
Abstract
Following intracerebral hemorrhage (ICH), extracellular heme precipitates secondary brain injury, which results in irreversible brain damage and enduring neurological deficits. Hemopexin (Hpx) is an endogenous protein responsible for scavenging heme, thereby modulating its intrinsic proxidant/proinflammatory properties. Although Hpx is present in the brain, the endogenous levels are insufficient to combat the massive heme overload following ICH. We hypothesized that increasing brain Hpx levels would improve ICH outcomes. Unique recombinant adeno-associated viral vectors were designed to specifically overexpress Hpx within the mouse brain. Western blotting, ELISA, and immunohistochemistry of brain homogenates/sections, CSF, and serum were performed. As compared to controls, Hpx mice have increased Hpx protein levels in all three types of biospecimens evaluated, which results in 45.6 ± 6.9% smaller lesions and improved functional recovery after ICH (n=14–19/group, p < 0.05). Local mechanistic analyses show significantly less tissue injury, trends toward smaller hematoma volumes, unchanged heme oxygenase 1 and iron levels, and significantly increased microgliosis and decreased astrogliosis and lipid peroxidation. Peripheral levels of heme-related markers indicate a positive modulation of iron-binding capacity. These findings reveal that high local Hpx levels improve ICH outcomes, likely through both central and peripheral clearance mechanisms, and establish the potential for therapeutically administering clinical-grade Hpx for ICH.
Introduction
Intracerebral hemorrhage (ICH) is a type of hemorrhagic stroke that affects 10–30 per 100,000 people worldwide each year and is associated with 30-day mortality rates as high as 35–52%. 1 Primary injury occurs early after the bleed from hematoma mass effects that cause mechanical disruption of neurons and glia. 2 Occurring later, secondary injury is largely due to the presence of blood components and their breakdown products that precipitate many parallel-operating neurotoxic processes, leading to irreversible brain damage and poor outcomes. 2 These processes include oxidative stress, inflammation, blood–brain barrier (BBB) breakdown, edema, oligemia, mitochondrial dysfunction, excitotoxicity, spreading depression, and cell death.1–3 Consequently, therapeutic paradigms aimed at detoxifying and improving the clearance of blood products would represent a clinically relevant treatment strategy for ICH.
Heme toxicity is a major contributor to secondary injury and consequently represents a possible preventable source of brain damage.4,5 Hemolysis after the bleed releases large quantities of deoxyhemoglobin.2,6 Deoxyhemoglobin is spontaneously and non-enzymatically oxidized to methemoglobin as the heme-iron is converted from ferrous to ferric form. 6 The toxic heme moieties quickly separate from methemoglobin, particularly in the presence of nitric oxide and reactive oxygen species, which exist at sites of inflammation such as the injured brain area after ICH. 7 Heme has several routes of toxicity, including the generation of superoxide and hydroxyl radicals, release of redox-active iron, depletion of cellular stores of NADPH and glutathione, peroxidation of membrane lipids, and sensitization of cells to subsequent noxious stimuli.4,7 Hemopexin (Hpx) is a glycoprotein that binds heme with the highest affinity among known proteins, thereby neutralizing its toxic properties. 7 The Hpx-heme complex is internalized by CD91/LRP1-mediated endocytosis where heme oxygenase enzymes degrade the heme.7,8 Hpx is present endogenously in the brain,9,10 although not at a level adequate to combat the heme toxicity load seen after ICH.
Using in vitro and in vivo approaches, the protective effects of Hpx have been reported in models of various disorders of the central nervous system (CNS).9,11–14 We have shown that genetic deletion of Hpx exacerbates outcomes following ischemic stroke. 9 Although it appears that Hpx is beneficial in these settings, the question remains as to whether high local Hpx levels reduce ICH-induced brain injury. The present study was designed to evaluate whether specific and local Hpx overexpression improves ICH outcomes, and if so, to investigate the potential in vivo mechanisms of Hpx-mediated neuroprotection.
Materials and methods
Mice
All animal procedures were approved by the University of Florida Institutional Animal Care and Use Committee, conducted in accordance with the National Institutes of Health PHS policy on Humane Care and Use of Laboratory Animals, and reported following ARRIVE guidelines. C57BL/6N mice were bred and maintained in our animal facilities in a temperature-controlled environment (23 ± 2℃) on a 12-h reverse dark/light cycle so that behavioral testing could be performed during the awake phase. The Hpx−/− mice included in this study for confirmation of anti-Hpx antibody specificity were originally generated by Dr. Emanuela Tolosano. 15 Mice were maintained on ad libitum food and water, including pre- and post-surgical procedures, and all efforts were made to minimize the possible suffering of the animals.
rAAV1 construction and preparation
Recombinant adeno-associated virus serotype 1 (rAAV1) vectors expressing eGFP, mouse Hpx (accession number BC019901), Hpx-eGFP, and Hpx-V5 under the control of the cytomegalovirus enhancer/chicken β-actin promoter, woodchuck post-transcriptional regulatory element, and bovine growth hormone poly(A) were generated as described. 16 Tags were fused to the C-terminus of the Hpx gene. The V5 tag used has the following amino acid sequence: GKPIPNPLLGLDST. The capsid serotype and timing of injection (see below) were selected such that transgene expression would be highest in regions surrounding the hematoma and disrupted the least by the ICH, thus resulting in sustained high local Hpx protein levels. 16 This approach results in predominately neuronal transduction, although some astrocytic transduction is also seen. 16 It should be noted that we used the exact same capsid 1 vector that was used in this cited report that characterizes the cellular and regional tropism of rAAV transduction in the mouse brain.
Prior to virus preparation, Hpx and control plasmids were independently validated by nucleotide sequencing performed at our institutional Interdisciplinary Center for Biotechnology Research. Viruses were prepared by polyethylenimine linear (PEI, Polysciences, Warrington, PA) co-transfection of the Hpx or control plasmid, and the AAV helper plasmid pDP1rs (Plasmid Factory, Germany) into HEK293T cells. At 72 h after transfection, cells were harvested and lysed in the presence of 0.5% sodium deoxycholate and 50 U/ml benzonase (Sigma, St. Louis, MO) by repeated rounds of freeze/thaws at −80℃ and 50℃. Viruses were isolated using a discontinuous Iodixanol gradient and samples were buffer exchanged to PBS using an Amicon ultra filter 100,000 MWCO centrifugation device (Millipore, Billerica, MA).
The genomic titer of each virus was determined by quantitative PCR using a CFX384 detection system (Bio-Rad, Hercules, CA). Briefly, viral DNA samples were prepared by treating the isolated virus with DNaseI (Life Technologies, Carlsbad, CA), heat inactivating the enzyme, digesting the protein coat with Proteinase K (Life Technologies), followed by a second heat inactivation. A standard curve of supercoiled plasmid diluted from 1 × 103 to 1 × 107 genomic equivalents/ml was used for comparison. Freshly prepared rAAV1s were aliquoted and stored at −80℃. When needed for injection, viruses were diluted to the same injection titer of 2 × 1013 genome equivalents/ml in sterile 1 × DPBS (pH 7.2) and used immediately.
rAAV1 injection
Neonatal rAAV1 injection procedures were adapted from a previous report. 17 For consistency in spatial transduction patterns, injections were performed within 12 h of birth. 16 C57BL/6N mouse pups were cryoanesthetized by placing them within an aluminum foil boat surrounded by ice for 3–4 min to reduce body temperature to <10℃, at which point the skin color visually changes from pink to purple and the pups are motionless so injections can easily and accurately be performed. 16 Bilateral intracerebroventricular injection of 2 µl of rAAV1 was performed using a 10-µl syringe with a 33-gauge needle and 30° bevel (Hamilton Company, Reno, NV) at a 45° angle to a depth of 1.5 mm. After injection, the needle was slowly retracted and the pups were placed on a heating pad to fully recover from cryoanesthesia and then returned to their mother and home cage. Mice were aged for 2.5–4.0 months at which point an ICH was surgically induced or brains were collected from naïve littermates to assess the level and localization of transgene-specific expression. Following neonatal rAAV1 transduction and recovery from cryoanesthesia, and throughout adolescent and adult life, all injected experimental groups were anatomically and behaviorally indistinguishable from the identical background wildtype (WT) breeding colony progeny used in this study (e.g. the no rAAV1 injection group, see below for experimental group specifics).
Randomization, exclusion, and blinding
Randomization in this study occurred with rAAV1 injection at birth. First, the total number of litters required for a sufficient number of male mice to use in this study was calculated, assuming a 50:50, male:female ratio, and a 20% addition was included to account for unforeseen loss (small litter size, skewed male:female ratio, cannibalism, etc.). Randomization of the particular rAAV1 to be injected (rAAV1-Hpx, rAAV1-Hpx-V5, rAAV1-Hpx-eGFP, rAAV1-eGFP) or no rAAV1 injection was performed by a single person not otherwise involved in the study and who had no knowledge of or contact, including no visual inspection, with the litters. Exclusion criteria was defined a priori and included any mouse with apparent abnormal conditions (skin, eye, abdominal, whisker, signs of infection, etc.), ear puncture during placement in the stereotactic device, blood reflux at any point during the autologous blood infusions, and bleeding on needle insertion (which precludes the ability to detect blood reflex). No mice were excluded from this study. Mice in different experimental groups were visibly indistinguishable (anatomically and behaviorally), thus blinding was incorporated throughout the study. The surgeon and investigators performing neurobehavioral testing had no knowledge of the experimental groups. Additionally, all anatomical outcomes were quantified in a blinded manner. In all cases, a unique numbering code was used with a linking list to the experimental treatment group and individual animal.
Neuronal-glial mixed primary cultures and rAAV1 transduction
To confirm the cellular versus secretory localization of our rAAV1-Hpx, rAAV1-Hpx-V5, rAAV1-Hpx-eGFP, and rAAV1-eGFP vectors, we utilized primary neuronal-glial cultures prepared as described.18,19 Briefly, cerebral cortices from P0 mouse brains were dissected and dissociated in 2 mg/ml papain (Worthington, Lakewood, NJ) and 50 μg/ml DNAase I (Sigma) in sterile Hank's Balanced Salt Solution (HBSS, Life Technologies) at 37℃ for 20 min. To inactivate the papain, they were washed three times in sterile HBSS and then switched to Neurobasal-A (Gibco, Waltham, MA) plating media containing 1% fetal bovine serum (HyClone, Logan, UT), 0.5 mM L-glutamine (Gibco), 0.5 mM GlutaMax (Life Technologies), 0.01% antibiotic-antimycotic (Gibco), and 0.02% SM1 supplement (Stemcell, Canada). The tissue mixture was then titrated three times using a 5-ml pipette followed by a Pasteur pipette and strained through a 70-μm cell strainer. Following centrifugation at 200 × g for 3 min, cells were resuspended in fresh plating media and plated onto poly-D lysine 6-well plates at around 100,000–200,000 cells/cm2. The following day, the media was replaced with maintenance media consisting of plating media without fetal bovine serum. Cells were maintained for 7 days with maintenance media prior to transduction with rAAV1s.
After a fresh media change, 5 μL of rAAV1-Hpx, rAAV1-Hpx-V5, rAAV1-Hpx-eGFP, or rAAV1-eGFP was added directly to the media. Wells with no rAAV1 transduction were included as negative controls. The media was not changed after this point. Three days later, images of the cultures were obtained using an EVOS FL cell imaging system (ThermoFisher Scientific, Waltham, MA). The media was then removed, centrifuged to remove any debris, and the supernatant, later referred to as media, was saved and stored at −80℃ until later immunoblotting. The cells were collected and washed in PBS prior to lysing for 15 min on ice with 1% Triton X-100 containing a protease inhibitor cocktail (Roche, Indianapolis, IN). The lysate was centrifuged at 15,000 × g and 4℃ for 30 min and the supernatant, later referred to as lysate, was saved and stored at −80℃ until later immunoblotting. After the initial plating and up until harvesting, cells were kept at 37℃ in a humidified 5% CO2 chamber.
ICH model
ICH was induced in male mice using the autologous whole blood double infusion model (30 µl total infusion). 20 Mice were anesthetized with isoflurane (4% induction, 1.5–2% maintenance) and immobilized on a stereotactic frame (Stoelting, Wood Dale, IL). After making a small midline sagittal incision in the skin overlying the skull, a craniotomy was performed 0.5 mm anterior and 2.4 mm right relative to the bregma. Autologous blood was collected onto a sterile surface by needle prick of the tail artery after first cleaning the area with 70% ethanol and warming the tail gently for 2 min with a heat lamp. Blood was immediately drawn into PE-20 tubing (Instech, Plymouth Meeting, PA) connected on one side to a 50-µl syringe with a 26-gauge luer tip needle (Hamilton Company) located within an automated injector, and the other side to a 26-gauge needle with the bevel end inserted into the tubing. The blunt end of this needle was inserted 3.9 mm ventral from the skull surface, removed to 3.6 mm, and left in place for 7 min; 10 µl of blood was infused, followed by a 5-min waiting period prior to the second infusion of 20 µl. All injections were performed at 1.0 µl/min using an automated injector (Stoelting). The needle was left in place for 10 min after the second infusion prior to slow removal over a 25-min period. Rectal temperatures were maintained at 37.0 ± 0.5℃ throughout all surgical procedures and mice were allowed to fully recover in temperature- and humidity-controlled chambers postoperatively.
The control mice (total n=18) include rAAV1-eGFP (n=7) and no rAAV1 injection (n=11), and the two groups were combined for statistical comparisons because no differences were observed. The experimental groups (total n=23) consisting of rAAV1-Hpx (n=8), rAAV1-Hpx-V5 (n=6), and rAAV1-Hpx-eGFP (n=9) were similarly combined and herein referred to as Hpx mice.
Functional outcomes
Two blinded investigators independently assessed the mice for focal neurological deficits daily post-ICH by neurological deficit scoring (NDS) as described.21,22 Testing was performed during the dark cycle (awake phase) at the same time each day. Briefly, a score of 0 (no deficits) to 4 (severe deficits) was assigned for six individual parameters, including body symmetry, gait, circling behavior, climbing, front limb symmetry, and compulsory circling. NDS is reported as the average of the sum of the individual scores for the two investigators.
Tissue and biospecimen harvesting
For those mice that underwent ICH, all collection procedures occurred at 72 h after surgery and were performed sequentially on the same mice. First, mice were maintained under isoflurane anesthesia and a maximal volume of CSF was extracted from the cisterna magna with careful avoidance of the dorsal spinal artery as described with slight modification.23,24 Rather than using a capillary tube, extraction was performed with a 10-µl pipette equipped with a fine-ended tip after puncturing the dura with a 25-gauge needle, and CSF was stored at −80℃ for later analysis. Blood was then obtained by cardiac puncture and kept on ice for 30 min prior to centrifugation at 1500 × g for 15 min. Serum was collected and stored at −80℃ for later analysis. After blood collection, mice were transcardially perfused with PBS followed by 4% paraformaldehyde. Brains were collected and kept in 4% paraformaldehyde for 24 h prior to cryopreservation in a 30% sucrose/PBS solution for subsequent histology.
Naïve littermates that received the same rAAV1 injections (and at the same time) as those mice that underwent ICH were used for confirmation of transgene-specific expression. Brain tissue was harvested after deep anesthetization with isoflurane and PBS perfusion. The cerebellum and olfactory bulbs were removed and brains were snap frozen in pre-cooled 2-methylbutane and stored at −80℃ for subsequent homogenization.
Liver tissue positive and negative controls were analogously harvested, processed, and stored after collection from naïve WT and Hpx−/− mice, respectively, which did not receive an rAAV1 injection.
Western blotting
Sodium dodecyl polyacrylamide gel electrophoresis and Western blotting of in vivo and in vitro samples was performed to characterize and localize rAAV1-mediated Hpx overexpression. Brain and liver tissue were homogenized and protein content was subsequently estimated using the bicinchoninic assay (ThermoFisher Scientific). Serum and CSF were loaded at a constant volume. Sample isolation for the in vitro experiments is described above and the media and lysate protein concentration was estimated in an identical manner to the tissue homogenates. The supplemental material and Table S1 have additional methods and details on the antibodies used.
Measurement of heme-related serum markers following ICH
ELISA was used to determine serum levels of Hpx and haptoglobin according to the manufacturer instructions (Life Diagnostics, West Chester, PA). Bilirubin (Sigma) and iron (Pointe Scientific, Canton, MI) levels were determined by standard colorimetric assays per manufacturer instructions.
Histology and quantification
Histological staining and quantification procedures were performed by blinded investigators as we have described21,22; 10 sets of 16 sections equally distributed throughout the entire hematoma and anteroposterior brain regions were processed on a CM 1850 cryostat (Leica Biosystems, Buffalo Grove, IL) at 30 µm and stored at −80℃ for later histological procedures. In this way, for each animal, multiple staining procedures can be performed and the staining pattern throughout the whole brain can be analyzed. Cresyl violet staining was used to assess lesion volume, perihematomal tissue injury, and hematoma volume. Perls' iron staining was performed to evaluate iron content. The antibodies used for immunohistochemistry to evaluate the localization of rAAV1 expression, HO1 expression, lipid peroxidation, astrogliosis, and microgliosis are provided in Table S1. All slides were scanned using an Aperio ScanScope CS and analyzed with ImageScope software (Leica Biosystems).
Quantification was performed in a blinded manner, and to reduce any potential bias and interindividual variability, for a given histological stain, all slides simultaneously underwent the staining protocol and a single investigator performed the quantification. For quantification procedures in which total brain pathology was analyzed (lesion volume, perihematomal tissue injury, hematoma volume, ferric iron content, and HO1 expression), all 16 sections were quantified for each animal. 4-HNE was evaluated on the two sections for each animal representing maximal lesion area. To assess astrogliosis and microgliosis, four sections for each animal representing maximal lesion area were analyzed. Lesion volume: injured brain regions were outlined, areas abstracted from the ImageScope software, and a volume was calculated using these areas, known distance between each section, and section thickness. Injured brain areas are defined as the hematoma and perihematomal tissue injury/cell death as shown in Figure S1. For all other quantification procedures (hematoma volume, perihematomal tissue injury, iron content, and immunohistochemical stains), an ImageScope Positive Pixel Count algorithm was used for quantification after the appropriate brain regions were outlined (see below). Each algorithm was tuned for each of the individual stains such that the appropriate signal and strength of signal were evaluated. 21 Thresholds were set intermediate between the signals seen in the two experimental groups on a representative slide in each group such that the algorithm allowed for optimal detection in either direction (i.e. more intense versus less intense). After running an algorithm, all slides were checked for specificity and accuracy and to ensure minimal interference from artifact before the signal data was abstracted from the ImageScope software. Hematoma volume: the injured brain regions described above were outlined. A volume was calculated in an identical manner to that described for lesion volume quantification. Perihematomal tissue injury: Using the calculated lesion and hematoma volumes, the amount of perihematomal tissue injury was calculated by subtracting the hematoma volume from total lesion volume. HO1, iron, and 4-HNE: the injured brain regions and surrounding areas were outlined. Microgliosis and astrogliosis: cortical gliosis was analyzed by placing identically sized boxes of 1000 × 1000 pixels in the ipsilateral and contralateral cortex. Striatal gliosis was analyzed by outlining of the ipsilateral and contralateral striatum, excluding the lesion area, and these data are presented as the signal per area quantified. Because gliosis differences were noticed in the contralateral hemisphere between treatment groups, ipsilateral data were not normalized for contralateral signal; instead, the data are presented separately for comparison. After all analyses, the appropriate algorithm was run and signal data were abstracted from the ImageScope software.
Statistics
Statistical analyses were performed using Prism 6 (GraphPad, San Diego, CA). Mortality and NDS were analyzed using a χ2 test and non-parametric Mann–Whitney U test, respectively. The remaining data sets were checked for differences in variances between groups and normality and the appropriate statistical test was used, either a Mann–Whitney U test or an unpaired two-tailed Student's t test with or without Welch's correction. Data are expressed as mean±SEM with p < 0.05 considered statistically significant.
Results
Characterization of rAAV1 expression
The secretory nature of the rAAV1-Hpx-(tag) protein products was validated with in vitro transduction of mixed neuronal-glial cell cultures and Western blotting (Figure 1). Furthermore, these results confirm that the rAAV1-Hpx-V5 and rAAV1-Hpx-eGFP protein products are indeed fusion proteins.
In vitro characterization of rAAV1 expression. (a) Brightfield, fluorescent (GFP), and overlay images of mixed primary neuronal-glial cell cultures 72 h after no rAAV1 transduction or transduction with rAAV1-eGFP, rAAV1-Hpx-eGFP, rAAV1-Hpx-V5, or rAAV1-Hpx are shown. As expected, no fluorescence is seen for the no rAAV1 transduction negative control and the rAAV1-Hpx-V5 and rAAV1-Hpx transduced cultures. Transduction with rAAV1-eGFP resulted in intense intracellular fluorescence throughout neurons, whereas transduction with rAAV1-Hpx-eGFP had a significantly different pattern, with fluorescence restricted to perinuclear areas of neurons, consistent with a secretory protein cellular localization (white arrows). Furthermore, the comparatively low fluorescent intensity is consistent with the expected low intracellular levels of transgene-derived Hpx protein. (b) To verify the culture image results, the intracellular versus secretory distribution of the rAAV1 vector protein products was confirmed by Western blotting of culture media (“M”) and cell lysates (“P”) (6 μg). A near-infrared fluorescence Odyssey detection system was used such that the relation between Hpx and tags could simultaneously be shown, using the rAAV1-Hpx-eGFP vector as a convenient example. Hpx is identified with green (800) and eGFP is identified with red (680). The Hpx-eGFP protein product appears yellow on overlay images of culture media and cell lysates, confirming that the expressed Hpx-eGFP is a protein fusion. The culture media from rAAV1-Hpx-eGFP, rAAV1-Hpx-V5, and rAAV1-Hpx transduction shows a substantial amount of Hpx-eGFP, Hpx-V5, and Hpx protein, which directly confirms the secretory nature of these rAAV1 vector protein products. This finding is not as a result of potential cross-reactivity with Hpx present in fetal bovine serum because cultures were maintained in serum-free maintenance media. Further confirmation of specificity is provided by the lack of immunoreactivity in the no rAAV1 transduction lanes. A small amount of transgene-derived Hpx protein is also found in the cell lysates (confirming the low fluorescent intensity seen in culture images), which corresponds to protein being actively synthesized and post-translationally modified prior to secretion. In comparison, rAAV1-eGFP transduction results in an intracellular eGFP protein, as no eGFP is seen in the culture media. Last, after rAAV1 transduction, all protein products run at the expected molecular weights, with eGFP at 30 kDa, Hpx at 70 kDa (running equivalent to endogenous liver and brain Hpx), Hpx-V5 is shifted slightly higher than Hpx due to the small tag size, and Hpx-eGFP at 100 kDa.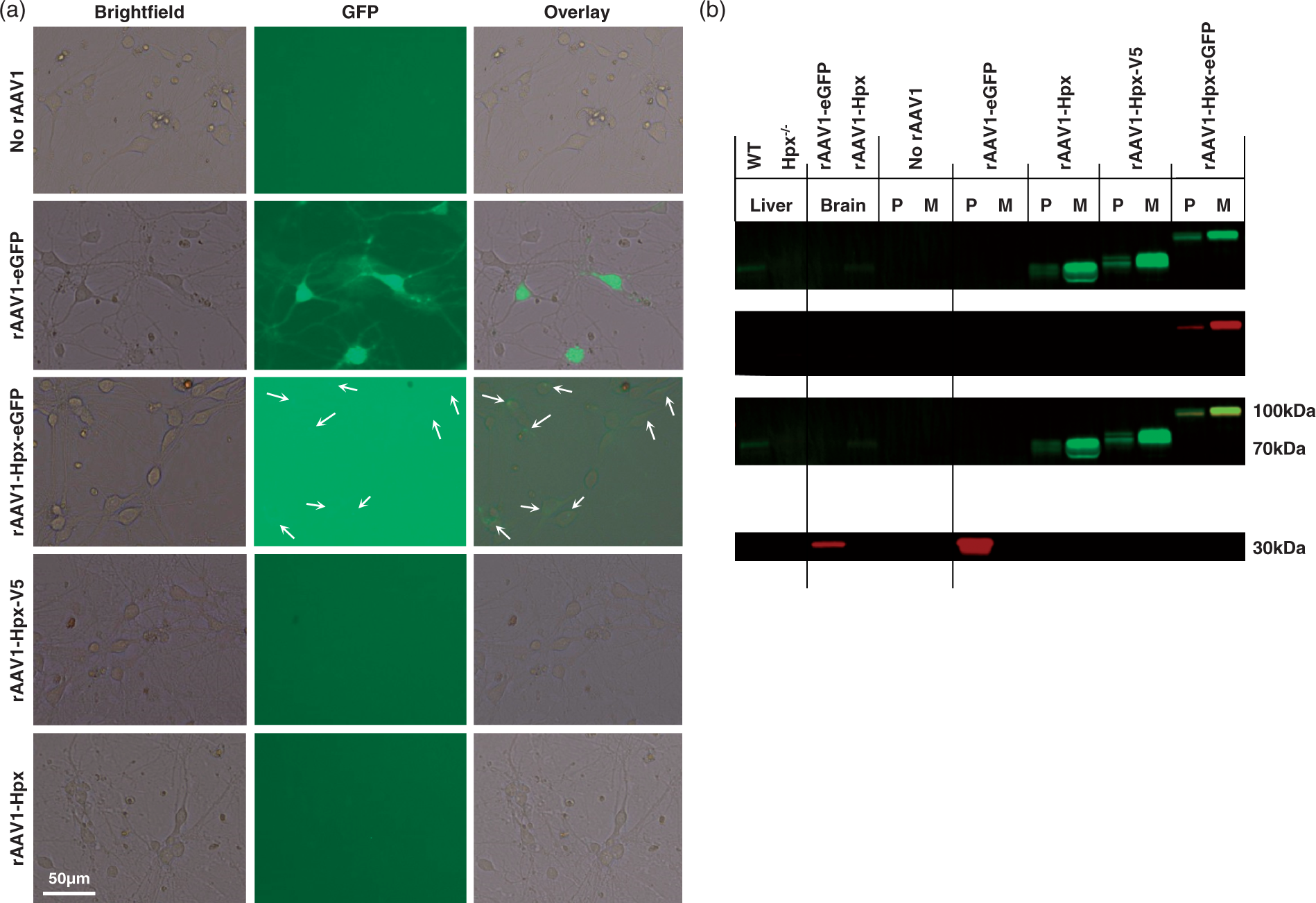
To demonstrate in vivo transgene spatial expression, immunohistochemistry for GFP was performed using sections from rAAV1-eGFP mice. The rAAV1 expression is primarily neuronal and is observed in cortical, striatal, thalamic, and hippocampal brain regions, as well as in the corpus callosum, internal capsule, several other white matter tracts, and periventricular areas (Figure 2(a)). No GFP staining is seen in the negative control mice that did not receive an rAAV1 injection (Figure 2(b)). Western blotting was performed to characterize the level of Hpx protein in control and Hpx mice. As compared to both control groups, naïve rAAV1-Hpx mice have an estimated 60-fold higher Hpx level in brain homogenates (Figure 2(c)). No difference is seen between the two control groups, indicating that in vivo rAAV1 transduction does not alter endogenous Hpx levels. CSF and serum from rAAV1-Hpx-eGFP mice that underwent ICH also shows high levels of transgene-derived Hpx (Figure 2(c)), confirming the in vivo secretory nature of the rAAV1-Hpx-(tag) protein products.
In vivo characterization of rAAV1 expression. (a) Representative images showing the reproducible spatial localization of GFP immunoreactivity in two separate rAAV1-eGFP-expressing mice. Numbered boxes on whole brain images denote the location of high magnification images in the quadrants, where the left and right quadrants correspond to the anterior and posterior coronal sections, respectively. (b) Representative images of a no rAAV1 injection negative control showing no GFP immunoreactivity. (c) Western blotting of brain homogenates (40 µg) from naïve control (rAAV1-eGFP and no rAAV1 injection) and rAAV1-Hpx mice was performed to evaluate the relative Hpx protein levels. Liver homogenates (10 µg) from WT and Hpx knockout (Hpx−/−) mice were used as positive and negative controls, respectively, to confirm antibody specificity. Actin was used as a loading control. Western blotting of 3 µl of CSF and 1.5 µl of serum obtained from control and rAAV1-Hpx-eGFP mice that underwent ICH was performed to identify the biocompartmental localization and confirm the in vivo secretory nature of the transgene-derived Hpx protein. The observed molecular weight for untagged Hpx and Hpx-eGFP was 70 and 100 kDa, respectively.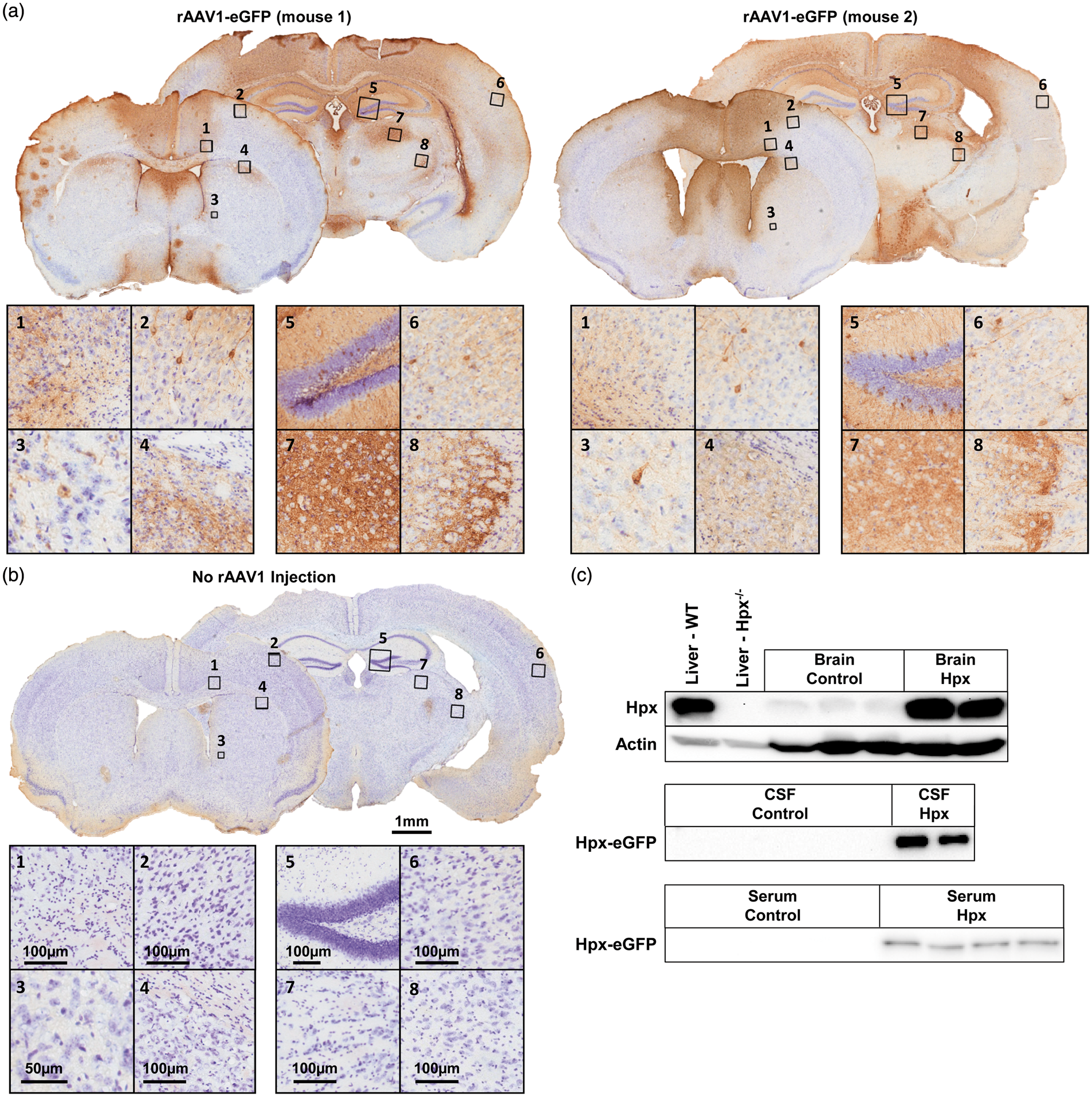
High local Hpx levels improve anatomical and functional outcomes following ICH
Hpx mice have significantly smaller lesion volumes associated with less tissue injury and trends toward reduced hematoma volumes (Figure 3(a)). Less ICH-induced brain injury in Hpx mice is accompanied by improved neurologic function (Figure 3(b)). Quantification reveals that Hpx mice have 45.6 ± 6.9% smaller lesion volumes (5.3 ± 0.7 mm3 vs. 9.7 ± 2.0 mm3, p = 0.0449; Figure 3(c)). Perihematomal tissue injury is reduced by 42.5 ± 9.8% in Hpx mice (3.5 ± 0.4 mm3 vs. 6.6 ±1.3 mm3, p = 0.0449; Figure 3(c)), and a trend toward reduced residual blood volume is also seen (1.8 ±0.3 mm3 vs. 3.1 ± 0.8 mm3, p = 0.1334; Figure 3(c)).
High local Hpx levels improve anatomical and functional outcomes following ICH. (a) Representative cresyl violet-stained brain images are shown for control and Hpx mice. The black outline signifies the lesion area, which consists of the hematoma (red area) and the surrounding injured brain tissue. Inserts are provided to show the reduced tissue injury/perihematomal cell death seen after ICH with high local Hpx levels. (b) Hpx mice have less focal neurological deficits after ICH. (c) Quantification reveals that Hpx mice have significantly smaller lesion volumes associated with less tissue injury and trends toward smaller hematoma volumes. Hpx mice also have significantly less body weight loss after ICH. Statistical comparisons include n=14–19 mice/group. *p < 0.05.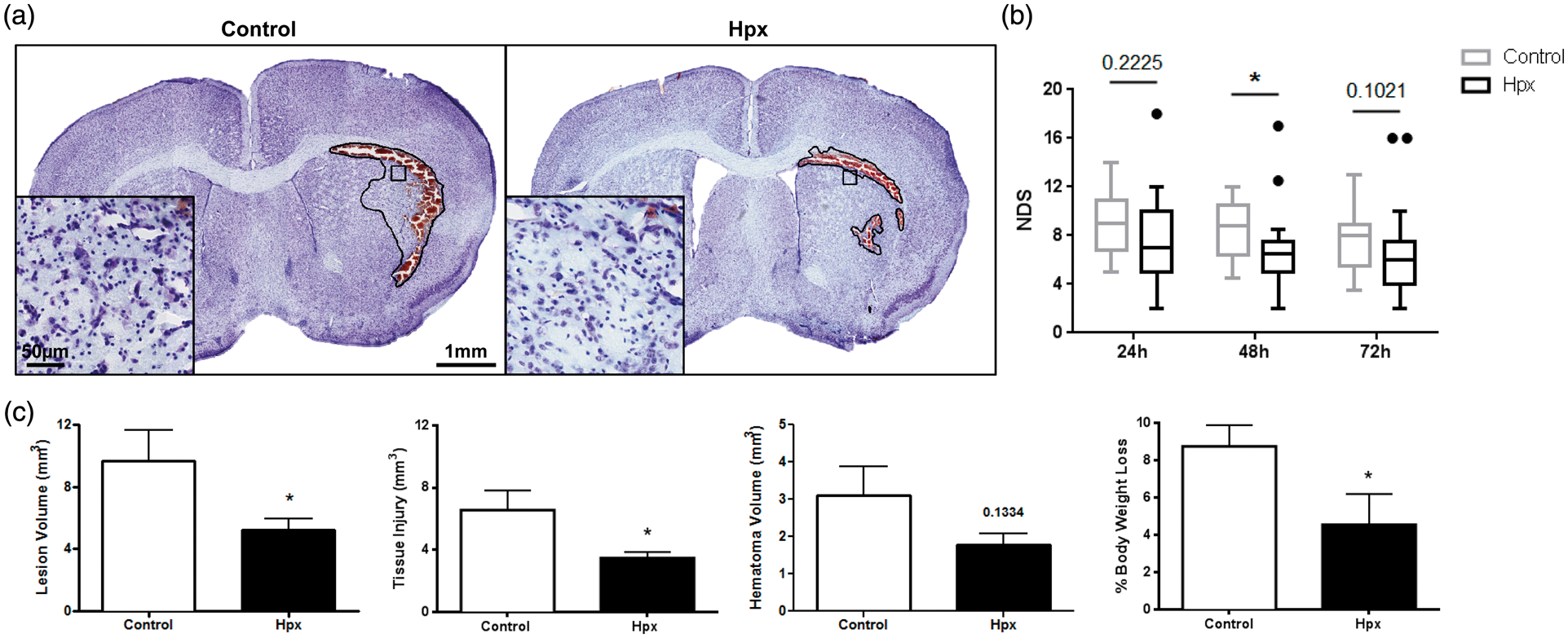
Prior to inducing an ICH, no difference in body weight is seen between groups (Hpx: 26.4 ± 0.5 g, control: 26.3 ± 0.4 g, p = 0.8607). At 72 h post-ICH, Hpx mice have 48.0 ± 18.8% less percent change in body weight (4.5 ± 1.6% vs. 8.7 ± 1.1%, p = 0.0499; Figure 3(c)). No difference in mortality is seen between the groups (Hpx: 21.7%, control: 22.2%, p = 0.9704). In all cases, death occurred during the second blood infusion as a result of respiratory depression and subsequent cessation.
Increased Hpx levels modulate serum levels of heme-related markers after ICH
Serum levels of heme-related markers.
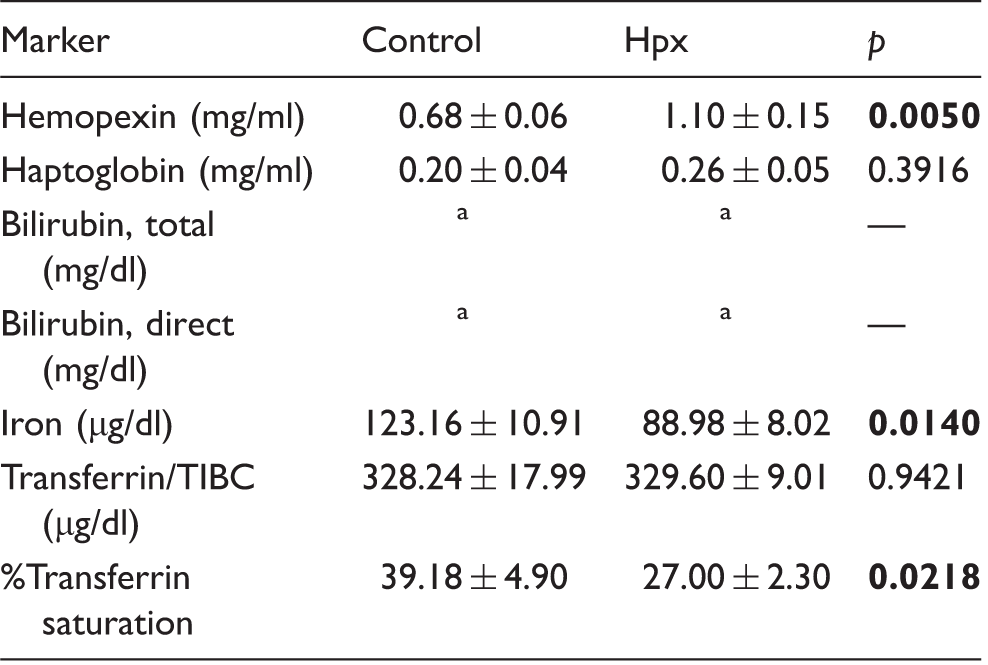
Below detection limit (values are < 0.2 mg/dl).
Effect of high local Hpx levels on HO1 and iron levels after ICH
Histology was used to start identifying local mechanisms of Hpx-mediated neuroprotection following ICH. To begin, immunohistochemistry for HO1 and Perls' iron staining was performed to evaluate the relative level and localization of HO1 and iron in control and Hpx mice (Figure 4(a) and (b)). Quantification reveals no difference in HO1 (Hpx: 6.4 ± 2.3 × 106A.U., control: 7.0 ± 2.4 × 106A.U., p = 0.8610; Figure 4(a)) or iron (Hpx: 2.8 ± 0.7 × 106A.U., control: 2.0 ± 0.2 ×106A.U., p = 0.2890; Figure 4(b)) between groups.
Effect of high local Hpx levels on lipid peroxidation and HO1 and iron levels after ICH. (a) Representative images showing the localization and relative level of HO1 immunoreactivity for control and Hpx mice. Left and right high magnification images correspond to the leftmost and rightmost square selections on the low magnification hemi-brain images, respectively. HO1 expression is clearly evident in perihematomal microglia/macrophages as well as glia more distant from the lesion and endothelial cells. Quantification shows no significant difference in HO1 levels between groups. (b) Representative images showing the localization and relative level of iron (blue) for control and Hpx mice. Square selections denote the location of magnified regions. After quantification, no significant difference in iron levels is seen between groups. (c) Representative images showing the relative level and localization of lipid peroxidation (4-HNE immunoreactivity) for control and Hpx mice. Square selections denote the location of magnified regions. Quantification reveals that Hpx mice have significantly less lipid peroxidation. (a–c) Inserts are placed in the perihematomal region. Statistical comparisons include n=10–14 mice/group. *p < 0.05.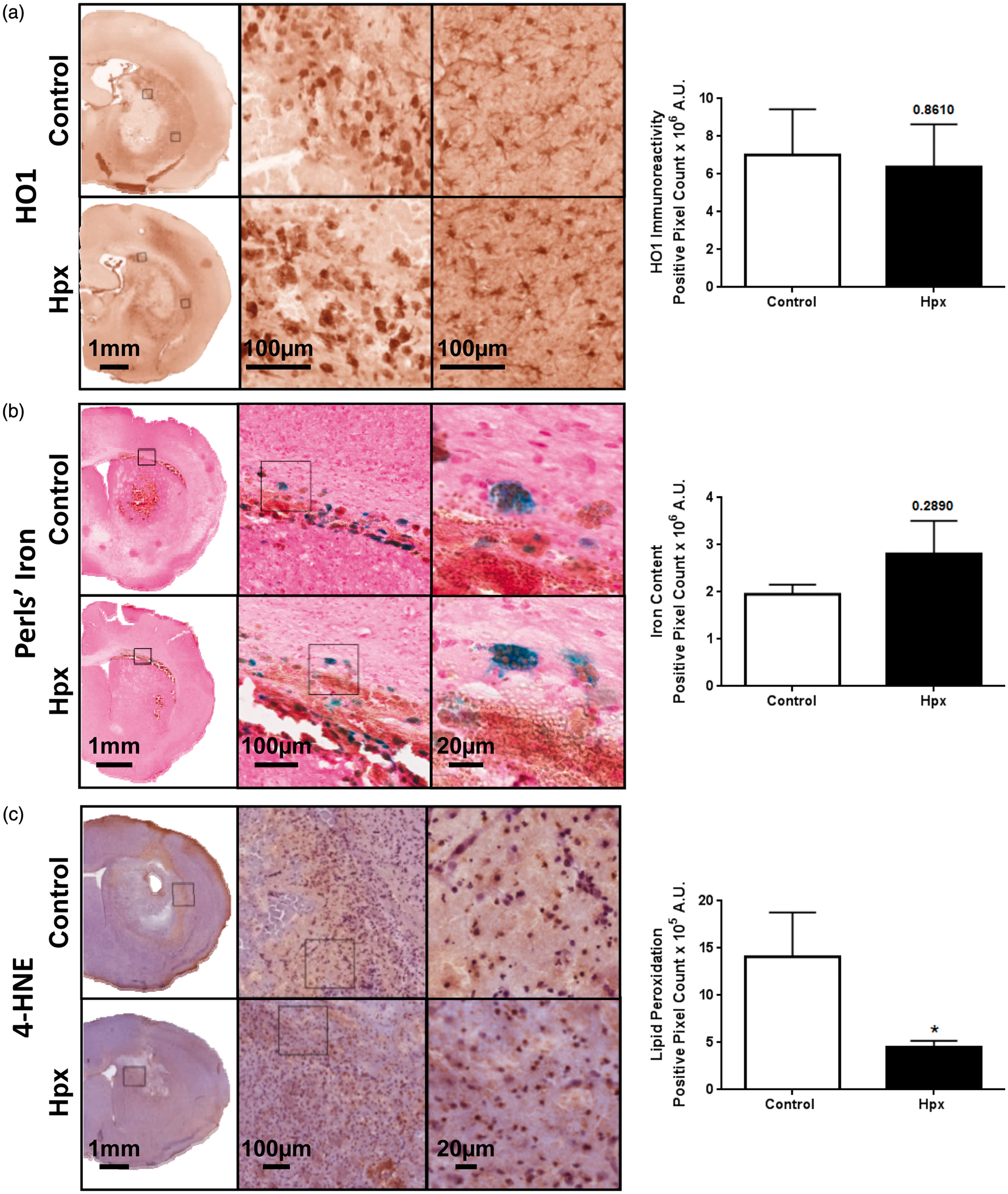
High local Hpx levels decrease lipid peroxidation following ICH
Immunohistochemical staining for 4-HNE was performed to evaluate the relative degree of lipid peroxidation after ICH. Immunoreactivity is observed primarily in the perihematomal region (Figure 4(c)). Quantification reveals that Hpx mice have 68.0 ± 4.7% less lipid peroxidation (4.5 ± 0.7 × 105A.U. vs. 14.1 ± 4.7 × 105A.U, p = 0.0431; Figure 4(c)).
High local Hpx levels decrease astrogliosis following ICH
GFAP immunohistochemistry was performed to assess astrogliosis. Overall, Hpx mice have less astrocyte activation and fewer morphological changes in both the ipsilateral and contralateral hemispheres (Figure 5(a)). After quantification (Figure 5(b)), no significant difference in cortical ipsilateral (Hpx: 3.7 ± 0.3 × 10−2A.U., control: 4.8 ± 1.0 × 10−2A.U., p = 0.5780) or contralateral (Hpx: 0.7 ± 0.2 × 10−2A.U., control: 2.0 ± 0.6 ×10−2A.U., p = 0.2654) astrogliosis is seen between groups. Hpx mice have 51.0 ± 5.8% less contralateral striatal astrogliosis (1.2 ± 0.1 × 10−2A.U. vs. 2.5 ±0.5 × 10−2A.U., p = 0.0346), and tend to have less ipsilateral striatal astrogliosis (4.9 ± 0.3 × 10−2A.U. vs. 5.7 ± 0.6 × 10−2A.U., p = 0.2371).
High local Hpx levels decrease astrogliosis following ICH. (a) Representative images showing characteristic morphological changes and differences in GFAP immunoreactivity in the ipsilateral and contralateral cortex and striatum for control and Hpx mice. Square selections in the inserts denote the location of magnified regions. Striatal quantification represents the perihematomal region, and thus is a quantification of astrogliosis from cellular activation in response to local processes (e.g. heme clearance). Cortical quantification was performed due to its connection with the site of injury (striatum) and involvement in sensory and motor neurologic function. (b) Quantification reveals a decrease in cortical and striatal astrogliosis for Hpx mice in the ipsilateral and contralateral hemispheres. Statistical comparisons include n=7–10 mice/group. *p < 0.05 and results above bars represent comparisons between control and Hpx mice. ##p < 0.01, ###p < 0.001, and ####p < 0.0001.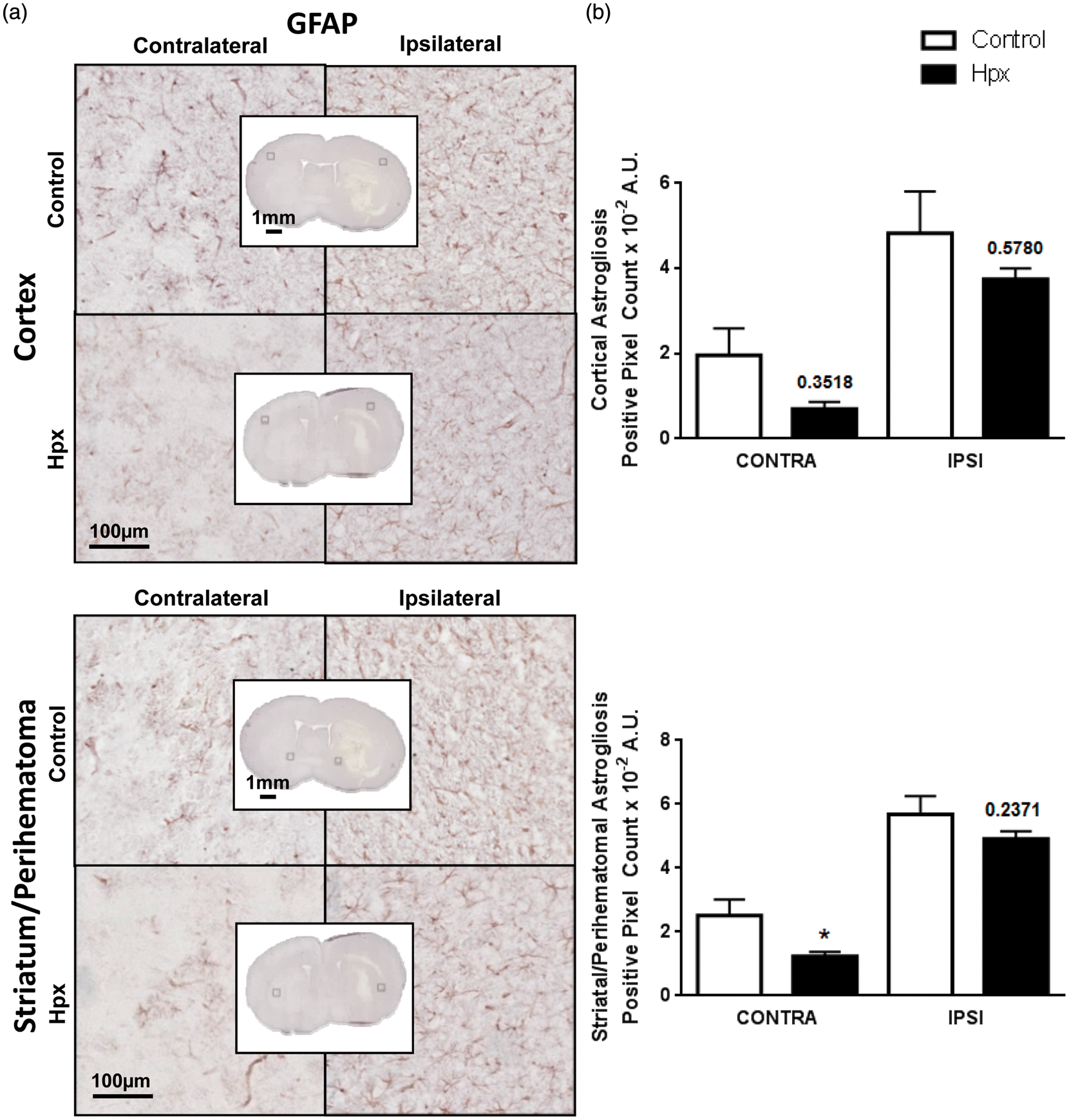
High local Hpx levels increase microgliosis following ICH
Iba1 immunohistochemistry was performed to assess microgliosis. Overall, Hpx mice have more microglial activation and morphological changes compared to controls in both the ipsilateral and contralateral hemispheres (Figure 6(a)). After quantification (Figure 6(b)), Hpx mice show 1176.0 ± 351.0% and 793.3 ± 224.2% more ipsilateral cortical (12.8 ±3.5 × 10−2A.U. vs. 1.2 ± 0.2 × 10−2A.U., p = 0.0163) and striatal (8.9 ± 2.2 × 10−2A.U. vs. 2.8 ± 0.4 ×10−2A.U., p = 0.0004) microgliosis, respectively. Hpx mice also have 155.6 ± 87.5% and 299.9 ± 115.6% increased contralateral cortical (2.6 ± 0.9 × 10−2A.U. vs. 0.8 ± 0.1 × 10–2A.U., p = 0.0858) and striatal (4.0 ± 1.2 × 10−2A.U. vs. 1.4 ± 0.3 × 10−2A.U., p =0.0068) microgliosis, respectively.
High local Hpx levels increase microgliosis following ICH. (a) Representative images showing characteristic morphological changes and differences in Iba1 immunoreactivity in the ipsilateral and contralateral cortex and striatum for control and Hpx mice. Square selections in the inserts denote the location of magnified regions. Striatal quantification represents the perihematomal region, and thus is a quantification of microgliosis from cellular activation in response to local processes (e.g. heme clearance). Cortical quantification was performed due to its connection with the site of injury (striatum) and involvement in sensory and motor neurologic function. (b) A significant increase in cortical and striatal microgliosis for Hpx mice was noted in the ipsilateral and contralateral hemispheres. Statistical comparisons include n=7–10 mice/group. *p < 0.05, **p < 0.01, ***p < 0.001, and results above bars represent comparisons between control and Hpx mice. #p < 0.05 and ##p < 0.01.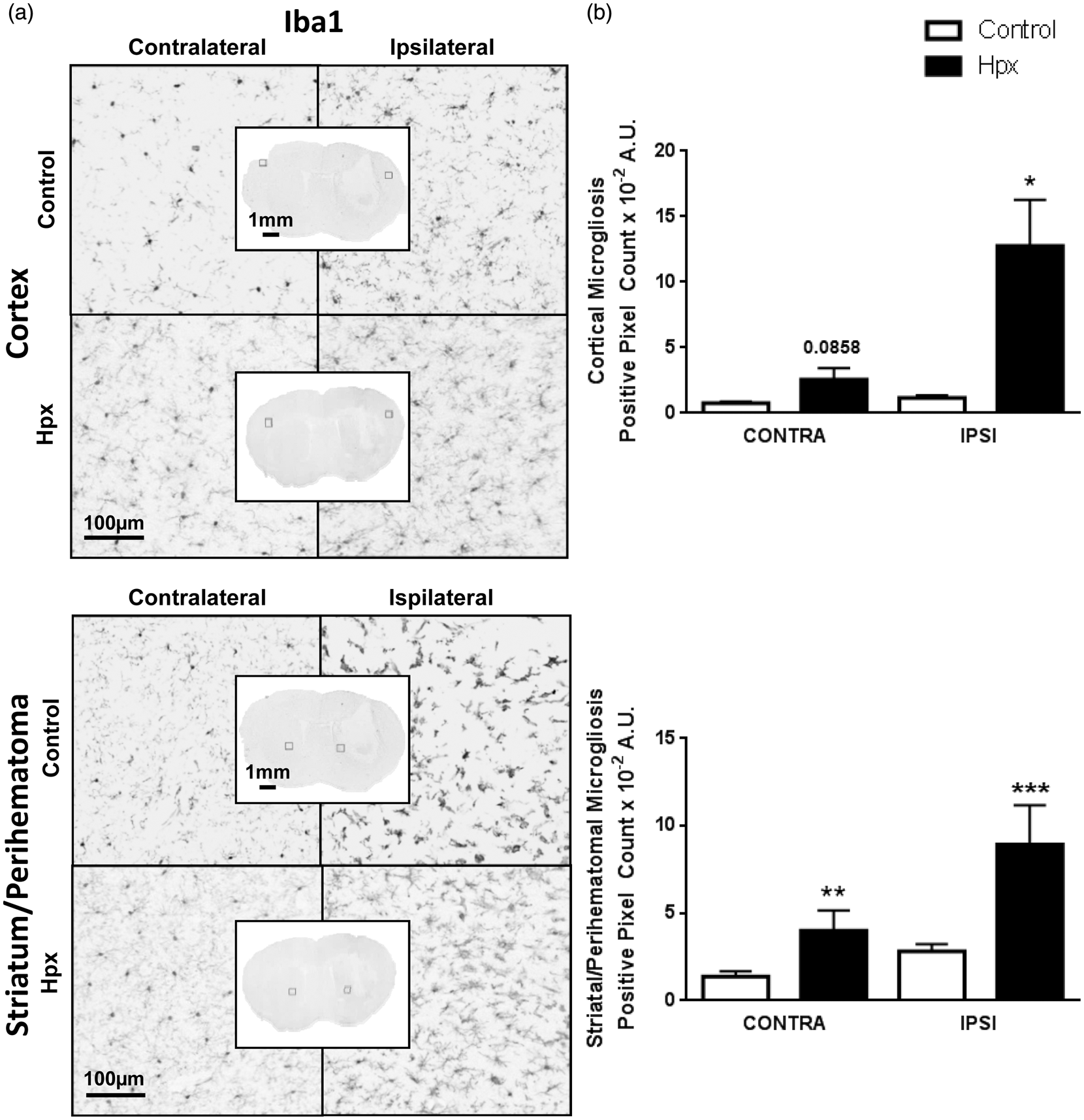
Discussion
This study is the first to evaluate the neuroprotective potential of high local Hpx levels after ICH. We reveal that rAAV1-mediated Hpx overexpression in the brain results in significantly smaller lesion volumes associated with less perihematomal tissue injury and trends toward reduced hematoma volume. This reduced anatomical damage is accompanied by significantly less body weight loss and reduced focal neurological deficits. Hpx mice have unchanged brain levels of iron and HO1, display significantly increased microgliosis, and have decreased astrogliosis and lipid peroxidation. Transgene-derived Hpx and/or Hpx-heme complexes are present in the CSF and serum after ICH, and peripheral heme-related markers are positively altered, possibly indicating that both central and peripheral heme clearance mechanisms are simultaneously operating. Collectively, these results suggest a neuroprotective role for Hpx after ICH and establish the possibility of administering clinical-grade Hpx locally as a therapeutic strategy for the treatment of ICH.
In agreement with previous studies demonstrating Hpx expression in the CNS,9–11,25,26 here we show that Hpx is present endogenously in the brain, albeit at low levels. These previous studies have shown Hpx expression from the vasculature, astrocytes adjacent to these vessels, ependymal cells, and neurons in the cortex, cerebellum, hippocampus, striatum, and CSF.9–11,25 A few studies have suggested that at least some degree of local Hpx upregulation occurs after an inflammatory insult or stroke.11,26,27 Hpx is reported to increase 4-fold in CSF following yeast-induced generalized acute inflammation, which was likely not a result of increased BBB permeability because the CSF total protein level was unchanged. 27 At 24 h after transient focal cerebral ischemia in rats, Hpx is upregulated approximately 2-fold in penumbral neurons and astrocytes in the cortex, hippocampus, and striatum. 11 Recently, it has been reported that Hpx levels also increase after ICH. 26 Although it is unclear what the relative contribution is from systemic infiltration of Hpx as part of the bleed and subsequent BBB dysfunction, and that of local Hpx upregulation, some insight can be gained by comparing the approximate 7-fold increase at 72 h relative to sham, and 3-fold increase at 72 h relative to 1 day post-ICH. 26 In summary, it appears that Hpx can be locally upregulated to some extent following general brain damage and after hemorrhage specifically, although, from a heme-binding stoichiometric perspective, this small upregulatation of Hpx levels would still be inadequate to handle the massive heme burden seen after ICH.
Here, we use rAAV1 vectors to specifically and constitutively increase brain Hpx levels, which results in very high in vivo expression, an estimated 60-fold relative increase from endogenous brain levels. Evaluation of the level of Hpx overexpression was performed by Western blotting of brain homogenates from non-ICH mice to avoid the confounding measurement of systemic-derived Hpx present after ICH. The local transgene-derived untagged Hpx runs at an identical molecular weight to endogenous liver and brain Hpx, implying similar processing and functionality. Furthermore, Western blotting of CSF from ICH mice demonstrates high levels of transgene-derived Hpx, suggesting it is secreted and soluble (anti-tag antibodies were used to specifically assess transgene-derived CSF Hpx and not endogenous CSF Hpx or Hpx present from potential blood contamination during CSF extraction, although the CSF used here was visibly clear). With in vitro transduction of mixed neuronal-glial cultures and Western blotting, we confirmed the secretory nature of our rAAV1-Hpx-(tag) constructs and that the protein products (Hpx, Hpx-V5, and Hpx-eGFP) run at the expected molecular weights. In vivo localization by immunohistochemistry showed that transgene expression is predominately neuronal mediated and highest surrounding the brain regions normally affected by an ICH. Thus, the highly overexpressed and secretory Hpx is perfectly positioned to bind and neutralize extracorpuscular heme released by hemolysis occurring within the hematoma, thereby potentially protecting surrounding viable brain tissue from secondary heme-mediated damage. Indeed, Hpx mice have significantly smaller lesion volumes with less perihematomal cell death and reduced neurological deficits, collectively demonstrating improved recovery after ICH.
Next, we investigated whether the transgene-derived Hpx expressed in the brain is able to clear to the periphery by ELISA and Western blotting of serum from mice that underwent ICH. For the latter approach, probing with an anti-tag antibody was used for similar reasons as described above for the CSF analyses. Intriguingly, we find a substantial and reproducible level of transgene-derived Hpx in the serum. Quantitative ELISA revealed a 62% increase in total serum Hpx levels for the Hpx mice. This transgene-derived Hpx seen in the periphery after ICH comes from the brain and could be the apoform of Hpx and/or represent Hpx-heme complexes because the methods used here do not distinguish between these forms. A couple possibilities exist to explain how brain-derived Hpx and/or Hpx-heme complexes appear in the serum: (1) receptor-mediated export and/or (2) bulk outflow, since ICH compromises the BBB. Regarding the former possibility, CD91/LRP1, the receptor for Hpx-heme complexes, among other ligands, does not recognize apo-Hpx. 28 Therefore, if receptor-mediated export were the operating mechanism, then the increased serum Hpx seen in Hpx mice would represent Hpx-heme complexes. In either case, a concentration gradient is likely to be present given the very high transgene-derived brain Hpx levels as compared to serum, driving transport/filtration of Hpx-heme complexes into the periphery. Furthermore, LRP1 is present on endothelial cells and has been shown to mediate transcytosis of its amyloid-ß ligand out of the brain, without a role for LRP1-mediated amyloid-ß degredation. 29 Additional studies will clarify the exact mechanisms leading to the results observed here. Nonetheless, the biocompartmental distribution specifically of the transgene-derived Hpx after ICH suggests that it is positioned to immediately bind and protect the CNS from heme toxicity and subsequently may mediate the safe degradation of heme by either central and/or peripheral mechanisms.
The balance between these two clearance pathways is of importance because too much internalization of Hpx-heme complexes centrally could lead to uncontrollably high intracellular heme and iron levels, increased oxidative stress, and persistent inflammation if other protective heme degradation and iron regulatory pathways are not concomitantly and locally induced to a safe level. Furthermore, if peripheral clearance mechanisms were also operating, it would allow some of the heme clearance to be taken care of in the large-capacity systemic system where scavenging pathways are not overloaded. Before commenting on these pathways and relevant work in the area, it should be noted that Hpx and Hpx-heme complexes are in general stable to the expected high in vivo concentrations of various peroxides present in the potent local oxidizing environment after ICH. 14 We and others have shown that incubation of high concentrations of Hpx-heme complexes with murine primary neurons 9 and human neuroblastoma cells 14 under ICH-like inflammatory conditions results in no apparent toxicity, whereas heme alone causes significant cell death. Furthermore, this neuroprotection is afforded via HO1-dependent pathways. 9 Hpx-heme complexes induce expression of the cytoprotective HO1 enzyme responsible for heme degradation (among other functions), although induction is somewhat impaired by exposure to peroxides, particularly high concentrations of H2O2.9,14 However, in another in vitro study using primary neuron/glial mixed cultures, equimolar extracellular Hpx decreased the accumulation and catabolism of exogenous hemin by neural cells and increased hemin export/extraction from membranes 4-fold, while HO1 expression was decreased, suggesting that Hpx-mediated neuroprotection is not arbitrated by increasing heme breakdown. 13 These seemingly conflicting findings on HO1 induction could be due to the different model systems and analytical methods used in the various studies.9,13,14
In the present in vivo study, Hpx mice had no change in HO1 expression or iron accumulation, but did have less lipid peroxidation. The latter finding suggests a direct protective effect of Hpx through its ability to bind heme and prevent its deleterious intercalation and peroxidation of membranes and harmful uncontrolled passage of heme into cells. Consistent with other in vivo studies, it is clear that HO1 is significantly induced locally surrounding the hematoma 8 where iron also deposits, directly supporting that some central heme degradation is occurring in this study, the extent of which is not significantly different between Hpx and control groups. Several possible interacting explanations exist to explain the absence of differences in HO1 expression and iron accumulation between groups: (1) neuroprotection after ICH with Hpx overexpression is primarily through a peripheral clearance mechanism, (2) high in vivo peroxide levels inhibit further HO1 induction with increased Hpx-heme internalization in Hpx mice, 14 (3) other related heme and iron modulatory proteins are induced, (4) total brain HO1 expression and iron accumulation are measured here (e.g. we are not differentiating for cell type-specific changes), and (5) various combinations of these possibilities. Additionally, the lack of observed differences in iron accumulation may be due to the 72-h study endpoint, when iron is starting to accumulate. Although this time point is the peak of HO1 expression, and thus it is an unlikely reason to explain the HO1 findings, and if HO1 is not different between groups at 72 h, then subsequent iron accumulation at later time points is less likely. Collectively, the aforementioned findings, in combination with the observed differences in peripheral serum levels of heme-related markers, indirectly support that both central and peripheral clearance mechanisms are concurrently operating.
Glial cell activation and neuroinflammation are intimately connected and important dynamic processes following ICH that can be both neurotoxic and neuroprotective.2,3 Here, we observe distinct inverse microglial and astrocyte activation and morphological changes. At this early time point, the increased microglial activation and trend toward reduced residual blood in Hpx mice may imply augmented phagocytic responses and improved hematoma resolution. Indeed, Hpx mice demonstrate a trend toward smaller hematoma volumes. Activated microglia are also reported to be less susceptible to heme toxicity. 30 Decreased astrogliosis may also be protective via a plethora of mechanisms. 31 It is also possible that these findings are in part independent of the ICH and a result of high local Hpx levels throughout late adolescent and adult life (it is unlikely to affect early developmental processes because rAAV1 vector expression is delayed post-P0 injection), something that is suggested by the contralateral differences noted here; however, brain-wide changes have been observed in various models of acute focal brain injury, including ICH.32–34 In either case, these glial cell activation changes are accompanied by significantly less ICH-induced brain injury in Hpx mice and thus likely represent a positive modulation of neuroinflammatory processes.
These novel findings provide evidence for the use of exogenous locally administered Hpx as a potential therapeutic for ICH. Additional studies are needed to evaluate the relative contribution of peripheral versus central clearance mechanisms that are likely concurrently operating to reduce the heme burden and improve outcomes following ICH.
Footnotes
Funding
This work was supported by grants from the McKnight Brain Research Foundation, Brain and Spinal Cord Injury Research Trust Fund (SD).
Acknowledgements
We thank all members of the Doré and Golde labs for helpful discussions and technical assistance, with a special acknowledgment of Carolina Ceballos-Diaz. The results of this study, in part, were previously presented at the following international meetings: SFN 2015 and ISC 2016.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors' contributions
JLL and SD conceived and designed the study. JLL performed surgical procedures, coordinated blinded behavioral testing, histology and quantification, analyzed data, and drafted the manuscript; JSM and AD performed blinded histology and quantification; ASL assisted with the quantification, data analysis, and figure preparation; JLL, PEC, and AMR cloned, prepared, and tested the rAAV1 vectors. JLL, TEG, and SD interpreted results and wrote the manuscript. All authors have approved the manuscript for publication.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.