therapeutic tools to enhance rehabilitation. The majority of studies have focused on improving motor function (particularly of the upper-limb); however, therapeutic promise has also been shown using rTMS and related techniques to improve aphasia, visuospatial neglect, 123 pain124,125 and swallowing disturbances.126,127
Specific to deficits of the motor system, there is still significant debate surrounding the optimal method for rehabilitating post-stroke interhemispheric imbalance is an ongoing debate, particularly in regards to modulating either ipsi- or contralesional motor regions (for a full review, see Nowak et al. 128 ). At its most simple, the debate falls between delivery of excitatory (high train rTMS or iTBS) stimulation to the affected hemisphere to augment residual activity or encourage vicarious changes to ipsilesional areas and delivery of inhibitory (low frequency rTMS or cTBS) stimulation to the unaffected hemisphere to decrease interhemispheric inhibition towards the lesioned side. Multiple studies have investigated the effects of inhibitory vs. excitatory protocols to each hemisphere within patient groups. Khedr et al. 129 tested both inhibitory (1 Hz) and excitatory (3 Hz) rTMS delivered to the unaffected and affected hemispheres, respectively, and found a greater improvement in the 1 Hz (contralesional) group on stroke impairment scales and simple tests of motor function. Takeuchi et al. 130 tested a similar inhibitory protocol delivered to the contralesional hemisphere compared to a 10 Hz excitatory protocol to the ipseileasional hemisphere, and a combined bihemispheric protocol. The authors found significant improvement in both the inhibitory and bihemispheric protocols. An additional study has gone on to find contralesional inhibitory rTMS leads to a decrease in fMRI measured overacitvation in the ipsilesional motor cortex. 131 Another recent study applied daily 5 Hz rTMS to the ipsilesional M1 and found improvements in a number of measured of upper-limb dexterity and strength measures. 132 Taken together, these findings show promise for the development of rTMS as a therapeutic tool for motor rehabilitation.
A very recent meta-analysis of the rTMS and motor stroke rehabilitation literature 133 suggests that both high and low frequency rTMS are safe and effective in stroke populations, and can facilitate short-term gains. However, the long-term effects of such interventions, which are a better measure of potential for rehabilitation, have been less widely researched. Interestingly, the authors observed a session number dependent effect, such that while single session studies were effective, the most beneficial results were observed in studies when multiple sessions were completed.
Within the TBS literature, somewhat contrasting findings have been reported. One study 134 found ipsilesional iTBS-enhanced M1 excitability, while contralesional cTBS resulted in decreased ipsilesional M1 excitability, correlating with reduced motor function (measured using the ARAT). Contrastingly, a separate study found cTBS to have no effect on the paretic hand, either electrophysiologically or behaviourally. 135 The authors did, however, find iTBS to the ipsilesional cortex-enhanced behaviour and increased MEP size in the stimulated hemisphere. Meta-analysis review 136 of the TBS literature suggests a more beneficial effect of iTBS for motor recovery rather than cTBS.
Despite various positive findings, it is important to consider the possibility that the heterogeneous lesion size and location within the cohorts studied may affect dispersion of the current within the ipsilesional hemisphere. 137 Furthermore, the effects of rTMS appear to be stronger in the acute patient population, compared to the chronic population. 136 This is not necessarily an unsurprising finding, as many interventions are more effective in the early stages of recovery, due to the enhanced cortical plasticity during that time. Nevertheless, the studies within the chronic population are encouraging and rTMS and TBS as potential motor rehabilitation will undoubtedly continue to be investigated.
On a final note, further considerations not yet mentioned include the optimal dosage of stimulation (duration and frequency) that should be delivered, and the combination of these stimulation protocols and forms of motor training. Functional recovery requires motor learning, and therefore the coupling of non-invasive stimulation techniques with physical practice should benefit from engaging the active brain state during stimulation.
Transcranial direct current stimulation in healthy motor learning
tDCS is the most commonly used electrical stimulation technique for influencing motor learning. Like TMS, tDCS can have different effects depending on exact parameters used, with electrode placement being the most commonly varied parameter. The typical montage used in motor learning studies involving training of the right-hand, places the anode electrode over left M1, and the cathode electrode on the right supraorbital ridge. Studies using this anodal montage, with stimulation applied during training have found enhancements in learning on several tasks including sequence learning,138–143 visuomotor adaptation144–146 and dexterity.147–149 In addition, tDCS has also been demonstrated to enhance offline learning across many of these same tasks, e.g. sequence learning139,140,143,150 and visuomotor adaptation. 151
tDCS is thought to exert its concurrent behavioural effect by influencing cortical excitability, with anodal stimulation to M1 causing an increase in MEP amplitude in the stimulated cortex. 152 Applying cathodal tDCS, where the electrode placement montage is reversed, causes a decrease in excitability within the stimulated cortex, but an increase in excitability in the non-stimulated cortex 153 in much the same way as inhibitory rTMS. These tDCS-induced changes in cortical excitability have been shown to outlast the stimulation period by some tens of minutes. 154
Anodal tDCS has been demonstrated to reduce SICI,155,156 and increase intracortical facilitation, 155 indicating that tDCS may be causing an increase in glutamatergic activity and a decrease in GABAergic. In line with this, work by Stagg et al. 157 has demonstrated that anodal tDCS over M1 causes a decrease in MRS-measured GABA levels in the stimulated area of cortex. Decreases in GABA have also been demonstrated when individuals practice a motor skill 158 and so a decrease in GABA has been proposed as a potential mechanism for tDCS learning effects. 159
Results from combined tDCS and drug studies paint a more complicated picture of the role of GABA. The benzodiazepine, lorazepam, causes a delay, in the onset of the beneficial effects of anodal tDCS; however, this late-onset effect is enhanced and prolonged.
160
NMDA receptor agonist D-cycloserine has also been shown to extend the duration of cortical excitability enhancements by tDCS.
161
Additionally, work has shown that the (SSRI) citalopram can enhance the beneficial effect of anodal tDCS on motor plasticity, and reverse the normally inhibitory effect of cathodal tDCS,
162
indicating that the serotonin system is also involved.
tDCS as a potential therapy post-stroke
Single sessions of ipsilesional anodal tDCS have consistently been demonstrated to improve motor performance in the short-term.171–176 One study which delivered anodal tDCS, paired with a nine-day course of upper-limb motor training found significant behavioural improvements that outlasted the stimulation period by at least three months. 177 The behavioural effects of cathodal tDCS to the contralesional hemisphere are less consistent, with this stimulation protocol being shown either to induce a modest behavioural gain, or no improvement.143,171,178 Bilateral stimulation, where the anode is placed over the ipsilesional M1 and the cathode over the contralesional M1 has also been shown to induce modest behavioural improvements. 179
Functional MRI studies show both anodal and cathodal tDCS lead to increased movement-related fMRI signal in the ipsilesional M1. The magnitude of the ipsilesional anodal tDCS-induced changes correlates with behavioural improvement on a subject-by-subject basis.171,177
The neurochemical underpinnings of these behavioural and neural changes are not understood, although it would seem intuitive to suggest that the mechanisms could be very similar to those underpinning stimulation-induced improvements in motor learning. Indeed, a recent study suggested that baseline GABA levels would predict subsequent behavioural response to anodal tDCS in stroke patients 180 (Figure 3(c)). However, without further direct investigation of the effects of tDCS – including assessment of neurotransmitter involvement (either using MRS or TMS) – on the stroke brain, we cannot rule out the possibility of other potential physiological mechanisms specific to the stroke population.
Pharmacological modulation
Pharmacological modulation of healthy motor learning
As well as using drugs to probe the neurochemical changes which occur during brain stimulation, there have also been some studies examining the effect of neuromodulatory drugs on motor learning alone, though they are less numerous. This is presumably because drugs that could theoretically be assumed to be candidates for enhancing motor learning, such as GABA-receptor antagonists and NMDA-receptor agonists, have potential proconvulsive and psychomimetic properties which made have non-specific, brain-wide effects.
However, although drugs that act on the major neurotransmitter systems have significant potential side-effects, the role of a number of neuromodulators in motor learning has been studied. In line with its effects on enhancing the effects of tDCS, the SSRI paroxetine has been shown to significantly increase practice-induced performance gains on the Nine Peg Hole Test. 163 Paroxetine also increased fMRI measured motor activation, 164 and the paroxetine-induced increase in speed in a finger tapping task has been shown to correlate with enhancement of ICF. 165 Similarly, D-amphetamine, which causes release of dopamine, norepinephrine and serotonin, has also been shown to enhance training-induced changes in TMS responses.166,167 Another study showed increasing norepinephrine levels using selective norepinephrine-reuptake inhibitor reboxetine caused improvements in gross motor skill acquisition; 168 however, the same effect was not found when examining finer motor skill learning. 169 Use-dependent plasticity is also attenuated by muscarinic acetylcholine receptor antagonist scopolamine. 170
Pharmacological manipulations post-stroke
As discussed above, modulating GABA or glutamate activity using pharmacological agents has clear potential side-effects and therefore has not been widely attempted in patients.
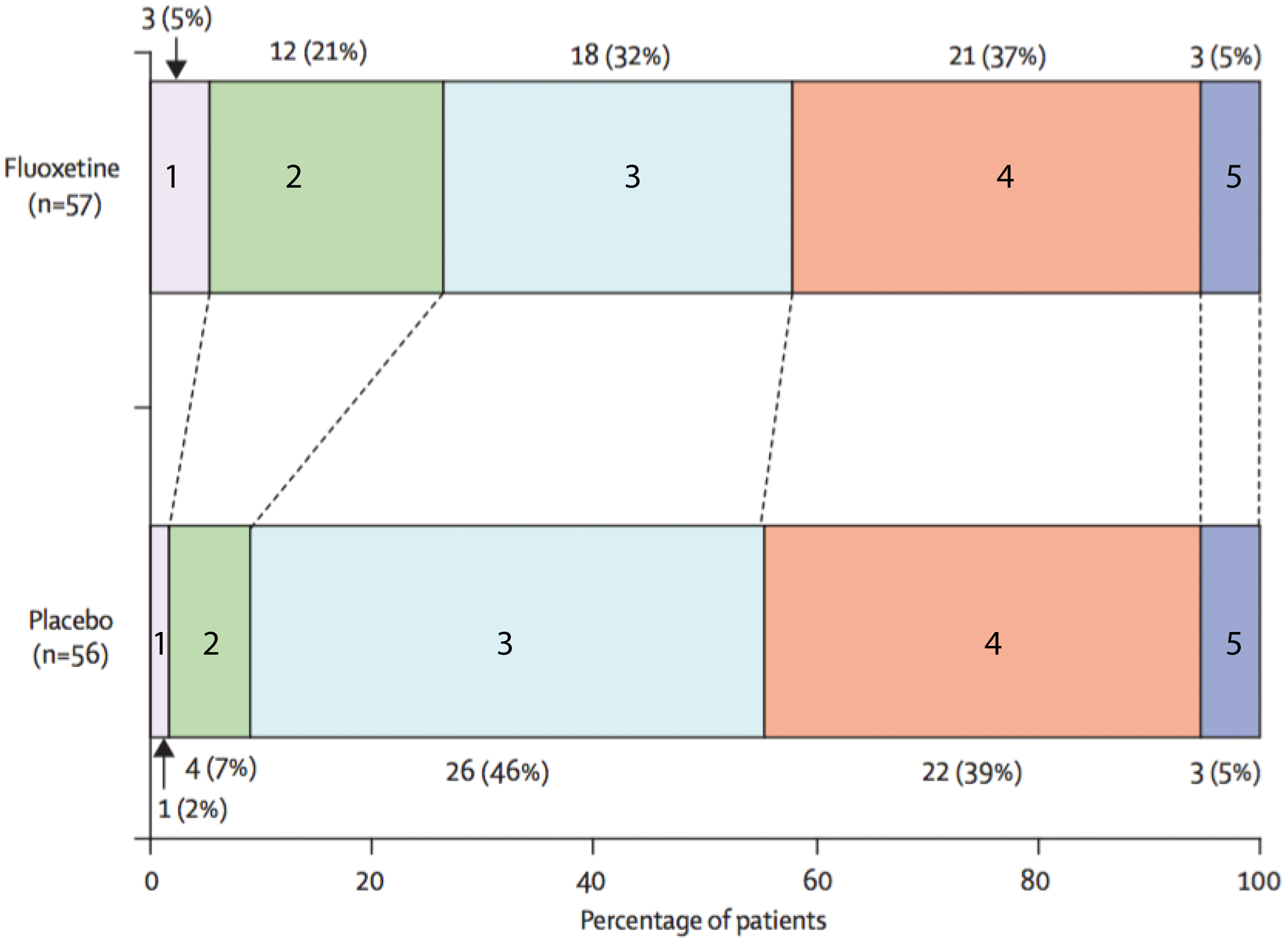
There is an increasing body of evidence that the SSRIs have a significant functional benefit in stroke patients, even in the absence of depressive symptoms74,75 (Figure 6). The evidence for positive benefits with other pharmacological interventions is still somewhat sparse, but this does not necessarily mean that directly manipulating neuromodulatory activity is ineffective for improving behavioural outcome. Trials to date have not paired a drug intervention with motor training, probably a crucial factor in maximising any potential pro-plastic changes.
Distribution of modified Rankin scores at 90 days post ischemic stroke for individuals on either fluoxetine or placebo. The modified Rankin score is a common outcome score in stroke trials and is an integer scale from 0 to 6, where 0 is “no symptoms at all”, 1 reflects no significant disability despite ongoing symptoms, 2 reflects a slight disability and so on with a score of 6 reflecting the death of a patient. Modified Rankin Scores are given on the coloured bars. Mean functional improvement was significantly higher in the fluoxetine group than in the placebo group, as can be observed by the higher numbers of patients with a score of 1 or 2 after 90 days. Figure adapted with permission from Chollet et al.
75
Future directions
We have summarised here the primary rationale for an interest in neurochemical changes in stroke as a theoretical framework for developing neuroscientifically informed neurorehabilitation interventions. We have described the neurochemical changes underpinning natural recovery after stroke, as far as they are currently understood, and then have discussed the most promising potential therapeutic interventions that are thought to directly affect neurochemical activity.
Is there evidence that neurochemical modulation may have a beneficial effect in stroke outcome? While studies are currently limited, high-quality evidence exists for the beneficial effects of the SSRIs to promote motor recovery, even in the absence of clinical depression. NIBS approaches, particularly tDCS, also to show promise as adjunct therapies in motor recovery after stroke when paired with physiotherapy. While the mechanism(s) underpinning functional benefits in patients is not yet clear, it seems likely that tDCS at least elicits functional benefits, at least in part through modulation of local GABAergic activity, a putative though as yet unproven, common pathway for both tDCS and the SSRIs.
There is convergent evidence that GABAergic activity plays a central role in facilitating functional recovery after stroke. Taken together, however, studies suggest that the pattern of inhibitory changes post-stroke are complex, and require further study to elucidate the best targets for the next generation of putative therapies. It is important to consider in this context that many stroke patients are prescribed GABAB agonists such as baclofen for the relief of spasticity – further work is needed to determine what effect this has, if any, on their capacity for post-stroke plasticity.
This review has focused on neurochemical changes in the ipsilesional motor regions. However, as discussed briefly above, it seems unlikely that anatomically distant regions that become recruited during movement post-stroke have no neurochemical changes, though these have been much less studied. The role of these regions is much debated, with conflicting evidence as to whether increased activity here is maladaptive or adaptive for recovery.6–8 Given the inherently limited potential for recovery of the ipsilesional cortex, these anatomically more distant regions provide increasingly important therapeutic targets, and therefore future work needs to be targeted on understanding their role in functional recovery.
Despite these limitations in the literature, it is to be hoped that future work will deepen our knowledge of the neurochemical changes after stroke, extend our conceptual framework, and allow for the development of more effective interventions to address this hugely under-met clinical need.
Footnotes
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.