
Abstract
Because our recent studies have demonstrated that miR-122 decreased in whole blood of patients and in whole blood of rats following ischemic stroke, we tested whether elevating blood miR-122 would improve stroke outcomes in rats. Young adult rats were subjected to a temporary middle cerebral artery occlusion (MCAO) or sham operation. A polyethylene glycol-liposome-based transfection system was used to administer a miR-122 mimic after MCAO. Neurological deficits, brain infarction, brain vessel integrity, adhesion molecule expression and expression of miR-122 target and indirect-target genes were examined in blood at 24 h after MCAO with or without miR-122 treatment. miR-122 decreased in blood after MCAO, whereas miR-122 mimic elevated miR-122 in blood 24 h after MCAO. Intravenous but not intracerebroventricular injection of miR-122 mimic decreased neurological deficits and brain infarction, attenuated ICAM-1 expression, and maintained vessel integrity after MCAO. The miR-122 mimic also down-regulated direct target genes (e.g. Vcam1, Nos2, Pla2g2a) and indirect target genes (e.g. Alox5, Itga2b, Timp3, Il1b, Il2, Mmp8) in blood after MCAO which are predicted to affect cell adhesion, diapedesis, leukocyte extravasation, eicosanoid and atherosclerosis signaling. The data show that elevating miR-122 improves stroke outcomes and we postulate this occurs via downregulating miR-122 target genes in blood leukocytes.
Introduction
Though a large number of compounds are effective in animal models of stroke, testing in human clinical stroke trials has not produced any treatments other than tissue plasminogen activator (tPA) for acute ischemic stroke. Such translational failures have been ascribed to various issues including animal models, experimental rigor, clinical trial designs and stroke heterogeneity. One potential contributor might be that drug development efforts for stroke have focused on small molecules that target a single gene, protein, or enzyme. Since ischemic tissue might die via many parallel pathways, blocking just one or two pathways may not be effective. A “cocktail approach” and agents with multiple mechanisms of action have been suggested, 1 but determining the optimum combination of therapies has been a challenge.2,3
These facts led us to consider miRNAs as a treatment for stroke in part because a single miRNA can modulate dozens to hundreds of target messenger RNAs (mRNA) by inducing mRNA degradation or blocking protein translation. 4 Since the initial discovery of miRNAs in the early 1990s,5,6 they have emerged as biomarkers for diagnosis and treatment of a large variety of diseases, including hepatitis, asthma, cancer and stroke.7–9 Moreover, there are now a number of reports showing that modulating miRNA (e.g. miR-497, Let 7f, miR-181, miR-15b, miR-30a, miR-23a-3p, miR-134, miR-200c, miR-424) can decrease brain infarction and attenuate behavioral deficits after cerebral ischemia in rodents.10–20 Moreover, increasing miR-133b in multipotent mesenchymal stromal cells (MSCs) promotes plasticity and functional recovery at 24 h after MCAO in rats. 21 Another promising feature of microRNA is that two miRNA drugs have already been advanced to human trials including a microRNA-122 (miR-122) inhibitor that treats hepatitis C infection 22 and a miR-34a mimic that treats cancer in humans. 23 Thus, miRNA-based therapeutics are feasible.
When we began to examine miRNA for the treatment of stroke, the question arose as to which of the >1000 miRNAs in the human genome might be the best candidates. To address this question, we proposed the following approach: miRNAs that are similarly regulated in blood of both rodents and in humans might be good targets to study as possible treatments for stroke. Our recent findings demonstrated that miR-122 was decreased in blood of ischemic stroke patients 9 and in the blood of ischemic rats. 8 In the current study, we confirm that miR-122 decreases in blood after MCAO in rats. This might make it more likely that results obtained in the rodent stroke models would apply to human stroke. Since the miR-122 sequence is conserved between vertebrate species, this also makes miR-122 a promising candidate for translation from rodent stroke models to human stroke.
In this study, we tested whether elevating blood miR-122 has the potential for treating stroke. Using a newly developed in vivo polyethylene glycol 2000 (PEG)-liposome based miRNA transfection system and the rat suture MCAO model, we show that intravenous (i.v.), but not intracerebroventricular (i.c.v.) injection of miR-122 mimic after MCAO dose-dependently improves stroke outcomes.
Materials and methods
Ischemic stroke model
Ischemic stroke was produced using the suture MCAO method.24,25Male Sprague-Dawley rats, at the age of 7–8 weeks, were anesthetized with isoflurane (Minrad, New York, NY, USA). A silicon-coated suture (Doccol Corporation, MA, USA) was introduced into the external carotid artery and advanced up the internal carotid artery to the Circle of Willis, where it blocks the origin of the MCA 24 to decrease blood flow of this artery for 1.5 h. Occlusions were confirmed by laser Doppler showing cerebral blood flow decreased to at least 20% of baseline during the 1.5 h occlusion. Measurements of physiological variables including temperature, pulse, blood pressure, respiratory rate, and blood gases showed no differences between the groups (not shown). Prior to the study, rats were blindly randomized to either sham surgery or MCAO surgery; and for each of these groups, they were further blindly randomized to receive either miR-122 mimic or scrambled sequence of miR-122 mimic as a control.
After closure of the operative sites, rats were allowed to recover in an incubator maintained at 37℃, and then returned to their home cages with free access to food and water. They were allowed to survive for 22.5 h after the 1.5-h MCAO. All experimental procedures were performed in accordance with National Institutes of Health guidelines and were approved by the Institutional Animal Care and Use Committee (IACUC) at the University of California at Davis. The manuscript was written up in compliance with the ARRIVE guidelines.
TaqMan miRNA assay of miR-122 expression in blood after MCAO
Male Sprague-Dawley rats (n = 24), at the age of 7–8 weeks, were divided into four groups (six rats/group). These included a sham operation group, and three groups of MCAO rats treated intravenously with scrambled miRNA mimic (2.4 mg/kg, i.v.) or miR-122 mimic (0.6 mg/kg and 2.4 mg/kg, i.v.) given within 5 min after the MCAO. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes and administered through the tail vein immediately after MCAO.
Whole blood was collected 24 h after MCAO in PAXgene tubes (Pre-AnalytiX) to lyse all leukocytes and other cells in blood and to also immediately stabilize RNA. Total RNA (including miRNA) was isolated from PAXgene tubes as previously described. 9 The concentration and integrity of the total RNA were determined using a NanoDrop-1000 spectrophotometer (Thermo Scientific, DE, USA) and Agilent-2100 Bioanalyzer (Agilent Technologies, CA, USA). RNA samples had a purity ratio of 28 s/18 s rRNA equal to or better than 1.8 and an A260/A280 absorbance ratio greater than 2.0. A TaqMan miR-122 assay (Life Technology, CA, USA) was employed to detect expression of miR-122 using a 7900HT real-time PCR instrument. The U6 miRNA was used as the endogenous control. Each group included three replicates (n = 6). Statistical differences were determined using ANOVA followed by Dunnett's post hoc test.
Animal groups and miRNA administration after MCAO
Male Sprague-Dawley rats (n = 30), at the age of 7–8 weeks, were divided into five groups (six rats/group). Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to administration (i.v. or i.c.v.) after MCAO. The groups included a sham operation group and three MCAO groups treated intravenously with scrambled miRNA mimic (2.4 mg/kg, i.v.) or miR-122 mimic (0.6 mg/kg and 2.4 mg/kg, i.v.) given within 5 min after MCAO. An additional group had the miR-122 mimic (0.6 mg/kg, i.c.v.) given intracerebroventricularly through a pre-drilled skull hole into the left lateral cerebral ventricles (coordinates: −0.9 mm caudal, −1.4 mm lateral, and −4.6 mm deep with respect to bregma) within 5 min after the MCAO while animals were still anesthetized. Behavioral evaluations, infarct measurements, and all analyses were performed by an investigator blinded to the treatment groups.
Assessment of neurological impairment
Neurological impairment was examined between 22 and 24 h after MCAO according to our previous methods. 24 Briefly, forelimb flexion, spontaneous rotation, and absence of response to contralateral whisker stimulation were scored on a 0–2 scale (0 – normal behavior; 1 – moderate impairment; and 2 – severely impaired). Statistical tests used the non-parametric Kruskal–Wallis test. Testing was performed by an investigator blinded to the treatment status.
Cresyl Violet staining and measurement of brain infarction
At 24-h post MCAO animals were anesthetized and euthanized. Brains were removed and processed for Cresyl Violet staining. 26 Brain sections were scanned, and infarcted and healthy tissues were measured using an image analysis system (Adobe Photoshop). 24 Statistical tests used an ANOVA followed by Dunnett's post hoc test. Measurements were performed by an investigator blinded to the treatment status.
Immunohistochemistry-double labeling for ICAM-1 and vWF
Formaldehyde-fixed brain sections were incubated with primary antibodies to mouse anti-ICAM-1 (1:200, Abcam, MA, USA) and rabbit anti-vonWillebrandt Factor (vWF) (1:200, Abcam, MA, USA). Species-specific IgG, conjugated to Alexa 594 or 488 (1:1,000; Life Technology, CA, USA), were used as secondary antibodies. Controls had the primary antibody deleted.
TaqMan qRT-PCR array to profile the expression of miR-122 target genes in blood after MCAO with or without the miR-122 treatment
Blood was collected via cardiac puncture 24 hours after MCAO, and total RNA was isolated and amount and purity quantified according to our previous protocols. 9 TaqMan custom mRNA 384-well fluidic cards (Supplemental Table 1 showing the plate layout, Life Technology, CA, USA) were used to profile gene expression in blood. A list of the genes tested by qRT-PCR is provided in Supplemental Table 1. This list was compiled based upon known miR-122 target genes, genes known to be regulated by miR-122 target genes (indirect target genes) and genes implicated in stroke but not known to be miR-122 direct or indirect target genes. The cards were run using a 7900HT real-time PCR instrument. 18S was used as the endogenous control. The statistical analyses were performed using Partek Genomics Suite 6.4 (Partek, MI, USA), and differentially expressed mRNAs in the group of MCAO/ miR-122 mimic were determined by student's t-test, as compared to the group of MCAO/scrambled miRNA mimic. A p < 0.05 and fold change > |1.4| were considered significant.
Custom database of miR-122 target genes
To predict miR-122 target genes, we used two separate publicly available miRNA-mRNA function algorithms, TargetScan and miRanda. This generated a custom database of 1120 predicted miR-122 target genes, including many immune response- or stroke outcome-related genes (e.g. Ccl22, Vcam1, Fkbp5, Il1rn, Il22ra2, C1qb, Nos2, Clic4, Epo, Aldoa, Calpain 1, and others) (Supplemental Table 2). A subgroup of these known miR-122 target genes was measured by qRT-PCR as described in the previous section.
Integrated miR-122, altered mRNA network and functional analyses
The integrated functional network analyses for miR-122 and altered mRNAs were performed according to our methods described previously with minor modifications. 27 Briefly, the data list of significantly down-regulated mRNAs, and the custom database of miR-122 and its putative target genes were uploaded into the integrated miRNA-mRNA functional network analyses software: Exploratory Gene Association Networks (EGAN). We expected that the genes down-regulated by the miR-122 mimic after MCAO could be divided into three groups: (1) known miR-122 direct target genes – that were defined as those with a mRNA sequence complementary to the miR-122 sequence; (2) miR-122 indirect target genes – that were defined as those known to be regulated by miR-122 target genes; and (3) novel miR-122 indirect target genes – that were defined as those genes regulated following miR-122 administration but had no sequence homology to miR-122 and were not previously known to be regulated by miR-122 direct or indirect target mRNA. After the integrated miR-122 and target genes networks were identified, a functional analysis was performed on miR-122 direct and indirect genes using Ingenuity Pathway Analysis (IPA).
Results
Effects of miR-122 mimic on miR-122 expression in blood after MCAO
The TaqMan miRNA assay showed that miR-122 decreased in blood after MCAO (Figure 1). That is, the miR-122 blood levels in sham-operated control animals were several fold more than in animals following MCAO (treated with control scramble miRNA, left panels of Figure 1). Intravenous administration of miR-122 mimic (0.6 mg/kg and 2.4 mg/kg, i.v.) dose-dependently elevated blood miR-122 after MCAO compared to MCAO treated with control scramble miRNA (*p < 0.05 for 2.4 mg/kg i.v miR-122 mimic vs. MCAO/scramble miRNA) (Figure 1). Though blood levels of miR-122 increased nearly four-fold following 0.6 mg/kg i.v. of miR-122 mimic compared to scramble miRNA, this increase was not statistically significant (Figure 1).
miR-122 levels decreased in blood 24 h following MCAO (Sham vs Scramble miRNA-MCAO). miR-122 mimic given intravenously (0.6 mg/kg and 2.4 mg/kg i.v) elevates miR-122 levels in blood 24 h after MCAO in rats. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to intravenous administration after MCAO. miR-122 levels in blood are shown on the Y axis. #p < 0.05 for scramble miRNA-MCAO vs. sham. *p < 0.05 for miR-122 mimic-MCAO (2.4 mg/kg) vs. scramble miRNA-MCAO. n=6/group.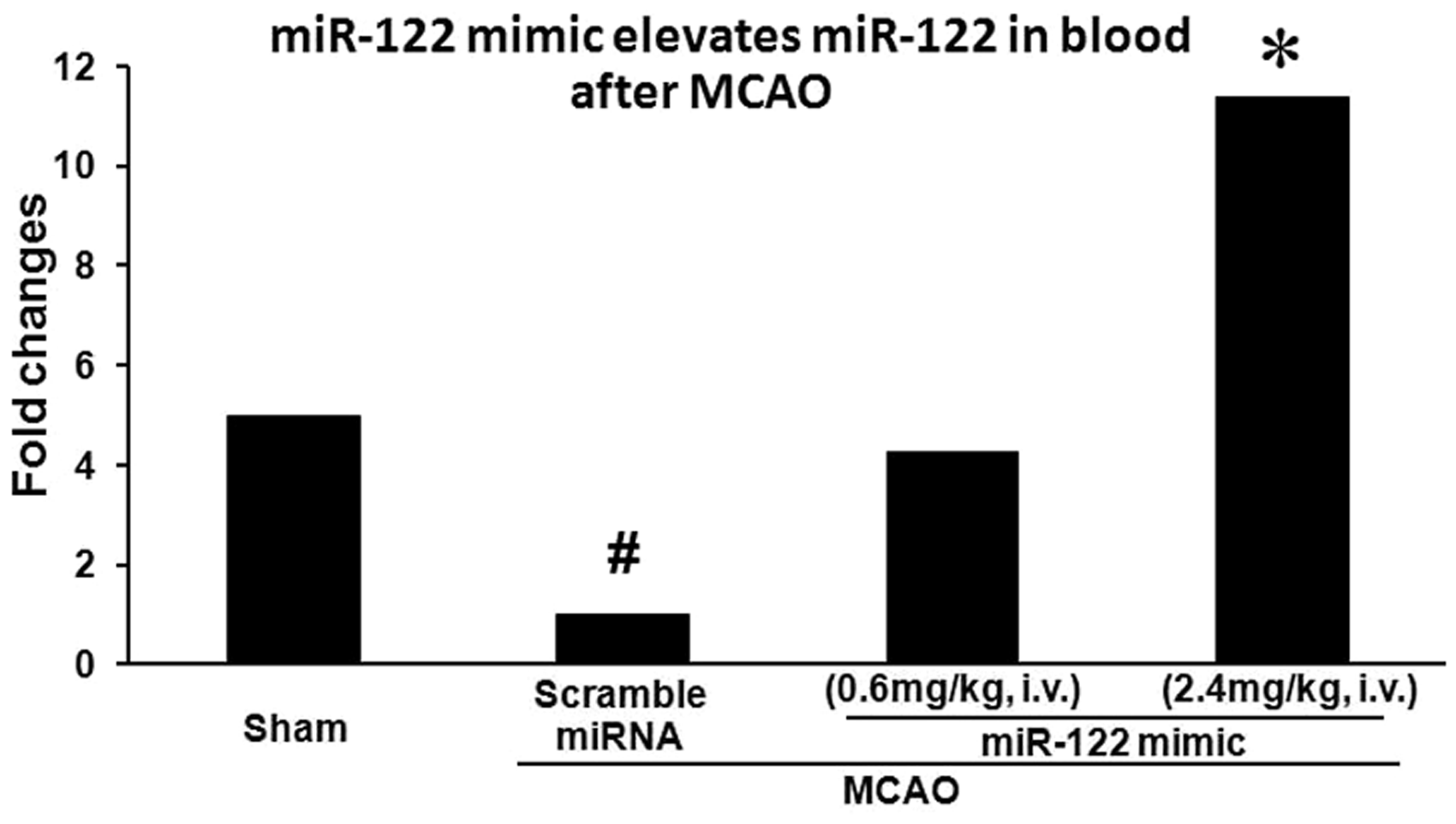
The effects of miR-122 mimic on behavioral outcomes after MCAO
The behavioral results showed that the scramble miRNA (control)-treated animals developed marked behavioral deficits after MCAO (#p < 0.05 vs. sham), while miR-122 mimic given i.v. (0.6, 2.4 mg/kg, i.v.) dose-dependently attenuated the MCAO-induced behavioral deficits (Figure 2, *p < 0.05 for 2.4 mg/kg i.v. miR-122 mimic vs. MCAO/ scramble miRNA). However, miR-122 mimic given intracerebroventricularly (0.6 mg/kg, i.c.v.) did not improve behavioral outcomes (Figure 2).
miR-122 mimic given intravenously (0.6 mg/kg and 2.4 mg/kg i.v.) attenuates behavioral deficits after MCAO in rats, but miR-122 mimic given intracerebroventricularly (0.6mg/kg i.c.v.) does not. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to i.v. or i.c.v. administration after MCAO. Neurological scores are shown on the Y axis. #p < 0.05 for scramble miRNA-MCAO vs. sham; *p < 0.05 for miR-122 mimic-MCAO vs. scramble miRNA-MCAO. n = 6/group.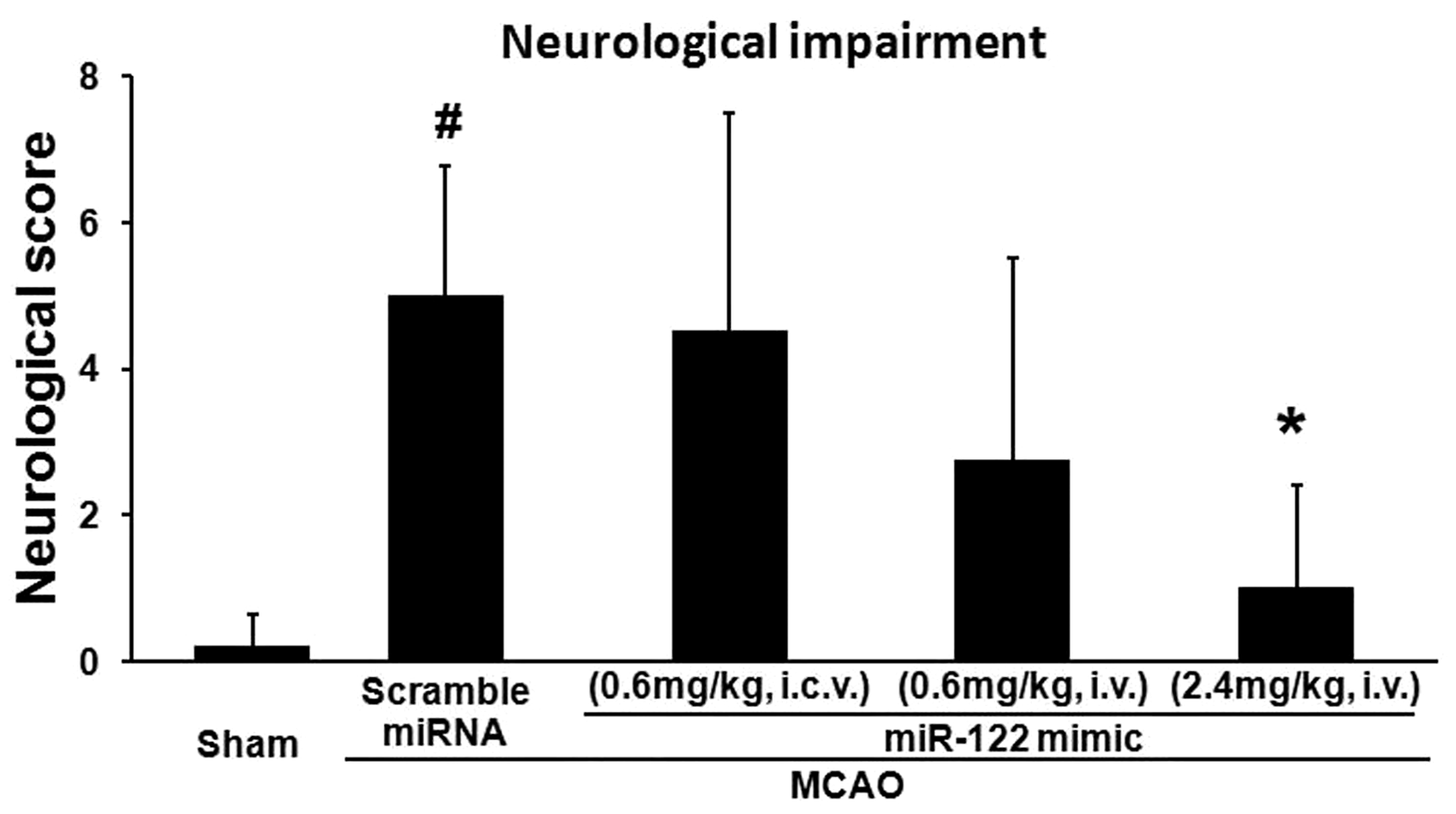
The effects of miR-122 mimic on brain infarction volumes after MCAO
Cresyl Violet histology showed that miR-122 mimic at doses of 0.6 mg/kg and 2.4 mg/kg dose-dependently attenuated the MCAO-induced brain infarction volume (Figure 3(a) and (b), *p < 0.05, **p < 0.01 for miR-122 mimic i.v. vs. MCAO/ scramble miRNA i.v.). However, intracerebroventricular miR-122 mimic at a dose of 0.6 mg/kg, i.c.v. did not decrease infarction volume (Figure 3(a) and (b)).
miR-122 mimic given intravenously (i.v.) but not intracerebroventricularly (i.c.v.) decreases brain infarction after MCAO in rats. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to i.v. or i.c.v. administration after MCAO. (a) Cresyl Violet staining of coronal brain sections from five different rats. Column 1 sections are from a sham control. Column 2 sections are from scramble miRNA-MCAO rats. Column 3 sections are from miR-122 mimic MCAO (0.6mg/kg i.c.v.). Column 4 sections are from miR-122 mimic MCAO (0.6mg/kg i.v.). Column 5 sections are from miR-122 mimic MCAO (2.4mg/kg i.v.). (b) Following MCAO, miR-122 mimic (0.6 mg/kg i.v.) decreased infarction volume compared to scramble miRNA (*p < 0.05) and miR-122 (2.4 mg/kg i.v.) also decreased infarction volume compared to scramble miRNA (**p < 0.01). There was no significant difference of infarction volume for miR-122 mimic (0.6 mg/kg i.c.v) compared to scramble miRNA 24 hours following MCAO. Infarction volumes are on the Y-axis. *p < 0.05, **p < 0.01 for miR-122 mimic-MCAO vs. scramble miRNA-MCAO. n = 6/group.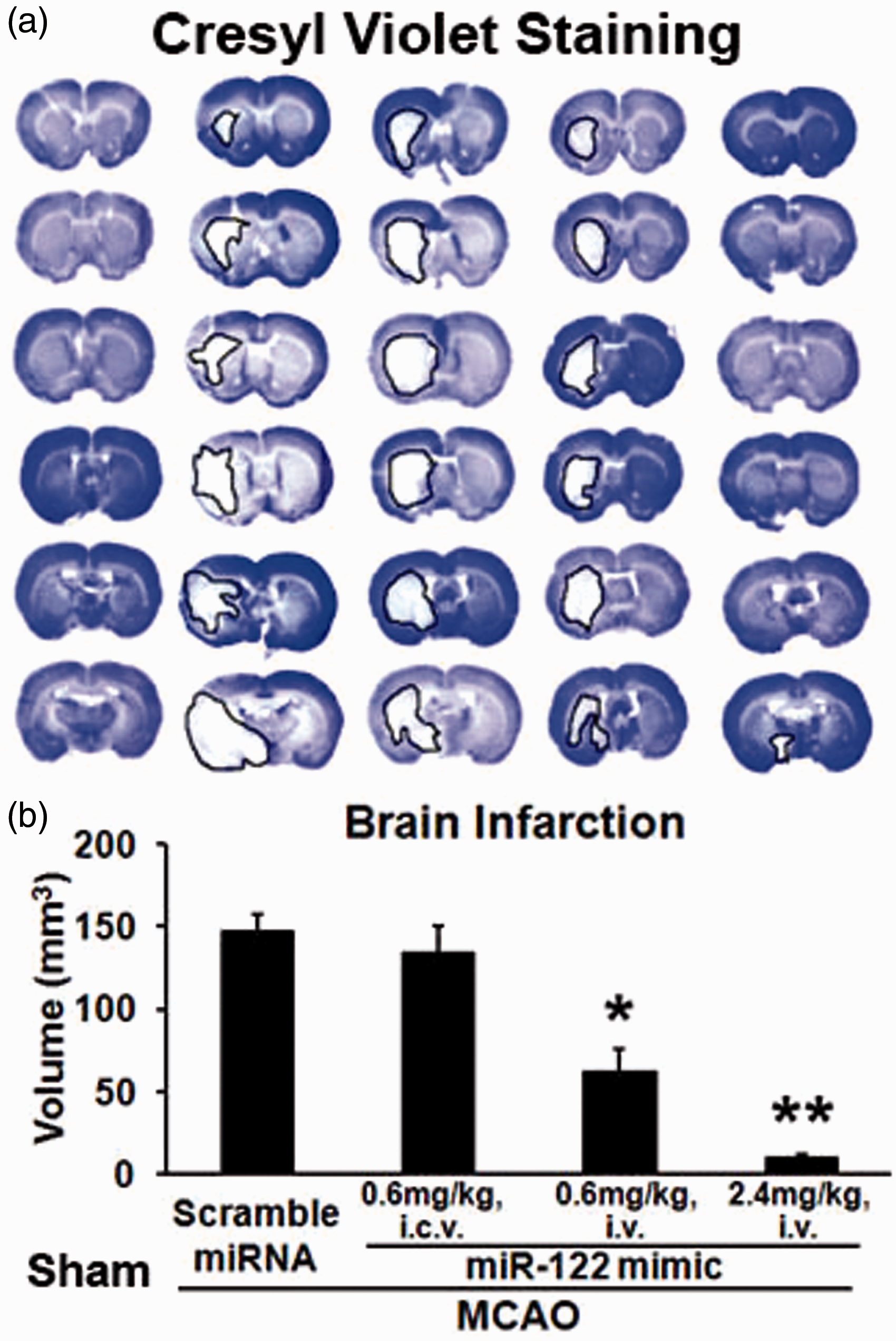
The effects of miR-122 mimic on blood brain barrier integrity and adhesion molecule expression after MCAO
In order to assess inflammation and integrity of brain vessels, brain sections were double labeled with intercellular adhesion molecule 1 (ICAM-1, a marker for immune cells and injured endothelial cells) and von Willebrand factor (vWF, a marker for endothelial cells). In sham-operated rats, the cerebral vessels had an intact tube shape, and no ICAM-1 was expressed in brain parenchyma and vessels (Figure 4(A) to (C)). In scramble miRNA-treated MCAO animals, ICAM-1 was observed in brain parenchyma (Figure 4(E) and (F)), and there was attenuation of the caliber of the vessels in areas of infarction (Figure 4D). MiR-122 mimic (2.4 mg/kg, i.v.), given immediately after MCAO, mostly prevented the MCAO-induced ICAM-1 expression in brain parenchyma (Figure 4(H) and (I)) and maintained the caliber and tube shape of the vessels (Figure 4(G) and (I)).
miR-122 (2.4mg/kg i.v.) mimic maintains vessel caliber, vWF immunoreactivity, and prevents ICAM-1 induction one day after MCAO in rats compared to scramble miRNA. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to i.v. administration after MCAO. Note induction of ICAM1 with scramble miRNA (E) which is blocked by miR-122 mimic (H). Note decrease in vessel caliber following scramble miRNA (D) which is prevented by miR-122 mimic (G). n = 4/group. Scale bars: A–I, 50 µm.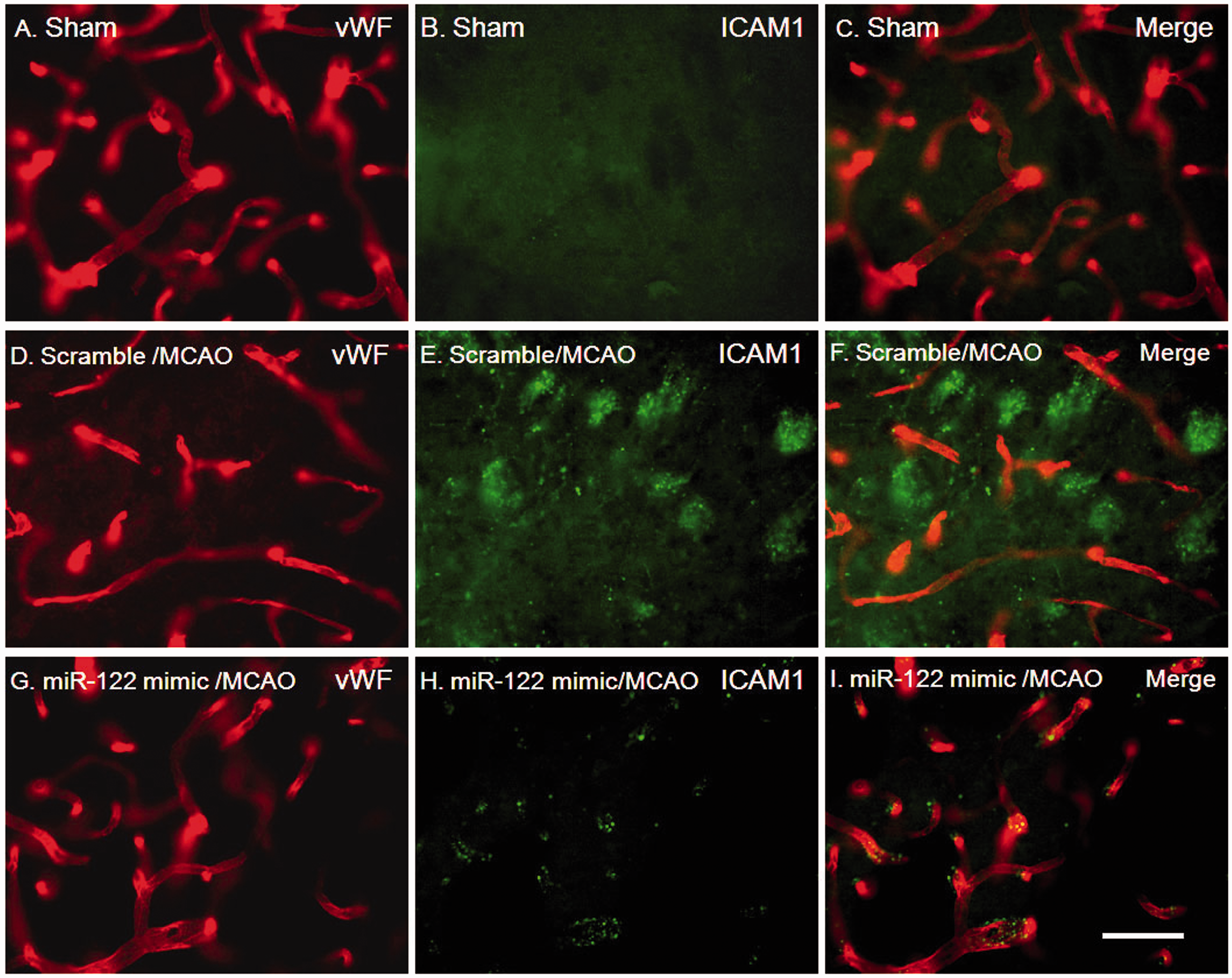
Predicted functions of genes down-regulated by miR-122 mimic in blood after MCAO
TaqMan PCR arrays were used to detect gene expression after MCAO with or without miR-122 mimic treatment. The data showed that miR-122 mimic (2.4mg/kg, i.v.) down-regulated a number of genes in blood after MCAO (p < 0.05 and fold change > |1.4|) (Supplemental Table 3). In these functional assays, we applied a relaxed cutoff (fold change > |1.4| but p > 0.05) to include more down-regulated genes in blood after MCAO (Supplemental Table 3). Using the Exploratory Gene Association Networks (EGAN), we classified the down-regulated genes into three sub-groups as described in the Methods section: (1) miR-122 direct target genes that included Vcam1, Nos2, Pla2g2a, Ucp2, Clic4, and others (upper left panel in Figure 5); (2) miR-122 indirect target genes that included Alox5, Itga2b, Timp3, IL1b, IL2, MMP8, and others (upper right panel in Figure 5); and (3) miR-122 novel indirect target genes that included Ltb4r2, Ifna1, Ilrapl2, Mmp11, Col4a2, Myl9, Mylk, F2rl2, Thbs1 and others (lowest panel in Figure 5).
miR-122 mimic down-regulates genes in blood after MCAO in rats. Scrambled miRNA mimic and miR-122 mimic were wrapped using PEG-liposomes prior to intravenous administration after MCAO. Upper left panel: miR-122 direct target genes down regulated 24 h following MCAO which have sequences complementary to miR-122. Upper right panel: miR-122 indirect target genes down-regulated 24 h following MCAO. The indirect target genes shown are known to be regulated by Vcam1 and/or Nos2- which are direct miR-122 target genes. Middle panel: top 5 ranked canonical pathways of the miR-122 direct and indirect target genes that were down-regulated by miR-122 mimic 24h following MCAO. Lower panel: these are mRNAs that were down regulated 24 h following miR-122 treatment. These genes are not known to be either direct or indirect miR-122 targets. Thus, these represent novel miR-122 indirect target genes that must be regulated by one of the known miR-122 direct target genes. There are two colors of the lined circles in upper and lower panels: (1) the red lined circled genes were deceased greater than three times; and (2) the black lined circled genes were decreased ranging from 1.4 through three times. In addition, there are three thicknesses of the lined circles in upper and lower panels: (1) the small thickness lined circled genes were decreased with p > 0.05; (2) the medium thickness lined circled genes were decreased with 0.01 < p < 0.05; and (3) the large thickness lined circled genes were decreased with p < 0.01. (n = 6/group).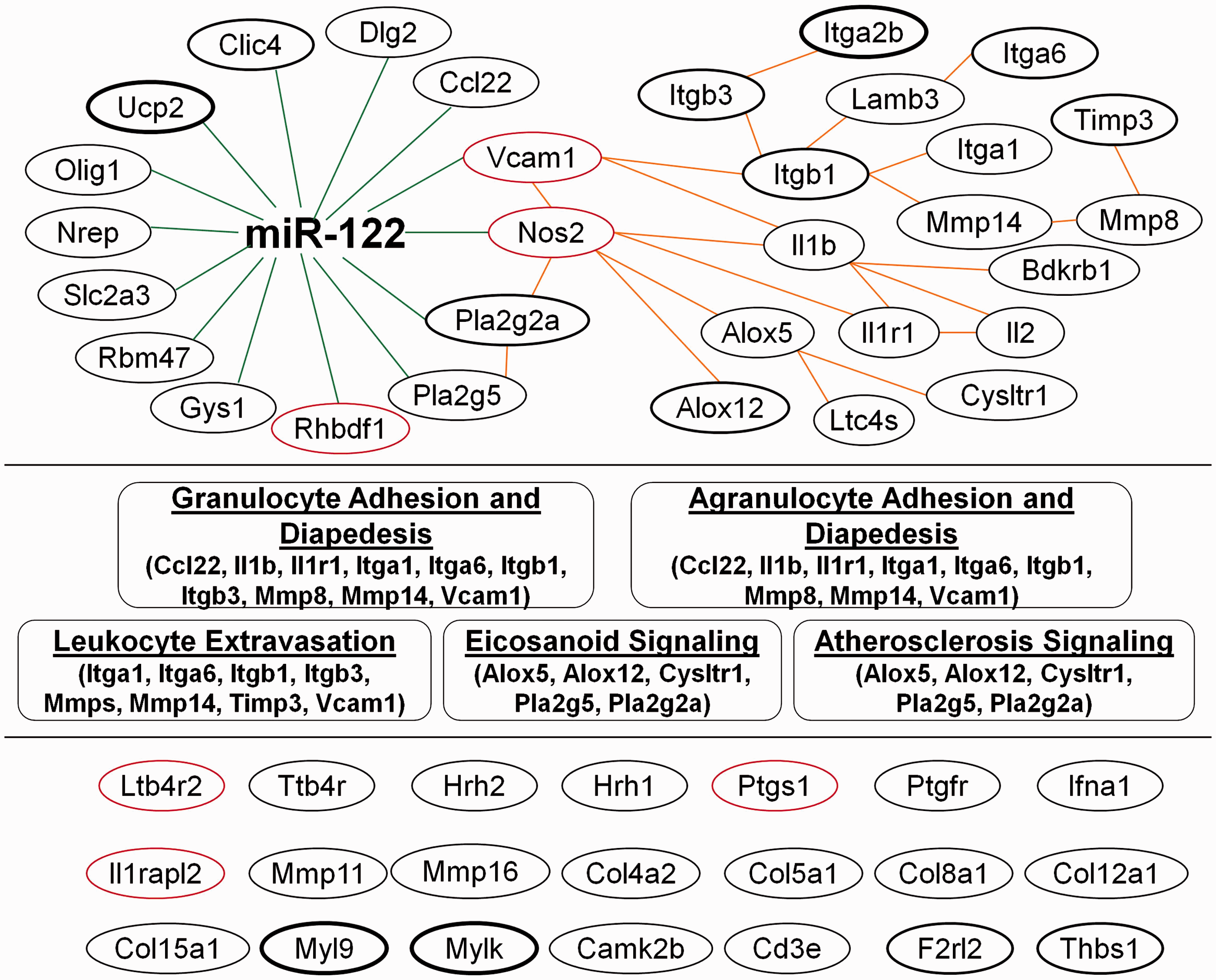
Using Ingenuity pathway analysis (IPA), we analyzed the functions of miR-122 direct and indirect target genes. The top 5 ranked canonical pathways included granulocyte/agranulocyte adhesion and diapedesis, leukocyte extravasation, eicosanoid signaling and atherosclerosis signaling (middle panel in Figure 5).
Discussion
This is the first study to show that administration of a miR-122 mimic improves ischemic stroke outcomes. It adds to a growing literature that many miRNA can modulate ischemic brain injury at least in rodent models.10–21 This proposal is unique in that it targeted miR-122 because miR-122 decreased in whole blood following both rat and human strokes,8,9 and miR-122 improved ischemic stroke outcomes when injected into blood but not when injected into brain.
In our previous human study, we showed that miR-122 and many other miRNA decreased in whole blood following ischemic stroke using both microarray and PCR based methods. 9 The miRNA described in that study likely represent mostly intracellular miRNA found in leukocytes, platelets, red blood cells, and other cells in whole blood since intracellular miRNA levels are usually an order of magnitude or more higher than extracellular miRNA levels in plasma or serum. 9 However, miR-122 has been detected in plasma/serum and reported to be altered in liver infections and liver cancer. 28 Though miR-122 was once thought to be liver-specific, elevated levels of plasma miR-122 have also been reported in coronary artery stenosis and myocardial infarction 29 and cardiogenic shock. 30 The current study confirmed our previous studies that ischemic stroke decreased miR-122 in whole blood of rats, 8 with similar findings in whole blood of humans after stroke. 9 These findings prompted us to test whether administration of miR-122 would affect ischemic stroke outcomes.
microRNA mimics and PEG-lysosome
To administer miR-122, we used the commonly used mimic approach. The miRNA mimics are chemically modified, double-stranded RNAs that are intended to “mimic” native miRNA which are incorporated into the RNA-induced silencing complex (RISC), which then recognizes target mRNAs through base-pairing, and decreases the target mRNA expression by post-transcriptional gene silencing. 31 The miRNA mimics are more stable and produce longer lasting effects in blood and tissues than native miRNA. To achieve effective intracellular delivery of the miRNA mimic in vivo, we utilized the polyethylene glycol 2000 (PEG)-liposome-based miRNA in vivo transfection method. There is little detectable toxicity or inflammatory response due to the biodegradable PEG liposome. The PEG-liposome-conjugated complexes are stable in serum for at least 16 h. This improves the in vivo miRNA mimic half-life from minutes to many hours and enhances miRNA mimic entry into cells. 32 The PEG-liposome-based transfection system produces a transient elevation of miR-122 after administration, thus avoiding long lasting or developmental side effects of transgenic over expression.
miR-122 may improve outcome by acting on leukocytes – not brain
The miR-122 mimic/ PEG-liposome complexes given intravenously dose dependently elevated miR-122 one day following MCAO. Though miR-122 mimic at the lower dose (0.6 mg/kg, i.v.) increased the level of miR-122 in blood after MCAO, these changes were not statistically significant (Figures 1 and 2). However, the low dose of the (0.6 mg/kg, i.v.) miR-122 mimic did significantly decrease brain infarction volume after MCAO (Figure 3). This suggested that the 0.6 mg/kg (i.v.) dose might be right at the threshold for improving outcomes after MCAO. Indeed, miR-122 mimic at a four-fold higher dose (2.4 mg/kg, i.v.) consistently and statistically significantly increased the level of miR-122 in blood, improved neurological impairment, and decreased brain infarction volume after MCAO.
Compared to scramble miRNA-treated MCAO animals, miR-122 mimic at the lower dose (0.6 mg/kg, i.v.) mostly protected cortex (penumbra) without having much effect on striatal infarction (core) (Figure 3). Since the striatal damage mostly contributes to neurological scores measured in this study, and the striatal damage was similar in the scramble and miR-122 (0.6 mg/kg, i.v.)-treated animals (Figure 3), this may help explain why the low dose miR-122 mimic (0.6 mg/kg, i.v.) significantly decreased brain infarction volume after MCAO (Figure 3) while tending to decrease neurological impairment, but this did not reach significance (Figure 2). Lastly, even though the 0.6 mg/kg iv dose of miR-122 mimic did increase miR-122 blood levels nearly four fold (Figure 1), this was not statistically significant because of the variability.
Though the low dose of the miR-122 mimic (0.6 mg/kg, i.c.v.) decreased infarct volume, the same dose injected into the brain ventricles (0.6 mg/kg, i.c.v.) did not decrease infarction volume. Since the intravenous dose is distributed in 250–300 g of body tissue, while the ventricular dose is distributed in 2 g of brain, concentrations nearly 100 times less in blood improved stroke outcome whereas concentrations over 100 times greater in brain did not affect stroke outcome. Therefore, we postulate that intravenous administration of miR-122 mimic likely acted on peripheral blood leukocytes or perhaps other blood cells to improve stroke outcomes. miR-122 expression has previously been reported in peripheral blood mononuclear cells. 33 Also, miR-122 has anti-inflammatory actions in the liver 34 and modulates immune genes in zebrafish following LPS stimulation. 35
It is also possible intravenous miR-122 acted on brain blood vessels. However, if this were the case, then one has to postulate that intraventricular miR-122 mimic did not improve stroke outcomes because it acted on abluminal sides of vessels, whereas intravenous miR-122 mimic acted on luminal sides of vessels directly on endothelial cells to improve stroke outcomes. If miR-122 acted directly on endothelium, this might help explain why miR-122 decreased ICAM-1 expression and maintained vessel integrity. Future studies will be needed to address these questions.
To examine possible mechanisms by which miR-122 might protect against ischemic stroke, we measured levels of selected miR-122 target mRNAs. In modeling miRNA-mRNA interactions, we used the classical/most commonly encountered model in which miRNAs bind to complementary sequences in the three prime untranslated regions (3′UTRs) of target mRNAs, resulting in degradation of the target mRNA or suppression of translation of the target mRNA.4,36,37 The data show that the miR-122 mimic down-regulated a number of mRNA in blood after MCAO. Using EGAN and IPA, we found that miR-122 down-regulated direct and indirect target genes associated with cell adhesion, diapedesis, leukocyte extravasation, elcosanoid, and atherosclerosis signaling. These pathways suggest that the miR-122 mimic might modulate cell infiltration after ischemic stroke, something which can be assessed in future studies. In addtion, miR-122 acts on other target genes and pathways that have been associated with stroke outcomes including Nos2 (neuronal Nos), Chloride intracellular channel 4, Erythropoietin, Aldolase A and calpain 1. It is important to emphasize that all of the possible miR-122 target mRNA were not examined here, and which include regulation of iron metabolism, 38 mitochondria, 39 and cholesterol metabolism. 40
There are several limitations of the current study. Future studies will need to examine miR-122 expression after i.c.v. administration of miR-122 mimic in MCAO animals, determine how i.c.v. administration affects miR-122 target gene expression in brain, and to determine if i.c.v. miR-122 mimic at higher doses could prevent brain damage after MCAO, as it was possible that miR-122 mimic at the dose (0.6 mg/kg, i.c.v.) used in this study was not high enough to protect brain cells after MCAO. Future studies will also need to directly test whether miR-122 mimic modulates gene expression of leukocytes and/or cerebral endothelial cells. Future studies will also need to address STAIR criteria for testing stroke therapeutics including testing both sexes, testing aged animals, defining the therapeutic time window, testing in another species and testing in animals with stroke risk factors. One more limitation is that miR-122 may or may not directly bind to the target genes predicted by EGAN. Therefore, future studies will need to be performed to evaluate miR-122 binding sites on its target genes using the luciferase reporter assay system.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article:
This study was supported by the University of California at Davis Innovative Development Award (DZL), NIH grant R01NS089901 (DZL), American Heart Association Fellow to Faculty Award (GCJ) and NIH grant R01NS066845 (FRS).
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Authors' contributions
DZL, GCJ, BPA, HH, XZ, CC, NS, CD-A, BS and FRS participated in the conduct and analysis of the studies and have read and reviewed the manuscript. DZL and FRS designed the experiments and wrote the manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.