
Abstract
In inhalation toxicity studies, drug-induced lesions are frequently reported in the larynx and sometimes at the tracheal bifurcation (carina) in the rat, but less so in the dog or monkey, bringing into question the relevance of these rodent findings for humans. The rat larynx is widely considered to be more sensitive than that of the dog and monkey in its response to inhaled xenobiotics, although we could find no published data to support this. In this review, data from 52 inhalation studies involving rodent and nonrodent species were collated and reviewed. These data showed that the rodent larynx, and to a lesser extent the carina, was far more commonly affected by treatment than those of the nonrodent. This review indicates the greater susceptibility of the rodent larynx and carina and emphasizes their lack of relevance for man. Observations and data suggest that the human larynx is much closer to the beagle dog and cynomolgus monkey in its response to inhaled xenobiotics and that greater clinical relevance should be placed on any specific findings in these animal models.
Inhalation toxicity studies using rodent and nonrodent species are usually required by regulatory agencies as part of the development and nonclinical safety testing of novel inhaled pharmaceuticals. Two key anatomical sites assessed during these studies are the larynx and tracheal bifurcation (carina). It is well accepted that the rodent larynx is particularly sensitive to aerosol damage (Osimitz, Droege, and Finch 2007; Gopinath, Prentice, and Lewis 1987), although no literature appears to exist that compares the response of the rodent larynx with that of the dog and primate.
Much published information is available describing laryngeal injuries in rodents induced by a wide range of compounds through inhalation exposure (Osimitz, Droege, and Finch 2007; Gopinath, Prentice, and Lewis 1987; Renne et al. 2007; Lewis 1991), but few reports exist for dogs and primates. Furthermore, very little published literature is available describing treatment-related lesions at the tracheal bifurcation in any species.
In this review, laryngeal and carinal findings were tabulated from a series of inhalation studies to compare the sensitivity of rodent and nonrodent tissues. In addition, the significance for humans of commonly observed pathological lesions at these target sites is discussed.
Materials and Methods
Data from 52 inhalation studies were included in the survey (Table 1). These studies were organized as 26 cases, with each case comprising a pair of studies where the same test material had been administered to a rodent and a nonrodent species. In each case, comparable doses were given to both species with the same length of exposure (typically 1 hr/day) and study duration (1 day to 91 days). Administration was by face mask to nonrodents, and nose (snout)-only tubes were used for rats and mice.
Number and Duration of Selected Studies.
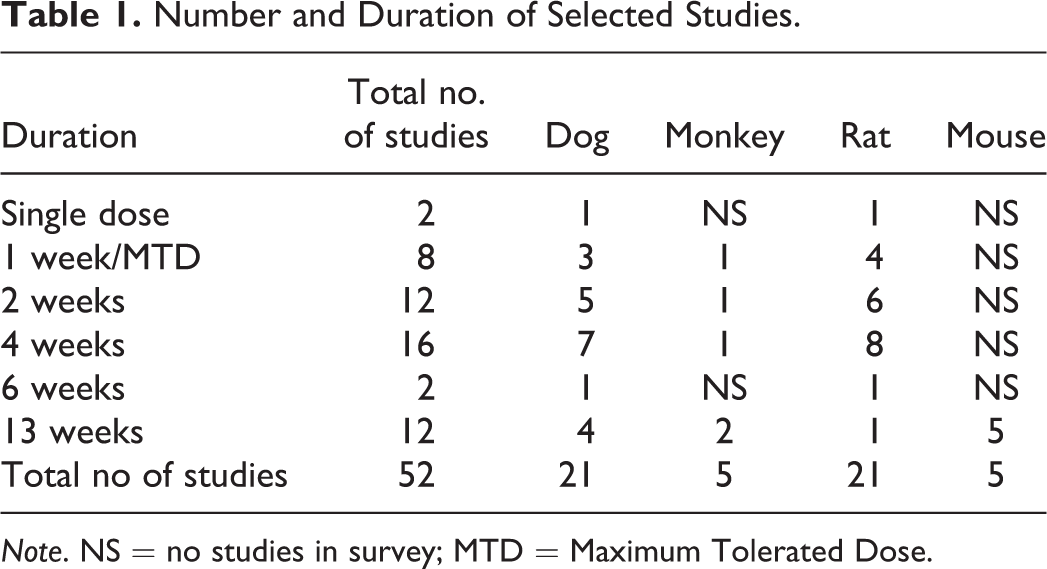
Note. NS = no studies in survey; MTD = Maximum Tolerated Dose.
For inclusion in the survey, cases were selected when at least 1 species displayed test article–related changes in the larynx and/or carina. Study pairs where neither species showed test item–related lesions were not included. The selected studies covered 22 compounds comprising vehicles or pharmaceuticals of various classes, including muscarinics, adrenergics, steroids, antibiotics, and biologics. All compounds were administered as liquid or dry powder aerosols. Particulate mean aerodynamic diameter for all studies was within acceptable levels for inhalation administration (1–4 µm) and was similar or identical between the species tested.
Animal Management
All studies and procedures were conducted in accordance with study plans, the principles of Good Laboratory Practice and UK Home Office Legislation (Animals [Scientific Procedures] 1986 Act). The following strains of laboratory species were used in this review: rats: Sprague-Dawley or Han Wistar; mice: CD-1; dogs: Harlan or Marshall beagles; and primates: cynomolgus monkeys sourced from Philippines or Vietnam.
Necropsy and Histology Procedures
Necropsies were conducted according to standard operating procedures. The tissues were fixed in 10% neutral buffered formalin. The tracheal bifurcation from all species was trimmed in accordance with published guidelines (Kittel et al. 2004). For rodents, 5 transverse sections of larynx were prepared, based on recommended guidelines (Kittel et al. 2004), with 2 additional sections to ensure coverage of critical areas. Two transverse sections of larynx were prepared from dog and monkey, 1 at the base of the epiglottis and 1 at the level of the vocal cords. Tissues were conventionally processed, embedded in paraffin wax, sectioned at 4 µm to 6 µm, and stained with hematoxylin and eosin. All light microscopic evaluations were subjected to peer review by a second pathologist.
Results
Larynx
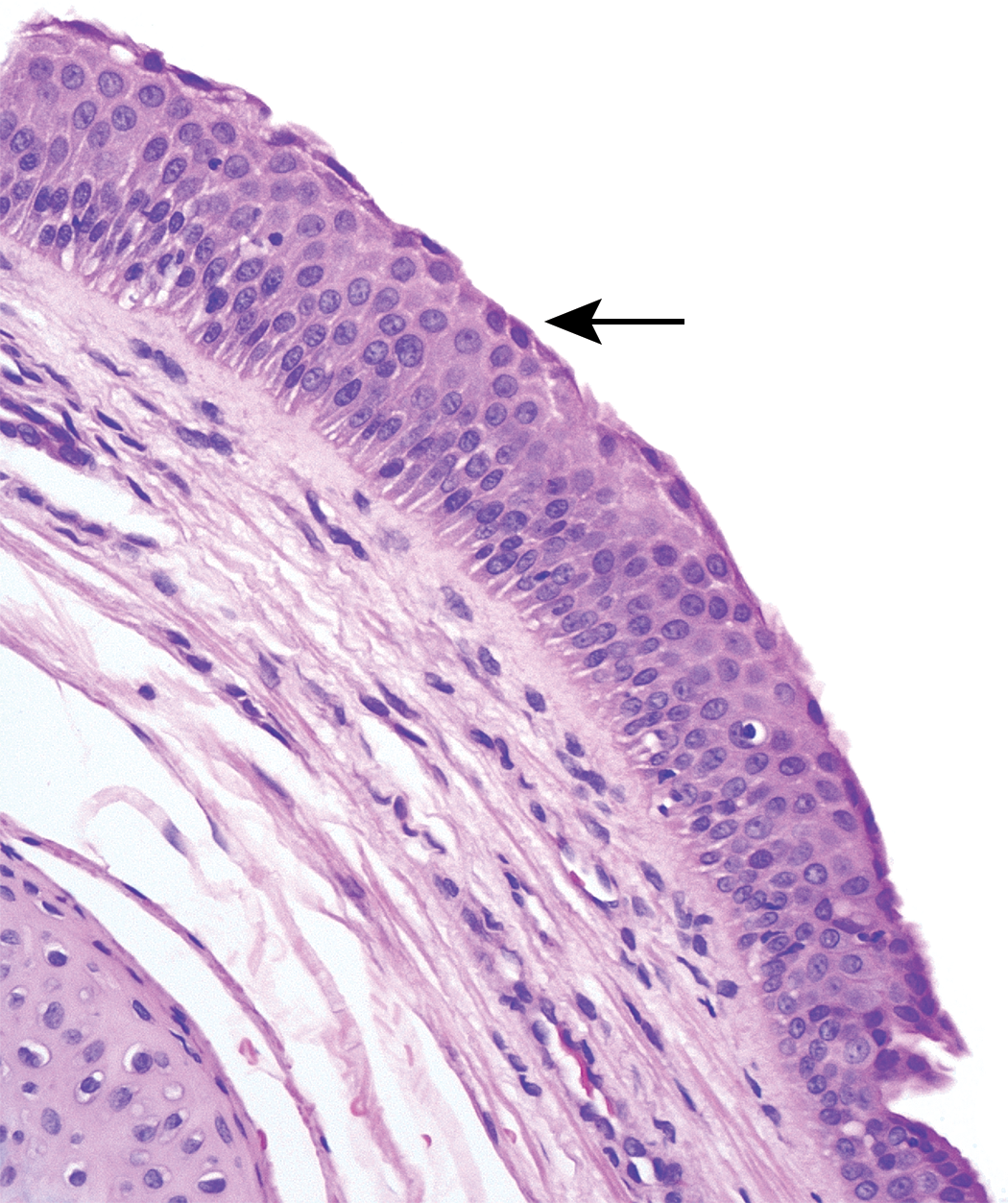
Laryngeal lesions were recorded in rodents in 24 of the 26 cases (20 rat and 4 mouse studies; Table 2). The most commonly recorded change was squamous metaplasia (Figure 1), but epithelial hyperplasia, ulceration, and mucosal inflammation were observed in a significant number of cases. These changes predominantly affected the ventral larynx, at the base of the epiglottis, with the exception of epithelial hyperplasia, which was usually present in the dorsal regions overlying the arytenoid cartilages. Necrosis of cartilage was noted in a small number of studies as a sequel to severe mucosal damage and typically involved the U-shaped cartilage supporting the ventral pouch (Figure 2).
Treatment-related Changes in Larynx and Carina.
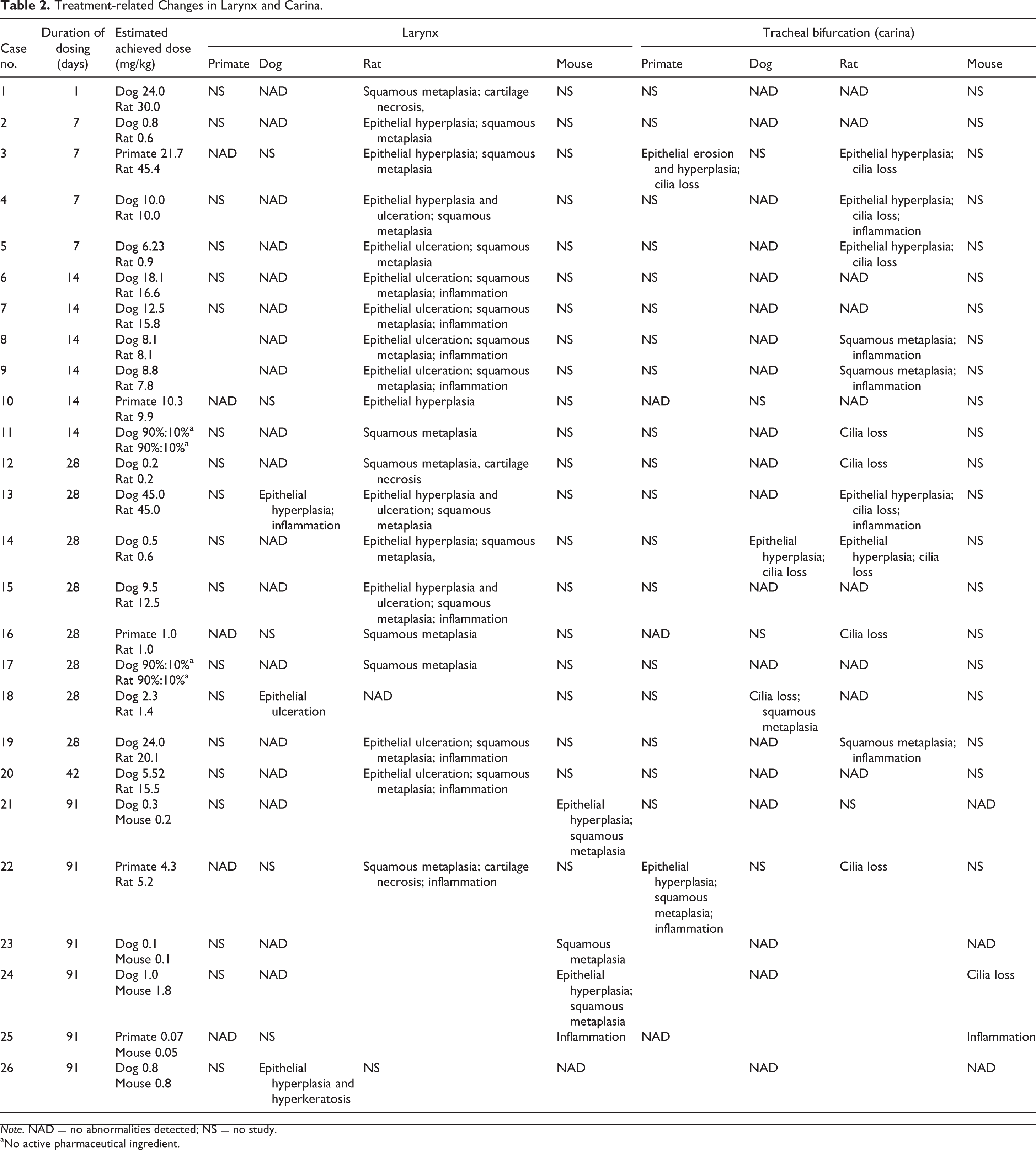
Note. NAD = no abnormalities detected; NS = no study.
aNo active pharmaceutical ingredient.
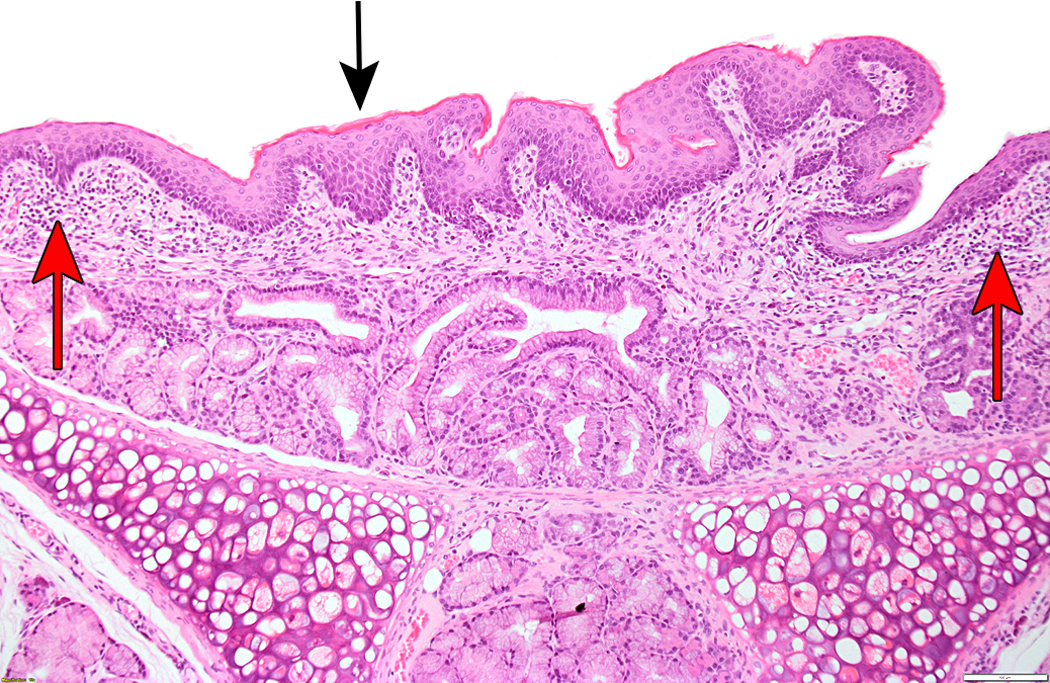
Squamous metaplasia with hyperkeratosis (black arrow) and minimal submucosal inflammation (red arrows) in rat larynx, level of epiglottis.
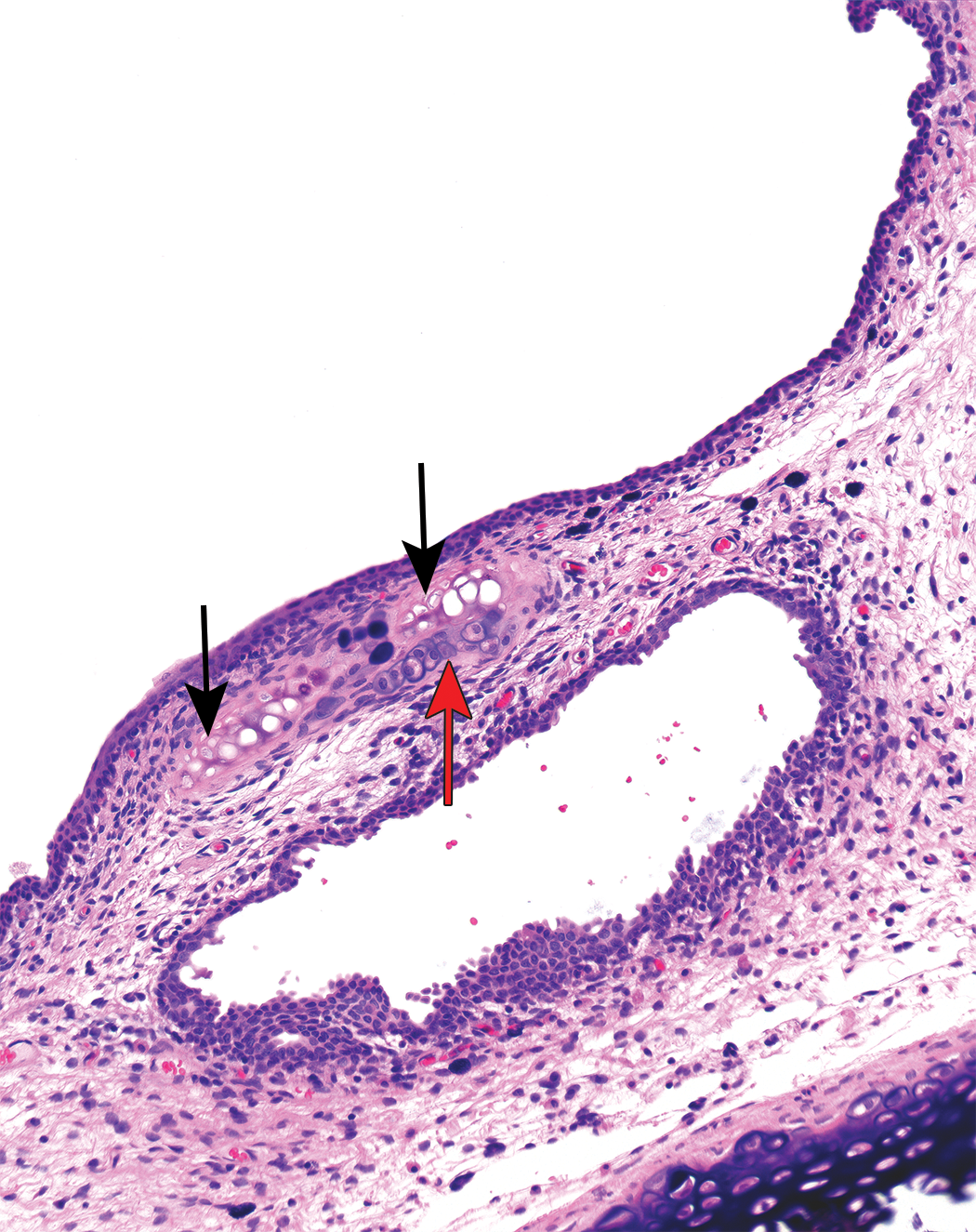
Necrotic cartilage (black arrows) with normal chondrocytes (red arrow), rat larynx.
Laryngeal findings were observed in nonrodents in only 3 of the 26 cases (3 dog studies; Table 2). These changes comprised epithelial hyperplasia, hyperkeratosis, ulceration, and mucosal inflammation (Figures 3 and 4). All these changes were present in the mucosa overlying or in close proximity to the vocal processes. Of the 3 dog studies with laryngeal changes, 2 of these were not corroborated by findings in the rodent larynx.
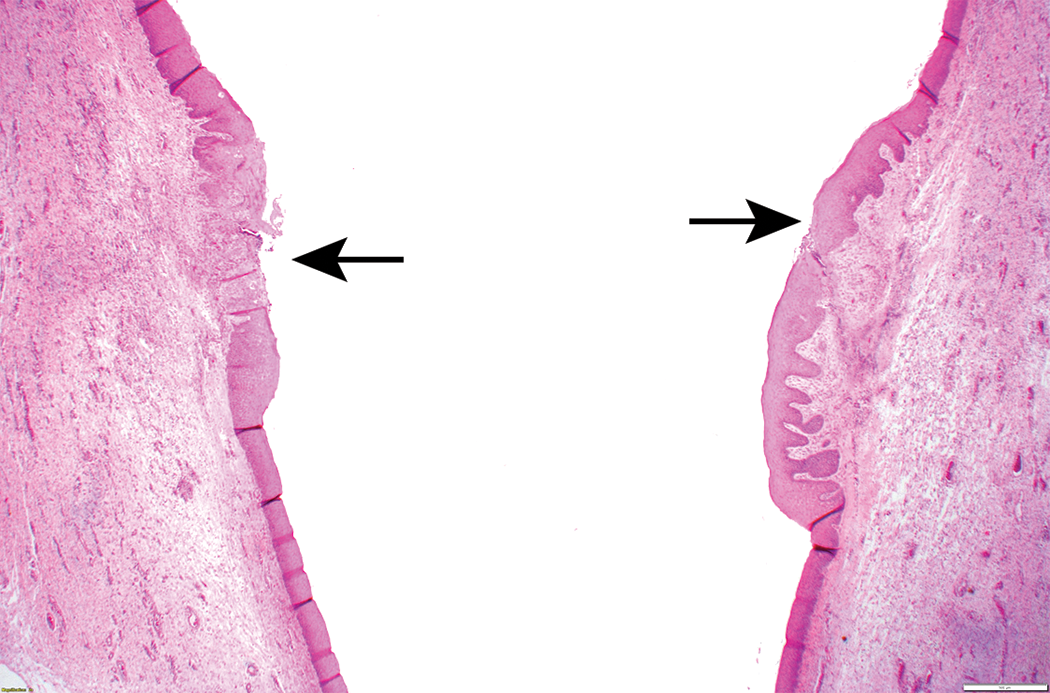
Bilateral erosion/ulceration (black arrows), epithelial hyperplasia, and inflammation of vocal cords, dog larynx.
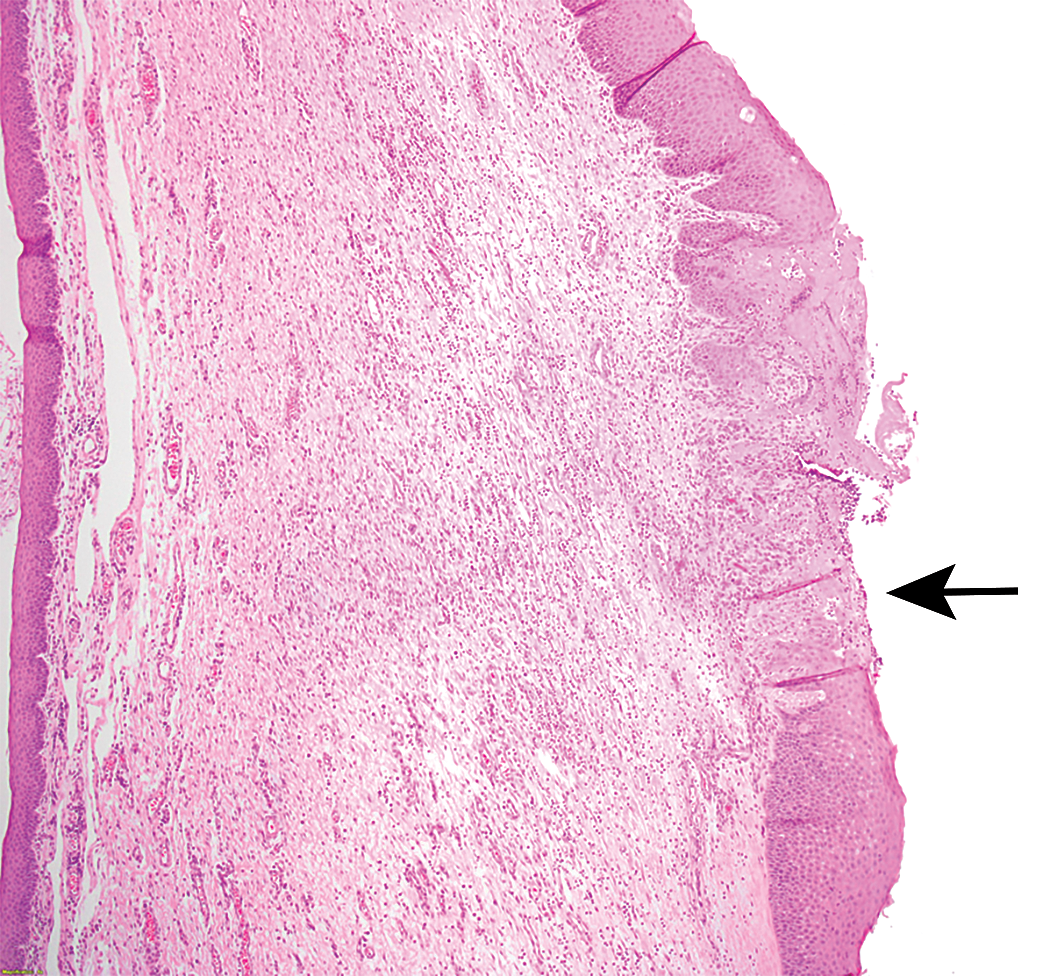
Ulceration, epithelial hyperplasia, and inflammation of vocal cord (black arrow), dog larynx.
Carina
Carinal lesions were recorded in rodents in 14 of the 26 cases (12 rat and 2 mouse studies; Table 2). The most frequent finding was loss of cilia, with epithelial hyperplasia, squamous metaplasia (Figure 5), and mucosal inflammation also being noted in these cases.
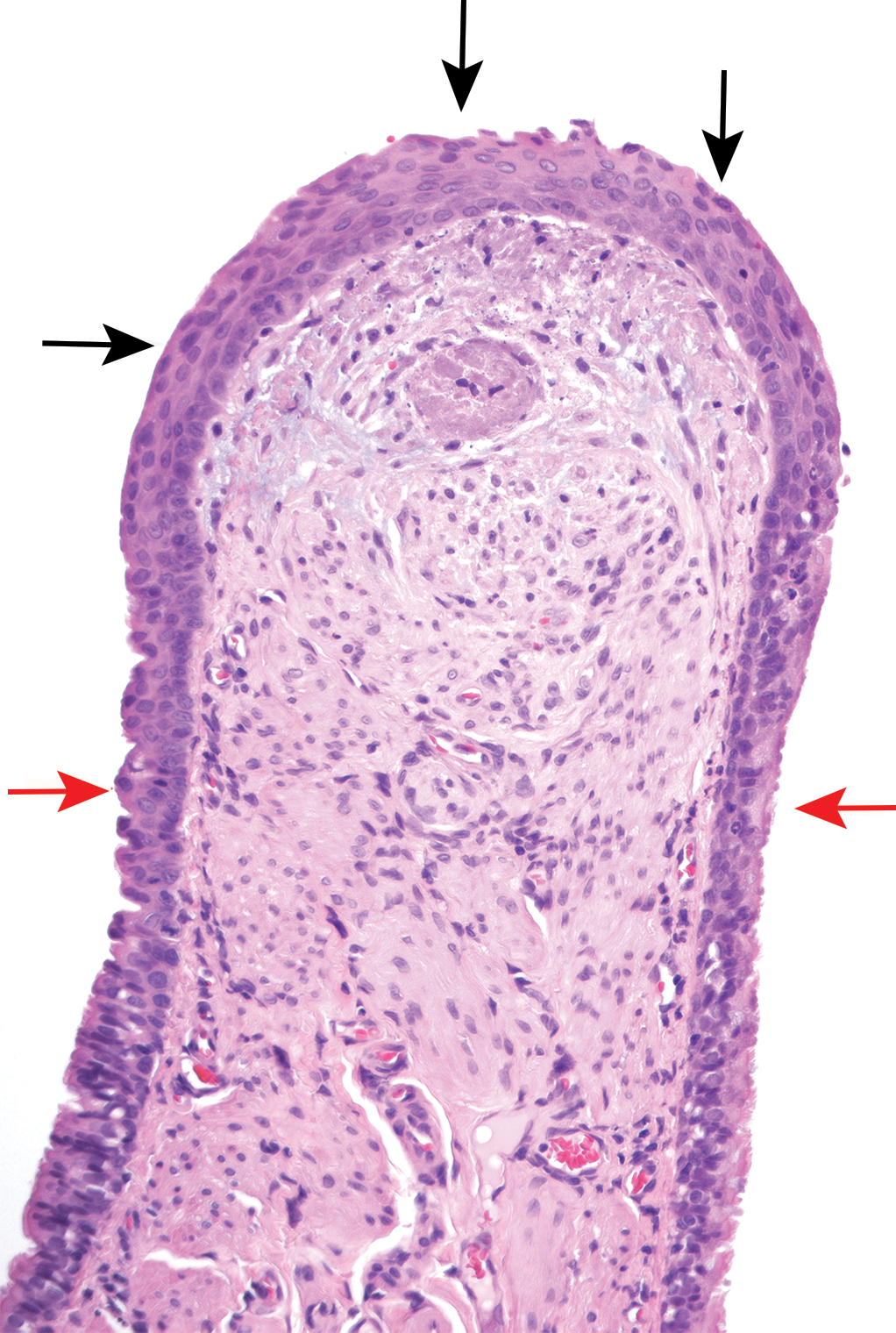
Squamous metaplasia, apex of rat carina (black arrows) with adjacent normal respiratory epithelium (red arrows).
Carinal findings were observed in nonrodents in only 4 of the 26 cases (2 monkey and 2 dog studies; Table 2). Loss of cilia, epithelial hyperplasia, and squamous metaplasia were the most frequently recorded findings with mucosal inflammation and erosions being present in the other cases. Of the 4 nonrodent studies with carinal changes, only 1 of these was not corroborated by findings in the rodent carina.
Discussion
Larynx
The rodent larynx is widely considered to be more sensitive than that of the dog and monkey in its response to inhaled xenobiotics (Osimitz, Droege, and Finch 2007), although this perception was previously unsupported by published data. In this review of 18 compounds involving 26 pairs of studies, laryngeal changes were present with much higher frequency in the rodent than nonrodent. This observation suggests that, with all other factors remaining similar, the larynx of the rodent is far more sensitive than that of the nonrodent to irritant exposure.
The reason for the susceptibility of the rodent larynx to aerosol formulations is based on several interrelated factors, but the most important of these are believed to be anatomical, airflow related, and histological (Gopinath, Prentice, and Lewis 1987; Lewis 1981).
Anatomically, the larynx is the first site of constriction in the respiratory tract, but in the rat the cranial surface of the larynx is directly targeted by the incoming airflow, as the larynx and trachea form a nearly straight line from the nasal turbinates (Kaufmann et al. 2009). This constriction, when combined with the rapid respiratory rate of the obligate nose-breathing rats, enhances the impaction of aerosols on the anterior surface of the rat larynx. In contrast, the larynx of nonrodents (including humans) is more sharply angled (approximately 90°) to the oronasal cavity, and respiration occurs through both oral and nasal cavities. These anatomical and functional differences are responsible for a reduced amount of aerosol impaction on the anterior surface of the larynx in nonrodents when compared to rats.
In general, particulate retention in the larynx is much less than in the lung. However, deposition in the rat larynx is significantly higher than one would predict from its small surface area (Lewis 1981; Raabe et al. 1977). It is probable that the rapid breathing and circuitous airflow pathway in the rat larynx lead to increased particulate deposition in comparison with other species (Werley et al. 2011). Furthermore, as metabolic rate is inversely proportional to body mass, smaller animals have relatively higher minute volumes in terms of volume/kg body weight and greater propensity for particle deposition than larger species (Phalen, Kenoyer, and David 1977).
Histologically, the mucosa of the larynx is considerably different in the rat to that of other species. In rats, there is an extensive transition zone covered by relatively thin epithelia of mixed cell types (Renne, Sagartz, and Burger 1993). In general, the thickest epithelial areas in the rodent larynx are appreciably thinner than the stratified squamous epithelial lining in the monkey or dog.
In this review, squamous metaplasia of the ventrolateral larynx was the most frequently observed finding in the rodent, which is consistent with literature reports (Osimitz, Droege, and Finch 2007). In particular, the distribution of aerosol-induced lesions in this species shows distinct predilection sites comprising the base of the epiglottis, area around the ventral pouch, and medial aspect of the arytenoid cartilages. Squamous metaplasia of the rodent larynx is generally considered to be an adaptive response and is not indicative of significant human risk (Osimitz, Droege, and Finch 2007). It is well established that toxicology data on effects in rodents may not apply to humans (Renne and Gideon 2006), but how does the larynx of beagle dog and cynomolgus monkey compare with that of human in terms of susceptibility to injury?
Generally, the larynx of the dog and cynomolgus monkey appears highly resistant to the effects of inhaled xenobiotics. As the most sensitive areas for cellular changes in animals inhaling xenobiotics are thought to be the sites of transition from relatively durable stratified squamous epithelium to a much more fragile respiratory epithelium (Renne et al. 2007), one would expect to find lesions further caudally in the larynx of dogs and primates. However, few published reports of laryngeal injury following inhalation exposure are available in these species and mostly anecdotal evidence exists of ulceration and mucosal inflammation close to the vocal processes (Renne and Gideon 2006). This impression is supported by the results of this review and by the present authors who, after many years’ experience with inhalation studies involving a wide range of compounds, have rarely seen induced lesions in the dog or monkey larynx. The conclusion must be that in these species the larynx is a relatively robust structure.
The present authors have also noted that pharmaceuticals inducing changes in rat larynx that are devoid of effects in beagle dog or cynomolgus monkeys rarely cause clinical evidence of respiratory tract irritation in humans (unpublished proprietary data). This is supported by the fact that regulatory authorities usually accept typical laryngeal changes in the rat as being nonspecific and generally not relevant to humans for pharmaceutical registration (DeGeorge et al. 1997).
Overall exposure to pharmaceutical aerosols is much higher in the nonclinical studies than the proposed human doses, with potentially higher relative deposition in rodents. The fact that laryngeal lesions are more commonly seen in rats than in dogs and cynomolgus monkeys given the same regime strongly supports the theory that the rat larynx is far more sensitive than the dog and monkey in its response to inhaled xenobiotics and will overemphasize irritant effects.
The data suggest that the human larynx is much more similar to the beagle dog and cynomolgus monkey in its response to inhaled xenobiotics, and more clinical relevance should be placed on any specific findings in these animal models
Carina
The apex of the carina at the tracheal bifurcation is routinely examined during inhalation toxicity studies in all species. In this review of 22 compounds involving 26 pairs of studies, carinal changes were present with much higher frequency in the rodent than nonrodent. This observation suggests that the carina of the rodent is far more sensitive than that of the nonrodent to irritant exposure, with all other factors remaining similar. Of the 24 rodent studies with laryngeal changes, 14 (58%) also showed changes at the carina indicating that of the 2 structures the larynx is probably the most sensitive.
Pathologists routinely engaged in inhalation studies acknowledge that incidental findings at the carina in untreated animals are not uncommon, although these data are largely unpublished. In this review, loss of cilia was the most frequently observed drug-induced carinal lesion, followed by epithelial hyperplasia, inflammation, and squamous metaplasia. In the literature, squamous metaplasia is the most commonly reported spontaneous finding at this site (Figure 6; Sato et al. 2012).
Squamous metaplasia (black arrow), apex of dog carina.
The carinal apex is a well-defined point of impaction for inhaled particles, and induced lesions at this site seem entirely predictable. Additionally, the efficiency of the mucociliary transport mechanism in vivo is also reduced at airway branching (Martonen 1992), further increasing the potential for increased retention at these points and thus the sensitivity of the site.
The observation that the rodent carina is more frequently affected than that of the nonrodent by lesions due to impaction can simply be explained by the disparity in mucosal thickness at this site, with the epithelium of the rat being approximately 3 to 4 times thinner than that of the dog (Pilling, personal observation). Therefore, it seems entirely justifiable that the typical lesions seen in the rat carina have a similar etiology to those seen in the larynx and can be regarded in the same manner for assessing potential clinical implications of an inhaled pharmaceutical.
In summary, the premise that the larynx of rodents is more sensitive than that of nonrodents is substantiated by data compiled from 52 inhalation studies which indicate a far higher incidence of drug-induced laryngeal lesions in rodents than in the dog or primate. The carinal apex at the tracheal bifurcation is also a well-defined point of impaction for inhaled particles and is more frequently injured in rodents than in dogs and primates, due to its relatively thin epithelium and airflow dynamics. Anatomically and histologically, the human larynx resembles that of the nonrodent more closely than the rodent and the relevance of rodent findings in these studies for humans is doubtful. More emphasis should be placed on any specific findings in nonrodent animal models than in rodent models when assessing potential clinical implications of an inhaled pharmaceutical.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.