
Abstract
Neurohistopathological changes in the brain were assessed in juvenile beagle dogs given vigabatrin at 30 or 100 mg/kg/day by oral gavage from postnatal day 22 (PND22) until 16 weeks of age (PND112), when brain myelination is considered to reach the adult stage in dogs. Separate subgroups were treated from PND22 to PND35 or PND36 to PND49 to assess early effects. In addition to extensive brain histopathology, there were assessments of toxicokinetics, clinical condition, body weight, organ weights, and macroscopic pathology. In animals treated for 14 days from PND22, minimal or slight vacuolation was seen in the neuropil of the septal nuclei, hippocampus, hypothalamus, thalamus, cerebellum, and globus pallidus at 100 mg/kg/day and minimal vacuolation in the thalamus, globus pallidus, and cerebellum at 30 mg/kg/day. In animals given 100 mg/kg/day for 91 days from PND22, minimal or slight vacuolation was observed only in the hippocampus, hypothalamus, and thalamus. No vigabatrin-related brain vacuolation was observed in animals given 30 or 100 mg/kg/day for 14 days from PND36. Clear evidence of recovery was observed after 14-day and 6-week off-dose periods that followed treatment from PND22 to PND35 or PND22 to PND112, respectively.
Introduction
Vigabatrin (marketed as SABRIL®) is an antiepileptic drug indicated for infantile spasms (IS) in children aged 1 month to 2 years. Administration of vigabatrin to neonatal rats in a previous study (Rasmussen et al. 2015) was associated with changes in the gray matter of the brain and decreased myelination, which were considered distinct from findings in adult animals given this drug. These findings were observed at doses associated with plasma concentrations lower than those achieved clinically in infants and children. Vigabatrin has, however, been demonstrated to show significant clinical benefit in the treatment of IS, a serious disease for which there was no approved therapy. It was approved for use in pediatric patients for whom the potential benefits outweighed the risk of adverse effects, but this continued use was subject to a regulatory requirement for a juvenile animal toxicology study in a nonrodent species.
The objective of the study reported here was the assessment of neurohistopathological changes in the brain during an oral gavage study in juvenile (preweaned) beagle dogs over a treatment period of up to 91 days. Dosing commenced at postnatal day 22 (PND22) and continued until the animals reached 16 weeks (PND112) of age; this overall period of treatment corresponded approximately to administration in rats from PND10 up to PND38 and in children from 1 month up to around 9 years of age. The 91-day treatment period was followed by an off-dose period of 6 weeks to assess the potential for recovery from any changes seen. In addition, in order to assess the potential for neurohistopathological changes that might arise specifically in the period up to around the age of weaning, which occurs in dogs at around 6 to 8 weeks of age, a second subgroup of animals were dosed with vigabatrin from PND22 to PND35 and a third subgroup from PND36 to PND49; each of these dose phases was followed by an off-dose period of 14 days and were selected as they represent notable development milestones including communication and social play, the end of completion of sensory and reflex and sleep cycle development, and completion of the fear response (Miot-Noirault et al. 1997; Robinson, Smith, and Viau 2012).
The study was conducted in accordance with the requirements of current, internationally recognized Good Laboratory Practice Standards, and the applicable sections of the United Kingdom Animals (Scientific Procedures) Act 1986, as well as the animal welfare policy of H. Lundbeck A/S. The dog was chosen as the test species because it is accepted as a predictor of toxic change in man and meets the requirement for a nonrodent species by regulatory agencies. The beagle dog was used because of the historical control data available and because this species has been used as the nonrodent model in previous toxicity studies on vigabatrin. The oral route of administration was chosen to simulate the conditions of clinical administration, as this route of administration has generally been used in animal safety testing on this compound.
Materials and Methods
Twelve pregnant female Beagle dogs of known Harlan interfauna lineage were obtained from Harlan (Harlan Laboratories UK Ltd, Sawtry Way, Wyton, Huntingdon, Cambs, PE28 2DX, UK) and delivered to Huntingdon Life Sciences no later than day 40 of gestation. They were allowed to give birth naturally (gestation days 61–67). Litters were allocated soon after parturition in a structured but randomized manner to ensure, as far as possible, an equitable distribution across the dose groups and phases. All available pups were allocated to the study in anticipation of subsequent preweaning losses. Parental bitches were housed with their offspring until weaning and then reassigned for appropriate investigations or rehomed to a suitable household. The majority of litters included animals from all 3 dose groups, including controls. Cross-fostering was not practicable in dogs due to the strong possibility of rejection and the age differences expected between the litters. Litter mates were spread across the dose groups to avoid the potential bias that was likely to occur if all of a litter was assigned to the same dose group. This bias might occur if there were genetic susceptibilities within a parental line or where there were differences in the general rate of growth and development due to variation in maternal care/capabilities. The presence of multiple dose levels (including controls) within a single litter provided the potential for cross-contamination, but steps were taken to minimize this and 2 litters were allocated solely to the control group in case of a need to exclude data from controls severely contaminated with test article. No such serious contamination was evident, although there was some minor cross-contamination.
Parental bitches were fed with Harlan Teklad 2027 (27% protein) Dog Maintenance Diet throughout. Pups received supplementary “Lamlac” milk powder (concentrated milk protein) from approximately 3 weeks of age. Milk was supplied once daily from approximately 3 weeks of age and then twice daily from approximately 4 weeks of age for at least 4 weeks. The supply of milk was reduced back to once a day during the period when the animals were approximately 10 to 11 weeks old and then no milk was given. Pups received Harlan Teklad 2027 (27% protein) Dog Maintenance Diet during the weaning period and until they were approximately 9 to 10 weeks old, Harlan Teklad 2025/2027 (25%/27% protein) Dog Maintenance Diet for approximately 2 weeks and then Harlan Teklad 2025 for approximately 4 weeks, Harlan Teklad 2021/2025 for 2 days, and finally Harlan Teklad 2021 (21% protein) Dog Maintenance Diet until termination. The body weight of the animals was reviewed regularly and the feeding regimen was amended accordingly. Potable water from the public supply was always available via automatic valves, which were checked daily, and fresh water provided in bowls for the pups until they were tall enough to reach the valves.
The animals were housed in a limited access purpose built facility. Parental females were removed from the pen on a daily basis for approximately 2 hr and taken to a separate area for a period of exercise; after dosing of the pups commenced, this period coincided with dose preparation and administration time for the offspring. Litters were given a period of exercise from 3 to 5 weeks of age onward. Once the pups were weaned, they were then reallocated into pens containing up to a maximum of 8 animals of the same dose group.
Dose Selection and Administration
Animals were administered vigabatrin (batch No. 120017) using water for injection as the vehicle. The pups were dosed once daily at approximately the same time each day with constant doses (in terms of mg/kg body weight/day) using a suitably graduated syringe and a rubber catheter inserted via the mouth and down the esophagus. The dose volume was 5 ml/kg body weight, calculated from the most recently recorded scheduled weight. To minimize the chance of cross-contamination under study conditions, pups were separated from the dam for at least 1 hr before dose administration and not allowed access for suckling for at least 1 hr after dosing. There was a period of gradual acclimatization to this separation in the last week of the predose period.
The high dose of 100 mg/kg/day set for this investigation was the mid-dose used on the original 3-month dog study (Megel 1979). In the older (adult) dog, this dose was well tolerated, no premature terminations occurred, and brain findings were detected at histopathological examination. A preliminary toxicity study in the preweaned juvenile dog (Bottomley 2013) demonstrated that 100 mg/kg/day was well tolerated in beagles when administered from PND22 for at least 6 weeks and provided evidence of good systemic exposure with no discernible difference in the exposure to male and female pups. The low dose on this study was set at 30 mg/kg/day, which was the lowest dose administered on the original 3-month study in older dogs and which was anticipated to provide an exposure level in the range of that experienced in young humans.
The dosing periods are given in Table 1 and were selected based on available knowledge on the central nervous system development of pups and juvenile dogs. Treatment was started on day 22, which corresponds to approximately 1 month of age in human infants, the earliest age for administration of SABRIL® to patients. It also corresponds approximately to 10 days in rats (Buelke-Sam 2003) and is in alignment with a previously published study on vigabatrin in juvenile rats (Rasmussen et al. 2015). Treatment was ended on day 112, which corresponds to week 16 after birth, where the dog brain appears almost fully developed, as cognitive functions are well advanced and brain myelination has reached the adult stage (Garman 2003; Miot-Noirault et al. 1997). The treatment period in dogs thus exceeded the juvenile human treatment period to allow for an assessment of the entire expected dog myelination period. The intermediate end point of day 35 (intermediate I) and day 49 (intermediate II) were selected, as these represent notable development milestones including communication and social play as well as covering the end of completion of sensory and reflex and sleep cycle development (day 35) and completion of the fear response (day 49) (Robinson, Smith, and Viau 2012; Miot-Noirault et al. 1997). Day 47 also approximates to PND30 in rats, thus matching the juvenile rat toxicity study (Rasmussen et al. 2015). A recovery period of 2 weeks was allowed for some of the animals administered vigabatrin in the periods PND22 to 35 or 36 to 49 while some of the animals administered vigabatrin during PND22 to 112 were allowed a recovery period of 6 weeks. A recovery period was included in this study, as extensive recovery from vigabatrin-induced changes has been seen in other animal species (Rasmussen et al. 2015). A visual representation of the time periods chosen is given in Figure 1.
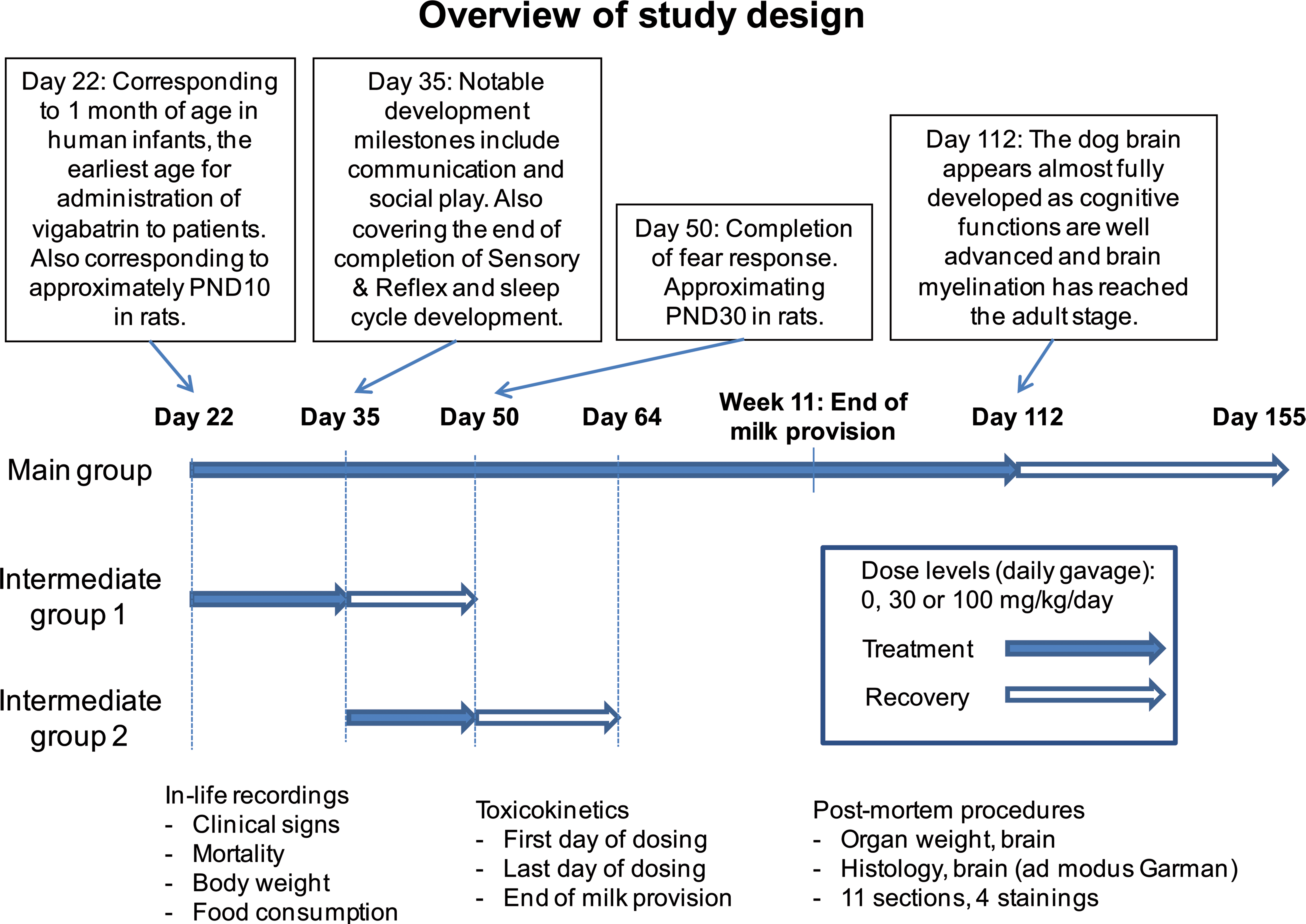
Overview of study design.
Overview of study design (dosing days, animal nos., group nos., and vigabatrin dose levels).
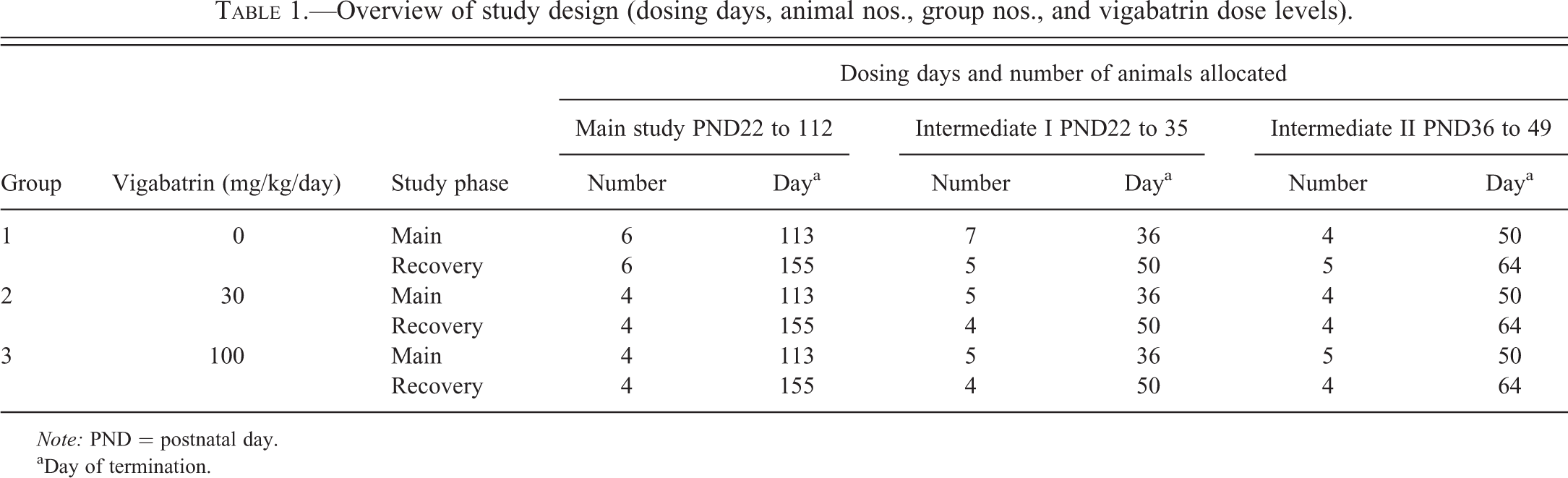
Note: PND = postnatal day.
aDay of termination.
Serial Observations
Blood samples for toxicokinetics were obtained, via the jugular vein, predose, and 0.5, 1, 2, 4, 8, 12, and 24 hr after administration on the first and last day of dosing for all main study pups (each phase) and also from the 91-day study animals at the end of the period of milk provision, to assess the impact of regular milk provision on bioavailability.
Animals were inspected visually at least twice daily for evidence of ill-health or reaction to treatment. Pens were inspected daily for evidence of ill-health among the occupants.
Detailed observations were recorded daily during and immediately after dosing, ½ to 2 hr after completion of dosing and as late as possible in the working day. A detailed weekly physical examination was performed on each animal to monitor general health.
The weight of each animal was recorded (prior to feeding) twice weekly during the acclimatization period, on the day that treatment commenced (day 1), twice weekly throughout the study, and before necropsy. Once the pups were weaned and separated from the maternal dam, food consumption was monitored by visual assessment of the residues in the food hoppers.
At termination, all pups were euthanized with an overdose of sodium pentobarbitone solution (200 mg/ml) by intravenous injection followed by exsanguination and subjected to a detailed necropsy. After a review of the history of each animal, a full macroscopic examination of the tissues was performed. All external features and orifices were examined visually. Any abnormality in the appearance or size of any organ and tissue (external and cut surface) was recorded, and the required tissue samples were preserved in appropriate fixative. The brain was weighed and a full regulatory list of tissues retained in fixative.
The brains from all animals from the main study (91-day study) and from those allocated to intermediate I and intermediate II sacrifices were immersion fixed in 10% neutral-buffered formalin for 48 to 72 hr before being processed to paraffin block. Following this limited period of immersion fixation, little or no fixation artifact was observed in histological sections. Brains from animals assigned to the control and 100-mg/kg/day dose groups (groups 1 and 3) of the 91-day study, intermediate I and intermediate II treatment and recovery phases and group 2 of the 91-day study, and intermediate I treatment and recovery phases were sampled (Garman 2003) to provide 6 transverse sections of the entire cerebrum, 1 of the cerebellum and 3 of the brain stem. Sections (4–5 µm) at each level were stained with hematoxylin and eosin, toluidine blue, Fluoro-Jade B, and Luxol fast blue. Hematoxylin and eosin was used as the standard pathology stain for examination of the general features of the brain sections, toluidine blue was used for closer examination of neurons, Luxol fast blue for examination of myelin, and Fluoro-Jade B for identification of degenerating neurons. The brain was examined for all animals sacrificed on completion of the 91-day, intermediate I and intermediate II periods in line with current practice. Findings were either reported as “present” or assigned a severity grade. In the latter case, 1 of the following 5 grades was used: minimal, slight, moderate, marked, or severe. A reviewing pathologist undertook a peer review of the microscopic findings, following guidelines published by the Society of Toxicologic Pathologists (Morton et al. 2010). Pathological examination was not blinded.
Results
Data on the systemic exposure to vigabatrin are given in Table 2. All treated animals were systemically exposed to vigabatrin and in general area under the curve and C max values increased with dose. Some samples from control animals had concentrations of vigabatrin above lowest level of quantification, but levels were very low compared to exposure in the low-dose group (30 mg/kg/day). On each occasion of examination (days 1, 14, 42, and 91), systemic exposure to vigabatrin for animals from the low-dose group was generally at its highest between 30 min and 4 hr after dosing and showed a clear descending exposure profile; plasma exposure during this period typically being in the range 3,000 to 38,000 ng/ml. For the control animals, the positive results seen in this period were in the range of 14.0 to 83.1 ng/ml and did not show the typical profile seen in dosed animals. It is clear that the contamination in control animals was extremely low compared with the low-dose group and was not associated with dose administration. The contamination most likely occurred during sample handling following removal of blood from the animals and/or reflected that animals from various dose groups had been housed together. There were no notable differences in exposure between the different vigabatrin-treated subgroups with respect to study days and sex, and there appeared to be no accumulation during dosing (accumulation indices being approximately 1).
Systemic exposure to vigabatrin (group nos., vigabatrin dose levels, C max, AUC, and AIobs values).
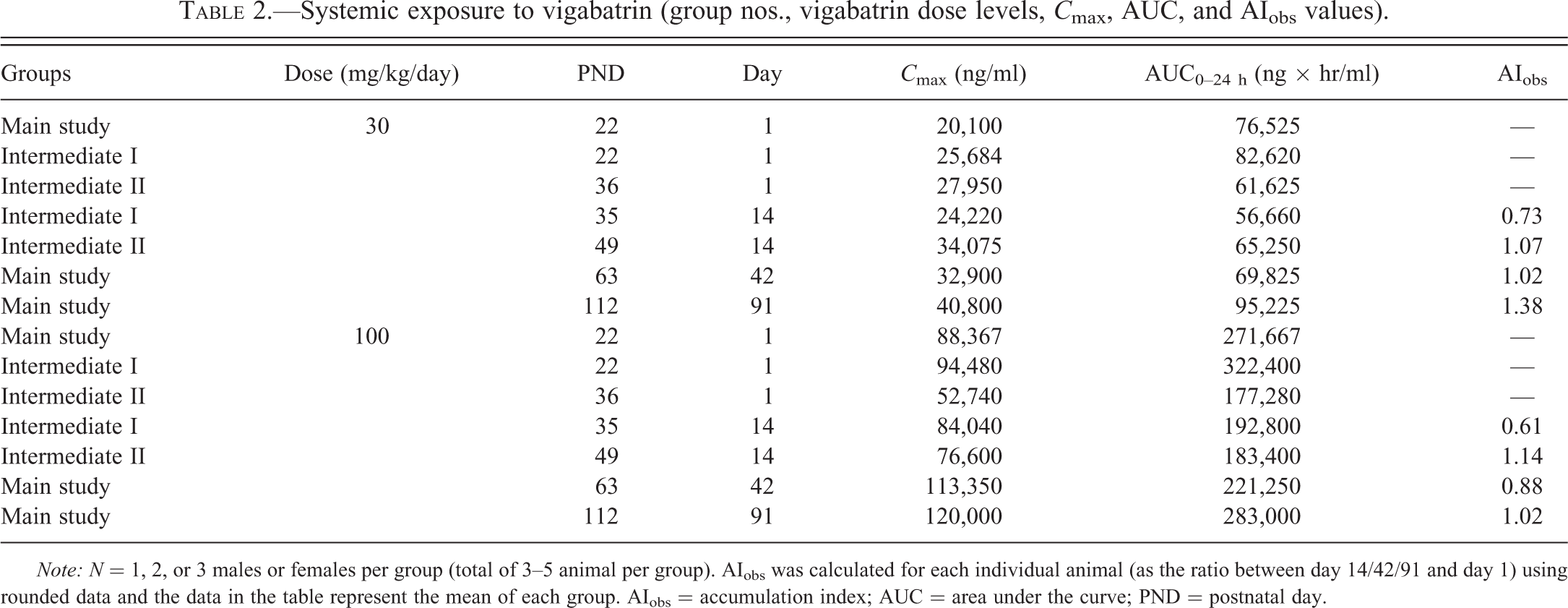
Note: N = 1, 2, or 3 males or females per group (total of 3–5 animal per group). AIobs was calculated for each individual animal (as the ratio between day 14/42/91 and day 1) using rounded data and the data in the table represent the mean of each group. AIobs = accumulation index; AUC = area under the curve; PND = postnatal day.
There were no clinical signs indicative of a reaction to treatment and there were no effects of treatment with vigabatrin on growth (no statistical significance). The brain weights recorded at the end of the main or recovery periods for animals assigned to the 91-day phase, intermediate I or II phases, of this study were similar across the dose groups and there were no differences considered to be associated with the administration of vigabatrin (Table 3). The macroscopic examination performed on animals from each treatment phase revealed no intergroup differences of note (data not presented).
Summary of brain weights for all dose phases.
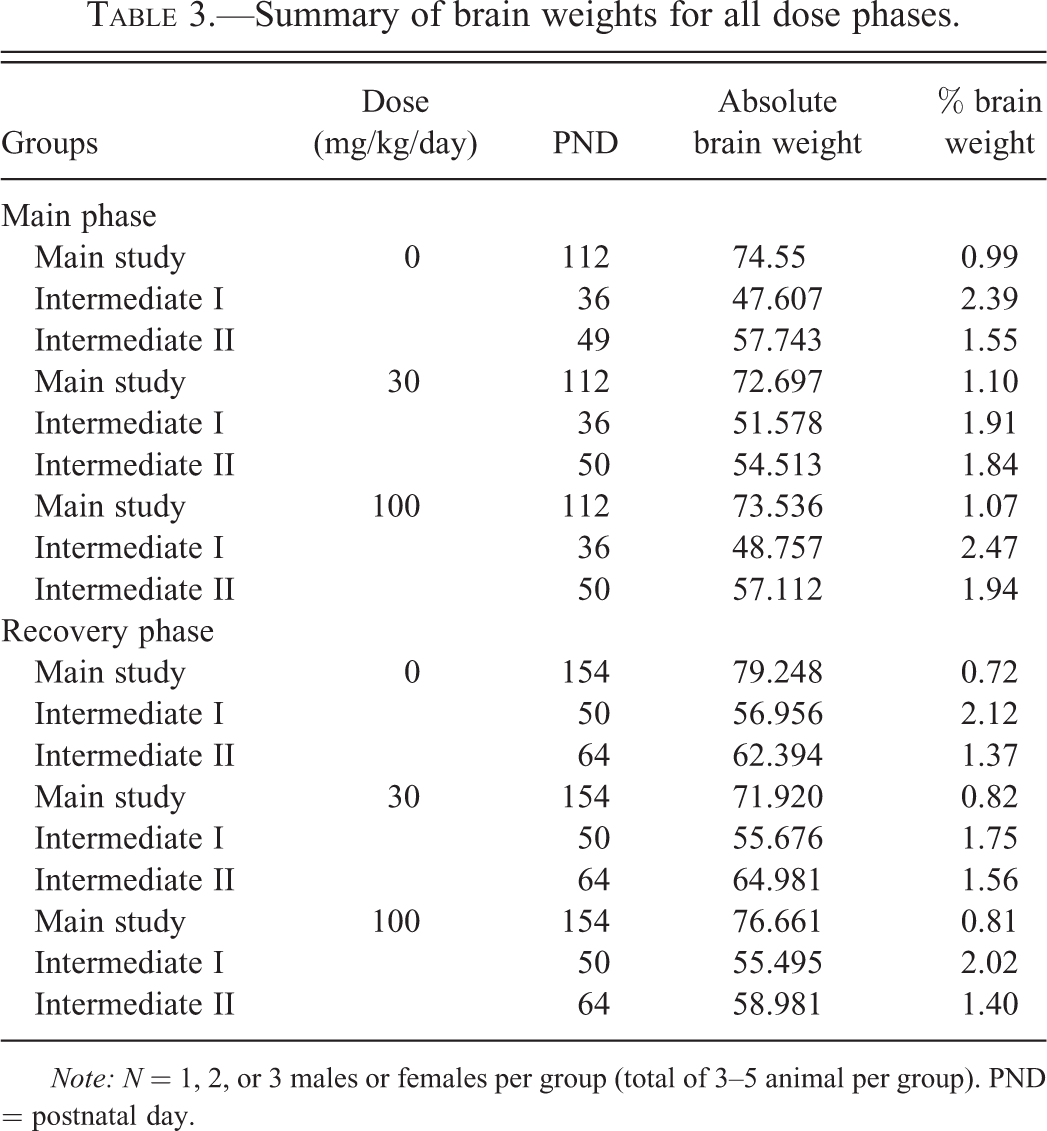
Note: N = 1, 2, or 3 males or females per group (total of 3–5 animal per group). PND = postnatal day.
A detailed account of the microscopic findings in the brains of animals administered 100 mg/kg/day vigabatrin is given in Table 4, and a summary of the microscopic findings in the brain from all animals is given in Table 5. Minimal or slight vacuolation was observed in the neuropil of the septal nuclei (Figure 2), hippocampus (Figure 3), hypothalamus and thalamus, and adjacent to cerebellar nuclei (Figure 4) in animals that received 100 mg/kg/day from PND22 to PND35. Minimal or slight vacuolation was also observed in the myelinated fiber tracts of the globus pallidus (Figure 5) in animals from this treatment group. In animals of this phase that received 30 mg/kg/day, minimal vacuolation was observed in the myelinated fiber tracts of the globus pallidus in one animal, and the neuropil of the thalamus and adjacent to cerebellar nuclei in another. The vacuolation was visible on the toluidine blue– and Luxol fast blue–stained sections at the same location and severity as seen on the H&E-stained sections. There was no evidence of a decrease in myelin staining in the Luxol fast blue–stained sections of brain from treated animals when compared with controls and no signs of neurodegeneration in the Fluoro-Jade B-stained sections.
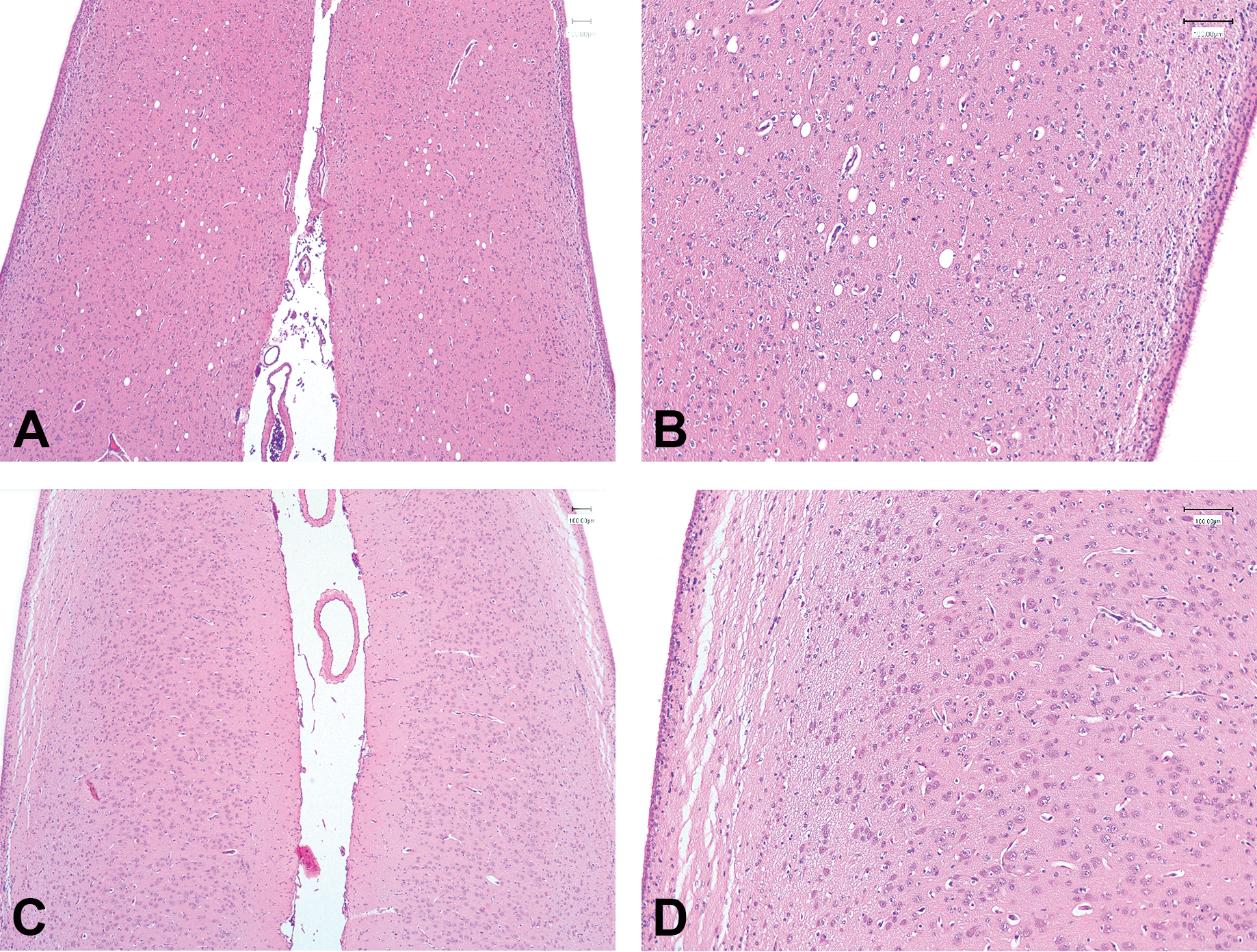
Vacuolation of the septum seen in an animal treated with 100-mg/kg/day vigabatrin for 14 days (PND22–35). Representative images of H&E-stained sections of the septum from an animal treated with 100 mg/kg/day vigabatrin (A, B) and an age-matched control from the same cohort (C, D). (A, C) Original objective magnification 4×; (B, D) original objective magnification 10×.
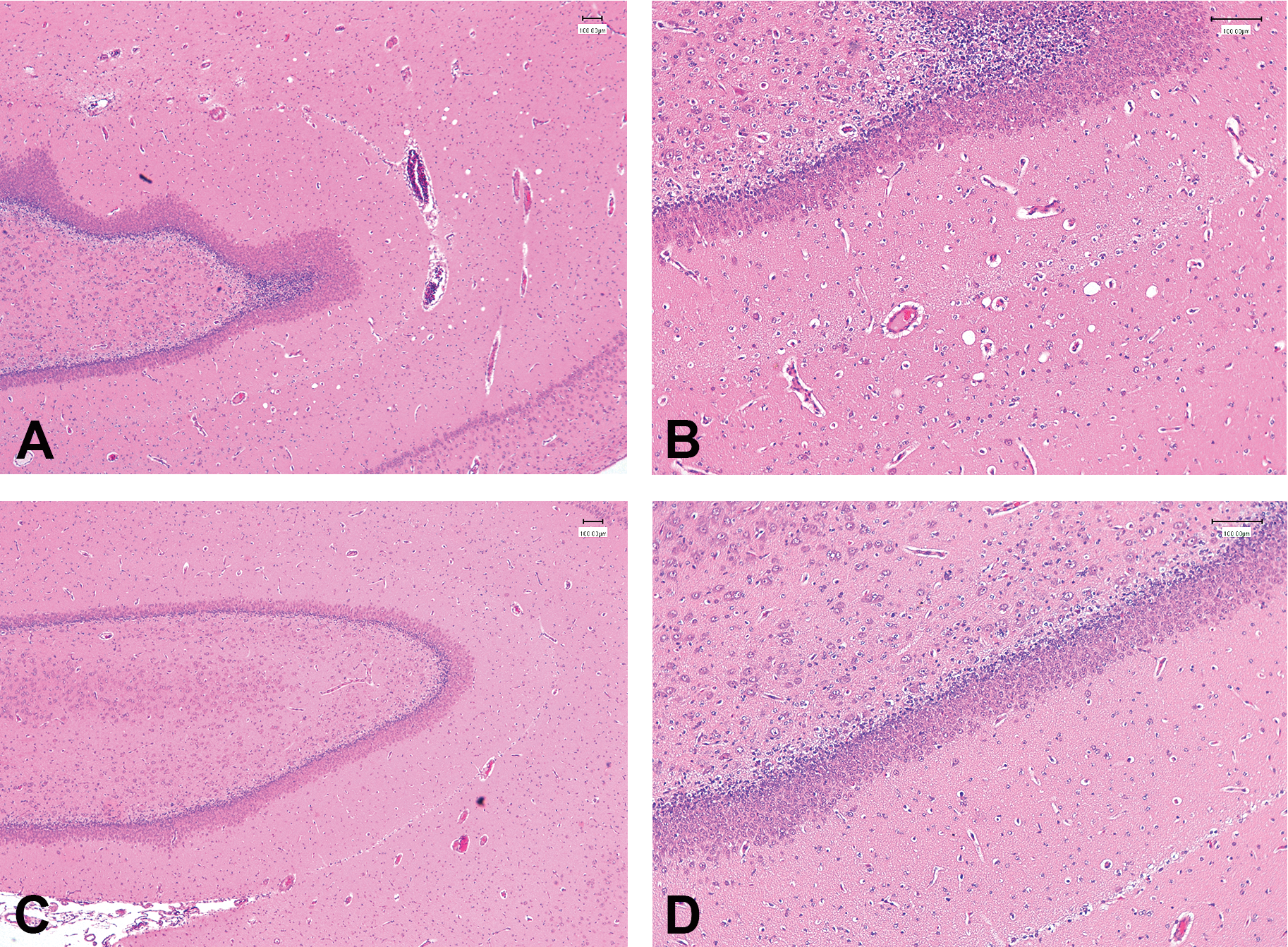
Minimal vacuolation of the hippocampus seen in an animal treated with 100-mg/kg/day vigabatrin for 14 days (PND22–35). Representative images of H&E-stained sections of the hippocampus from an animal treated with 100-mg/kg/day vigabatrin (A, B) and an age-matched control from the same cohort (C, D). (A, C) Original objective magnification 4×; (B, D) original objective magnification 10×.
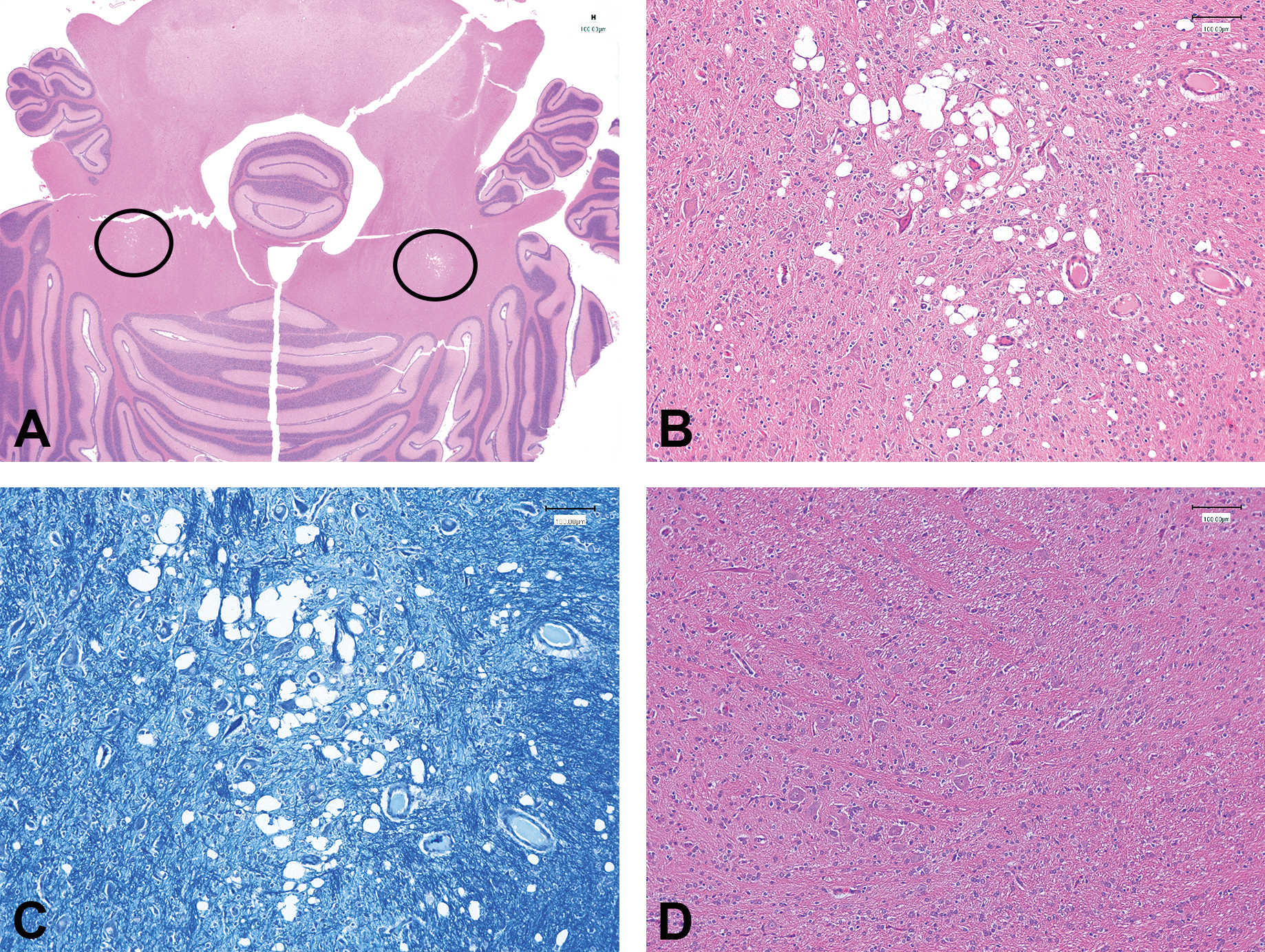
Bilateral focal vacuolation of the neuropil, adjacent to cerebellar nuclei, seen in an animal treated with 100-mg/kg/day vigabatrin for 14 days (PND22–35). (A) Image from an H&E-stained section of the cerebellum, showing bilateral location of vacuolated areas (circled) from an animal treated with 100-mg/kg/day vigabatrin; (B) image from the same H&E-stained section as (A) showing detail of the vacuolated area. (C) Image of Luxol fast blue–stained section from the same animal and area of the brain as (B). (D) Image from H&E-stained section taken from the same area as for (B) from a control animal from the same cohort as (B). (B, C, D) Original objective magnification 10×.
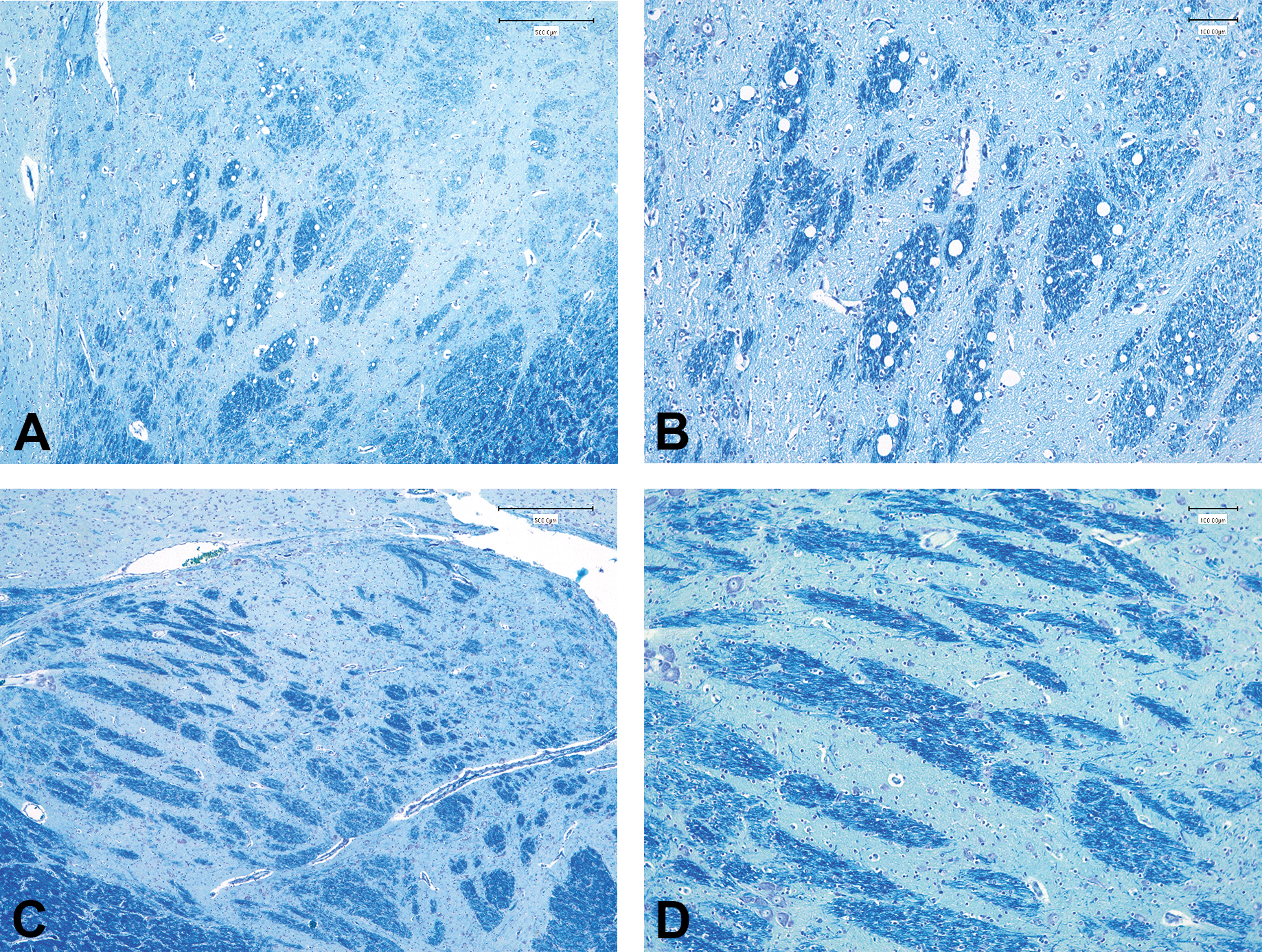
Vacuolation of the myelinated fibers of the globus pallidus seen in an animal treated with 100-mg/kg/day vigabatrin for 14 days (PND22–35). Representative images of Luxol fast blue–stained sections of the globus pallidus from an animal treated with 100-mg/kg/day vigabatrin (A, B) and an age-matched control from the same cohort (C, D). (A, C) Original objective magnification 4×; (B, D) original objective magnification 10×.
Summary of brain findings for all dose phases (100 mg/kg/day only).
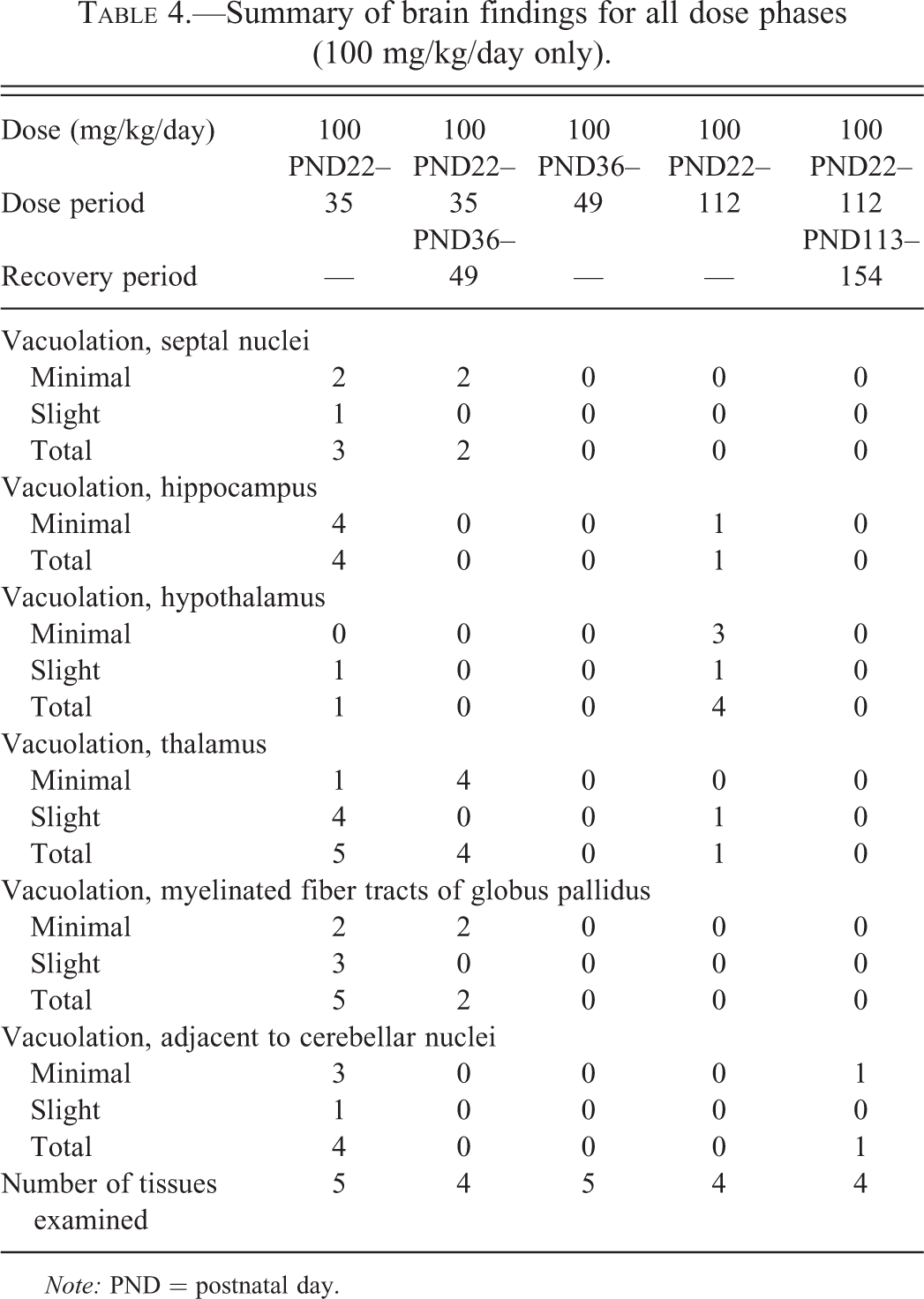
Note: PND = postnatal day.
Overview of the presence of vacuoles in dog brains in relation to dose levels and treatment period (recovery given in brackets).

Note: N/A = not applicable; PND = postnatal day.
After a 2-week off-dose period (PND36–PND49), vacuolation was observed in the thalamus of all animals that had previously received 100 mg/kg/day and in the myelinated fiber tracts of the globus pallidus and septal nuclei of 2 animals. Vacuolation was observed in the myelinated fiber tracts of the globus pallidus of 1 animal that had previously received 30 mg/kg/day. In all cases, the vacuolation was of minimal severity. The vacuolation was visible on the toluidine blue– and Luxol fast blue–stained sections at the same location and severity as seen on the H&E-stained sections. There was no evidence of a decrease in myelin staining in the Luxol fast blue–stained sections of brain from treated animals when compared with controls and no signs of neurodegeneration in the Fluoro-Jade B-stained sections. Minimal vacuolation of the accumbens nucleus was only observed in 1 control male.
There were no findings considered to be related to treatment with vigabatrin among pups that were dosed from PND36 to PND49. For pups that were dosed from PND22 to PND112, minimal or slight vacuolation was observed in the neuropil of the hypothalamus from all animals that received 100 mg/kg/day. In 1 animal, vacuolation was also observed in the hippocampus and thalamus. The vacuolation was visible on the toluidine blue– and Luxol fast blue–stained sections at the same location and severity as seen on the H&E-stained sections. There was no evidence of a decrease in myelin staining in the Luxol fast blue–stained sections of brain from treated animals when compared with controls, and no signs of neurogeneration was seen in the Fluoro-Jade B-stained sections. No changes were observed in animals that received 30 mg/kg/day. After a 6-week off-dose period, minimal vacuolation was observed in the neuropil adjacent to the cerebellar nuclei in 1 animal receiving 100 mg/kg/day. The vacuolation was visible on the toluidine blue– and Luxol fast blue–stained sections at the same location and severity as seen on the H&E-stained sections. No decrease in myelin staining was apparent in the Luxol fast blue–stained sections of brain from treated animals when compared with controls. No fluorescent staining of neuronal tissue was seen on the Fluoro-Jade B-stained sections. No vacuolation of the brain was observed in animals receiving 30 mg/kg/day.
Discussion
Vigabatrin is generally considered to have a relatively low degree of toxicity in rats and dogs (NDA 20-427 1994), and the highest dose level of 100 mg/kg/day caused no detectable behavioral effects in juvenile dogs in this study, in line with previous knowledge on adult dogs (Gibson et al. 1990). While juvenile rats do appear to be more sensitive than adult rats with respect to vigabatrin-induced changes in clinical signs, body weight, and eventually death (Rasmussen et al. 2015; Walzer et al. 2011), the data presented here do not allow for a similar comparison to be made in dogs because of the dose levels chosen.
Administration of 30- or 100-mg/kg/day vigabatrin resulted in maximum plasma concentrations of 20 to 41,000 ng/ml and 53 to 120,000 ng/ml. This corresponds well with the approximate C max values of 30 to 50,000 ng/ml observed in juvenile rats when administered 50-mg/kg/day vigabatrin (Rasmussen et al. 2015; Walzer et al. 2011). In patients, the mean plasma concentration of vigabatrin was found to be 42,000 ng/ml (Sánchez-Alcaraz et al. 1996) and C max values of approximately 65,000 ng/ml have been recorded in young children following administration of 50 mg/kg (Battino, Estienne, and Avanzini 1995). The data suggest that oral availability of vigabatrin is similar between humans and laboratory rats and dogs, linearly related to dose and close to complete. In monkeys, the oral vigabatrin absorption appears to be lower and even dose dependent, as the absorption goes down from 44% to 25% when the oral dose is increased from 50 to 300 mg/kg/day (Gibson et al. 1990).
Oral gavage administration of vigabatrin to juvenile beagle dogs from PND22 at dose levels of 30 or 100 mg/kg/day was associated with mild vacuolation in the brain. In animals treated for 14 days from PND22, minimal or slight vacuolation was seen in the neuropil of the septal nuclei, hippocampus, hypothalamus, thalamus, cerebellum, and globus pallidus at 100 mg/kg/day and minimal vacuolation in the thalamus, globus pallidus, and cerebellum at 30 mg/kg/day. In animals given 100 mg/kg/day for 91 days from PND22, minimal or slight vacuolation was observed only in the hippocampus, hypothalamus, and thalamus. Clear evidence of recovery was observed after the 14-day off-dose period that followed treatment from PND22 to PND35 and following the 6-week off-dose period that followed treatment from PND22 to PND112. No vigabatrin-related brain vacuolation was observed in animals given 30 or 100 mg/kg/day for 14 days from PND36.
We have not been able to find other studies where the effect of vigabatrin has been examined in juvenile dogs. However, when comparing with data from adult dogs, it appears that the juvenile animals develop the same kind of changes as the adult, that is, microvacuolation in several areas of the brain, for example, the hippocampus and thalamus. In a study by Gibson et al. (1990), the only findings following administration of up to 200 mg/kg/day vigabatrin to dogs for 12 months was vacuolation in selected myelinated tracts, which is in line with the results presented here. Also, Gibson et al. (1990) and Yarrington (1983) report no signs of neurodegeneration corresponding to our results. There does, however, appear to be age-specific differences in the dose relation of development of vacuolation. While we found vacuolation at 30 mg/kg/day in the juvenile animals only after 14 days of administration (PND22–35), no findings were apparent in dogs given 30 or 100 mg/kg/day for 14 days from PND36 or adult dogs given 50 mg/kg/day vigabatrin for 6 months. The same general finding was made in the original dog repeat-dose toxicity study by Yarrington (1983), where microvacuoles were only apparent after 1 year of treatment at 50 mg/kg/day (as was also seen by Gibson et al. 1990).
The difference in the findings between the time phases used in this study suggests that there is a specific time window during brain development in which treatment with vigabatrin is associated with vacuolation in the brain and that certain areas of the brain (septal nuclei, cerebellum, and globus pallidus) appear to show recovery from the vacuolation after this time window, even if dosing is continued. The characteristics of the findings indicate that the development of microvacuoles is related to the process of myelination, and the findings are similar to those observed in the juvenile rat (Rasmussen et al. 2015). Interestingly, recovery from microvaculoation has been reported in both rats and dogs, even during continued administration of vigabatrin. It could be argued that the observed recovery, in reality, reflects oligodendrocytes “catching up” during brain development. This is supported by the absence of cell degeneration as visualized in the Fluoro-Jade B stain. This observation is in line with previous results from studies in dogs (Megel 1979; Gibson et al. 1990; Yarrington 1983) and rats (Rasmussen et al. 2015).
In contrast to the findings in juvenile rats (Rasmussen et al. 2015), the progress of brain myelination was not affected in the dogs and there were also no histopathological effects on the oligodendrocytes (that appeared swollen in rats). Since the relationship between swollen oligodendrocytes, myelination, and microvacuolation is now well understood in rats, the absence of histopathological effects on oligodendrocytes in the dogs indicates that the juvenile dog is less sensitive than the juvenile rat with respect to both histopathology and clinical signs. As described previously, the systemic exposure to vigabatrin was closely related to dosages in both juvenile rats and juvenile dogs and was comparable between the 2 species, so a comparison may be made on a mg/kg basis: while clinical signs and even mortality were observed at 50 mg/kg/day in juvenile rats, no mortality and no clinical signs were recorded in the juvenile dogs at dosages at 100 mg/kg/day. Also, the brain vacuolation recorded in the juvenile dogs at 100 mg/kg/day was mild compared with the vacuolation (and reduced myelin) seen in the juvenile rats at 50 mg/kg/day.
In conclusion, the results presented here from juvenile dogs match previously published results from adult dogs and juvenile rats, that is, that administration of vigabatrin results in a dose- and time-dependent appearance of microvacuolation, which is reversible and appears to be linked to myelin and to the myelination process. Further, it appears that juvenile dogs (as well as rats) are more susceptible than adult animals as the findings occur at low-dose levels where no findings are made in adult animals, even following prolonged use. It is speculated that this is due to the myelination process occurring in the juvenile phase of the animals’ growth.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Author Contribution
Authors contributed to conception or design (AB, AR, DM, ADR); data acquisition, analysis, or interpretation (AB, AR, DM, ADR); drafting the manuscript (DM); and critically revising the manuscript (AB, AR, DM, ADR). All authors gave final approval and agreed to be accountable for all aspects of work in ensuring that questions relating to the accuracy or integrity of any part of the work are appropriately investigated and resolved.