
Abstract
The mechanistic relationship between increased food consumption, increased body weights, and increased incidence of tumors has been well established in 2-year rodent models. Body weight parameters such as initial body weights, terminal body weights, food consumption, and the body weight gains in grams and percentages were analyzed to determine whether such relationship exists between these parameters with the incidence of common spontaneous tumors in Tg.rasH2 mice. None of these body weight parameters had any statistically significant relationship with the incidence of common spontaneous tumors in Tg.rasH2 males, namely lung tumors, splenic hemangiosarcomas, nonsplenic hemangiosarcomas, combined incidence of all hemangiosarcomas, and Harderian gland tumors. These parameters also did not have any statistically significant relationship with the incidence of lung and Harderian gland tumors in females. However, in females, increased initial body weights did have a statistically significant relationship with the nonsplenic hemangiosarcomas, and increased terminal body weights did have a statistically significant relationship with the incidence of splenic hemangiosarcomas, nonsplenic hemangiosarcomas, and the combined incidence of all hemangiosarcomas. In addition, increased body weight gains in grams and percentages had a statistically significant relationship with the combined incidence of all hemangiosarcomas in females, but not separately with splenic and nonsplenic hemangiosarcomas.
Introduction
During the preclinical phase of the drug discovery process, the safety studies that evaluate the toxic and carcinogenic potential of the drugs are conducted in rodents. Traditionally, the 2-year studies have been conducted in rat models such as F344 and Sprague-Dawley and in mouse models such as CD-1 and B6C3F1. Over the past decade since the transgenic mouse model Tg.rasH2 became available, these studies are now also being performed for a shorter duration of 26 weeks, replacing about a quarter of all 2-year carcinogenicity studies performed in conventional mouse models (Jacobs and Hatfield 2013; Nambiar, Turnquist, and Morton 2012; Paranjpe, Elbekai, et al. 2013).
According to the American Cancer Society (2012), obesity is clearly associated with a variety of cancers in human beings, such as those involving breast, colon and rectum, uterus, esophagus, kidney, and pancreas. Overweight people are also at increased risk of developing cancers of the liver and gall bladder, non-Hodgkin lymphoma, multiple myeloma, and cancers involving the ovaries and cervix. The relationship between increased food consumption, increased body weights, hormonal perturbations, increased incidence of tumors, and increased mortality that eventually complicate the interpretation of 2-year rodent studies has been widely reported in the 2-year F344 rats, Sprague-Dawley rats, CD-1 mice, and B6C3F1 mice (Allaben et al. 1996; Biegel et al. 2001; Hart et al. 1995; Haseman et al. 1997; Haseman, Hailey, and Morris 1998; Keenan and Soper 1995; Keenan et al. 1995; Leakey, Seng, and Allaben 2003; Nohynek et al. 1993; Nunez et al. 2008; Rao 1996; Roe 1981, 1993; Seilkop 1995; Sistare et al. 2011; Turturro et al. 1996). More specifically in the F344 rat, increased body weights have been associated with increased numbers of pituitary tumors and mononuclear cell leukemias in males, and pituitary tumors and mammary gland fibroadenomas in females. In the B6C3F1 mouse, increased body weights have been associated with an increased incidence of liver tumors in both sexes (Haseman, Bourbina, and Eustis 1994; Haseman 1995a, 1995b; Haseman et al. 1997; Haseman, Hailey, and Morris 1998; Kari and Abdo 1995; Rao, Piegorsch, and Haseman 1987; Thurman et al. 1994; Witt, Sheldon, and Thurman 1991).
In our laboratory, we performed twenty-six, 26-week short-term carcinogenicity studies using the Tg.rasH2 mouse model. The data pertaining to spontaneous tumors in these mice have been published (Paranjpe, Elbekai, et al. 2013). However, relationship of body weight parameters with the incidence of tumors in the Tg.rasH2 mice, if any, has not been published so far. The purpose of this publication is to determine whether any relationship exists between body weight parameters such as initial body weights, terminal body weights, food consumption, and the body weight gains in grams and percentages with the incidence of spontaneous tumors in this mouse model. Specifically, we evaluated the common spontaneous tumors in this strain of mouse (Nambiar, Turnquist, and Morton 2012; Paranjpe, Elbekai, et al. 2013), namely lung tumors (adenomas and carcinomas), splenic hemangiosarcomas, nonsplenic hemangiosarcomas, the combined incidence of all splenic and nonsplenic hemangiosarcomas, and Harderian gland tumors (adenomas and carcinomas).
Materials and Methods
Animals
CByB6F1-Tg(HRAS)2Jic (+/− hemizygous c-Ha-ras) mice, obtained from Taconic Farms (Germantown, NY), were used in all studies. The knock-in Tg element (human prototype c-Ha-ras gene with its own promoter/enhancer) is injected into C57BL/6 × BALB/c F2 zygotes, which are crossed back to C57BL/6J forming C57BL/6JJic-Tg(HRAS)2Jic. The CByB6F1-Tg(HRAS)2Jic (+/− hemizygous c-Ha-ras) is the offspring from a cross of the C57BL/6JJic-Tg(HRAS)2Jic hemizygous male mice with the BALB/cByJJic female mice. Each mouse was genotyped by Taconic to verify the presence of the transgene before being placed on study. Animals were assigned to groups using a computer-generated randomization program. On the first day of treatment, animals were 6 to 10 weeks of age and weighed at least 20 g or 15 g (males and females, respectively). Individual body weights for each dose group of each sex were within ±20% of the mean at the start of the study. Even though the starting age has varied between 6 and 10 weeks between studies, every attempt was made to start any given study around starting age of 8 weeks. However, the starting age of the mice on the study depended on the breeding schedules of the supplier, Taconic. Nevertheless, in any given study, the starting age of males and females was similar.
Housing and Environmental Conditions
Housing and environmental conditions were similar in all studies. Animals were single housed in polycarbonate cages with hardwood bedding chips in environmentally controlled rooms. Animals were verified to be free of illness prior to being placed on a study. All animals had ad libitum access to water and powdered feed (Harlan TEKLAD Global Diet, Madison, WI). While the lot numbers and the milling dates of the feed varied between the studies, in general, all batches of feed contained about 18.5% of protein, 6% of fat, and 3.5% of fiber. Each batch of feed was analyzed by the manufacturer for concentrations of specified heavy metals, aflatoxins, chlorinated hydrocarbons, organophosphates, and specified nutrients. The results of these analyses are on file at BioReliance. There were no contaminants in any of the batches of feed at levels that would interfere with the study.
Regulatory Requirements
The numbers of animals, procedures, and experimental design for each study were reviewed and approved by the BioReliance Institutional Animal Care and Use Committee (IACUC). All procedures followed the specifications recommended in The Guide for the Care and Use of Laboratory Animals and were conducted in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-accredited facility. All procedures involving but not limited to quarantine and acclimation, randomization, application of unique identification system, housing, provision of food and water, administration of test article, recording of clinical signs, necropsy, and tissue processing were followed in strict accordance with the Good Laboratory Practice Regulations, Standard Operating Procedures, and protocol for each study.
Retrospective Analysis
The database was constructed based on Tg.rasH2 mice assigned to 26 studies conducted at our facility, following the same study design. The first of these studies was completed in 2004 and the last study was completed in 2012. Vehicle was administered to 710 male and 710 female mice by oral gavage (21 studies), dosed feed (3 studies), or intravenous injection (2 studies) for 26 weeks. Predose body weights were collected on day 1, and mice were weighed weekly thereafter for the next consecutive 13 weeks. After 13 weeks, the body weights were collected biweekly until the termination of the study at 26 weeks. Food consumption was recorded weekly for individual animals from day 1 until the termination of the study at 26 weeks. Upon the death or moribund sacrifice of an animal and at the termination of the study, all animals in each study were humanely sacrificed by CO2 overdose and subjected to complete necropsy. Protocol required tissues were saved in 10% neutral buffered formalin, processed routinely, stained with hematoxylin and eosin, and evaluated microscopically. Each study was peer reviewed.
Common spontaneous tumors noted in Tg.rasH2 male and female mice were grouped as follows: (1) lung tumors (adenomas and carcinomas), (2) splenic hemangiosarcomas, (3) nonsplenic hemangiosarcomas, (4) combined incidence of all hemangiosarcomas (splenic and nonsplenic), and (5) Harderian gland tumors (adenomas and carcinomas).
For each tumor category listed previously, the summary statistics for the group with tumors versus the group without tumors were calculated by gender. The initial body weights, the terminal body weights, food consumption, and the body weight gains in grams and percentages between the animals with or without tumor were compared by the nonparametric Wilcoxon rank sum test (Hollander and Wolfe 1973). A two-sided version of the test was used, and the p value indicating the significance of the test is presented in the summary tables. Analysis for each body weight parameter was performed at 2 time points, half way through the study at 13 weeks and at terminal sacrifice at 26 weeks. In addition, the average initial body weights and total tumor counts for each tumor category for each sex were used to calculate the Pearson correlation coefficient to rule out if the higher initial body weights due to higher starting age had any influence on the incidence of tumors in either sex (Rodgers and Nicewander 1988). All calculations were performed using SAS Version 9.2. Throughout this article, the average initial body weights, average terminal body weights, average body weight gains in grams, and average daily food consumption are expressed as M ± SD.
Results
The information pertaining to the mortality/survival, body weight parameters, and the incidence of common spontaneous tumors in percentages is provided in Table 1. The mortality in both male and female mice was identical at 3.94%. The average initial body weights, average terminal body weights, and the body weight gains in grams of male mice, expressed as M ± SD were 23.41 ± 0.86, 29.39 ± 2.20, and 5.98 ± 2.08, respectively. The average initial body weights, average terminal body weights, and the body weight gains in grams of female mice were 18.74 ± 0.60, 23.56 ± 0.98, and 4.82 ± 1.00, respectively. The percentage body weight gain in male and female mice was 25.62% and 25.79%, respectively. The average daily food consumption in grams in male and female mice was 3.47 ± 0.25 and 3.79 ± 0.72, respectively.
Mortality/survival, body weight parameters, and incidence of tumors.

In 710 male mice, there were a total of 164 male mice with tumors, whereas in 710 female mice there were a total of 167 female mice with tumors. The body weight parameters were compared in animals with or without the common spontaneous tumors. The common spontaneous tumors in male mice included 87 lung tumors (53%), 26 splenic hemangiosarcomas (16%), 13 nonsplenic hemangiosarcomas (7%), 39 all splenic and nonsplenic hemangiosarcomas (23%), and 11 Harderian gland tumors (7%). The common spontaneous tumors in female mice included 55 lung tumors (33%), 26 splenic hemangiosarcomas (16%), 24 nonsplenic hemangiosarcomas (14%), 50 all splenic and nonsplenic hemangiosarcomas (30%), and 25 Harderian gland tumors (15%).
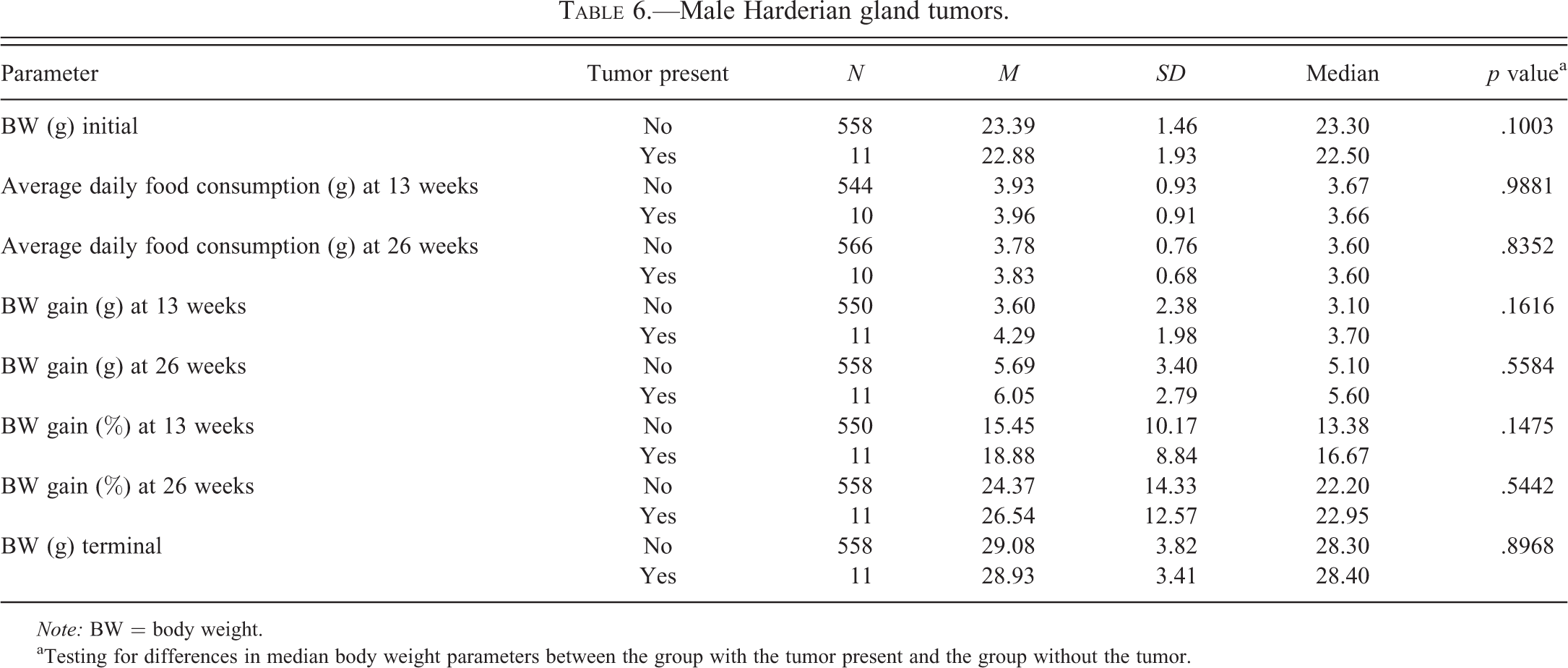
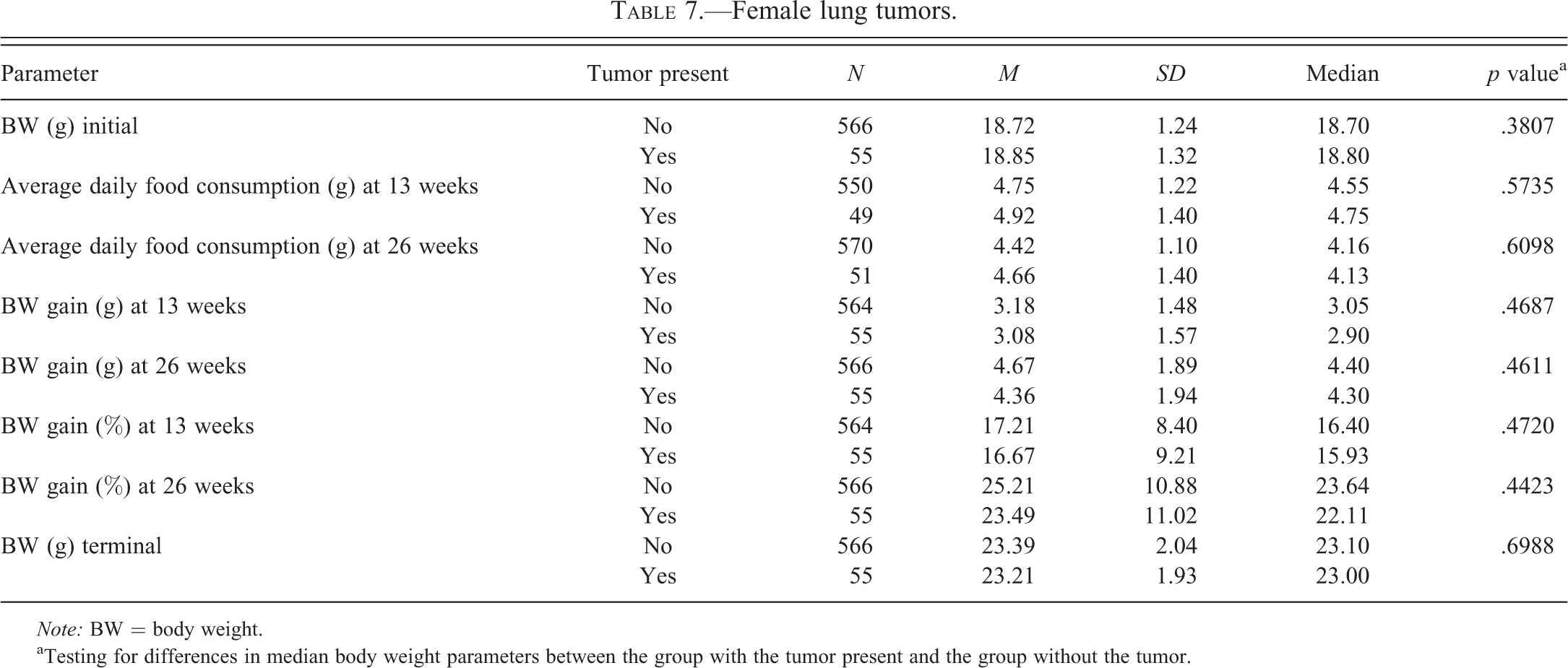
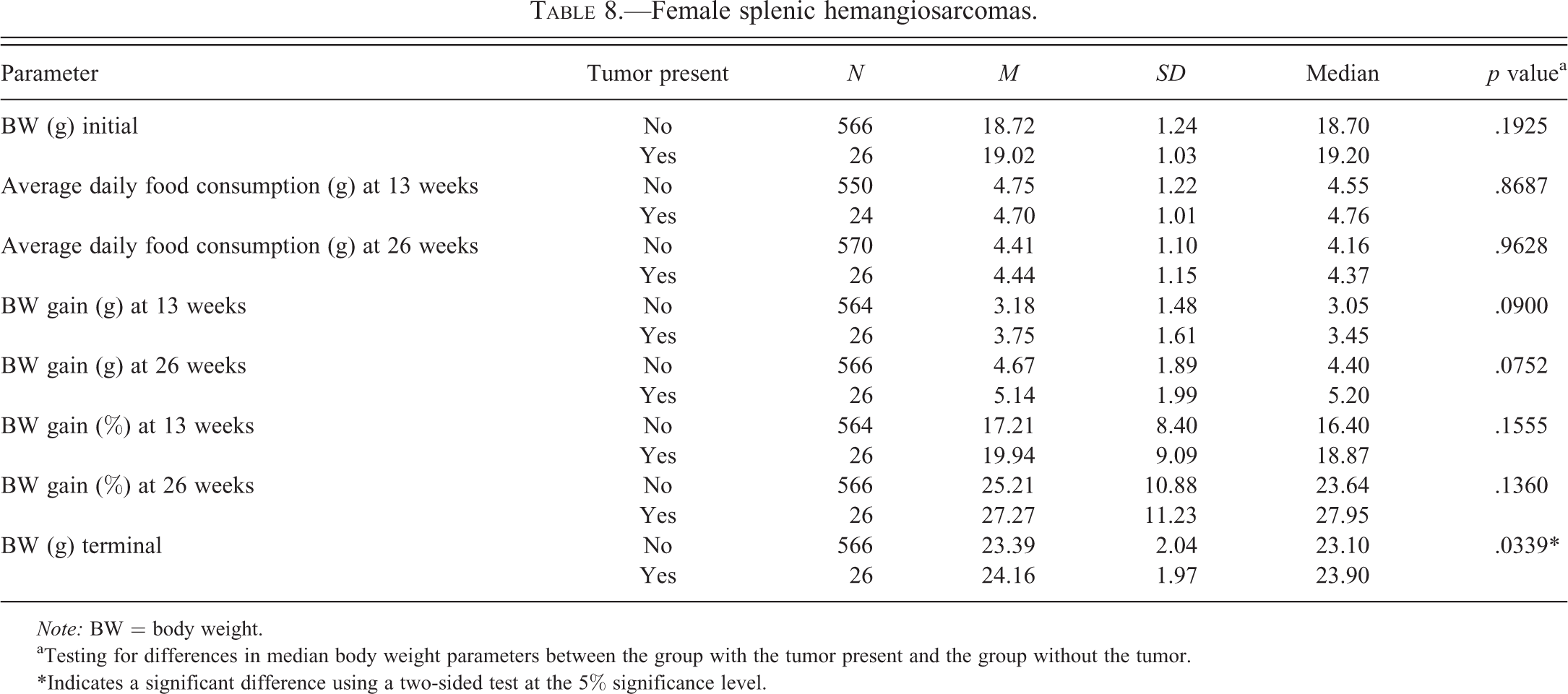
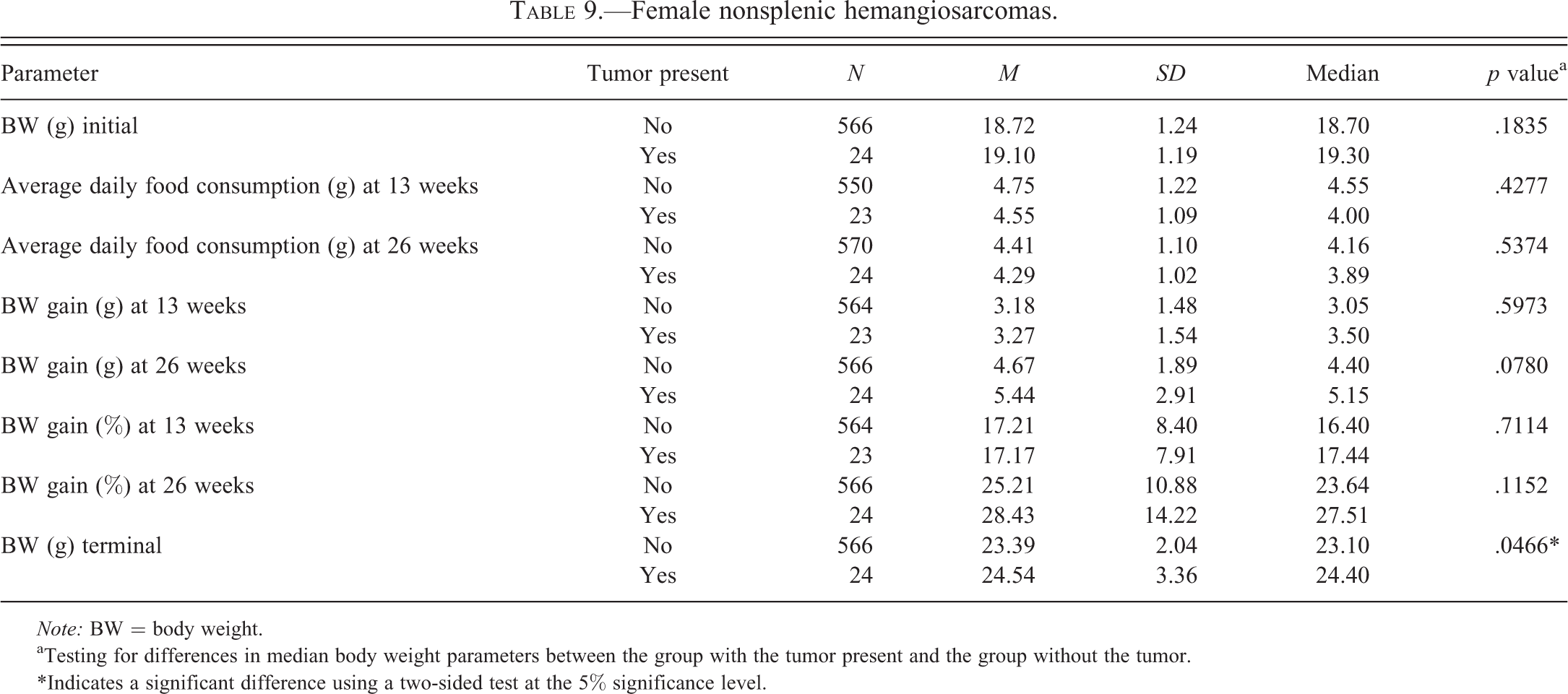
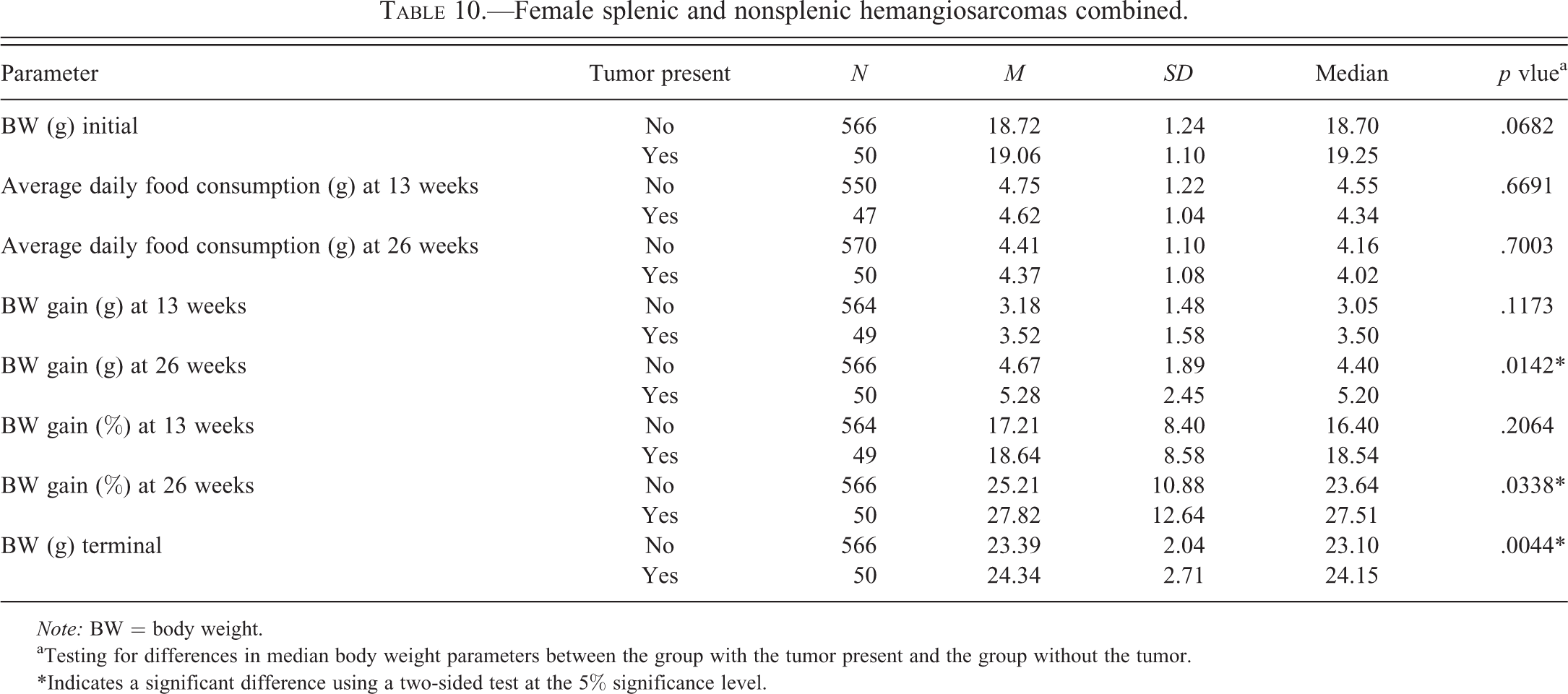
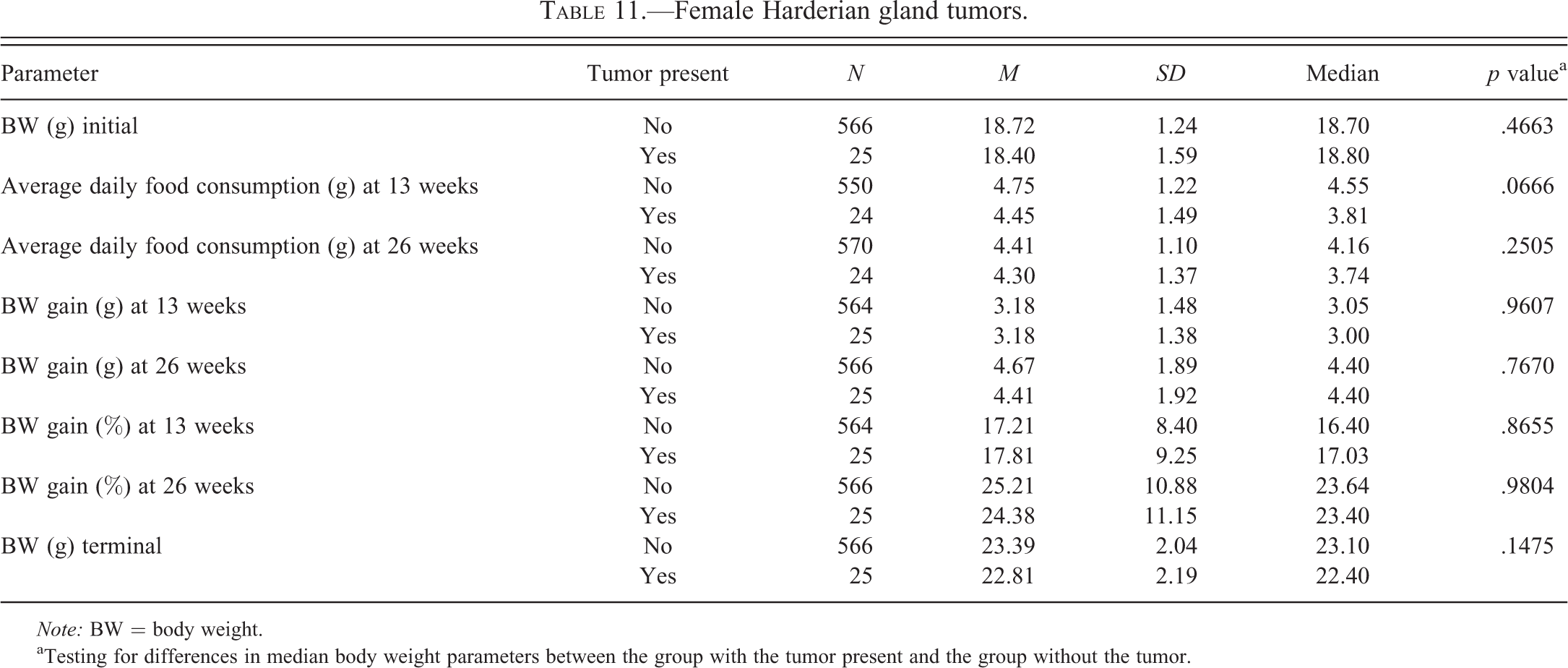
Tables 2 to 6 for males and Tables 7 to 11 for females illustrate the relationship of body weight parameters, such as initial body weights, terminal body weights, food consumption, and body weight gains in grams and percentages with the incidence of common spontaneous tumors at 2 time points, 13 weeks and 26 weeks. There were no statistically significant differences and therefore no relationship with any of the body weight parameters for all tumor types in male mice at both time points, 13 weeks and 26 weeks. In the female mice, there were no statistically significant differences and therefore no relationship with any of the body weight parameters for all tumor types at the 13 weeks time point. At 26 weeks in the female mice, there were no statistically significant differences and therefore no relationship with any of the body weight parameters for lung and Harderian gland tumors. However, in female mice at the 26 weeks time point, increased terminal body weights did have a statistically significant relationship with the incidence of splenic hemangiosarcomas, nonsplenic hemangiosarcomas, and the combined incidence of all hemangiosarcomas. In addition, in female mice, at the 26 weeks time point, the body weight gain in grams and percentages had a statistically significant relationship with the combined incidence of all hemangiosarcomas, but not with the splenic and nonsplenic hemangiosarcomas separately.
Male lung tumors.
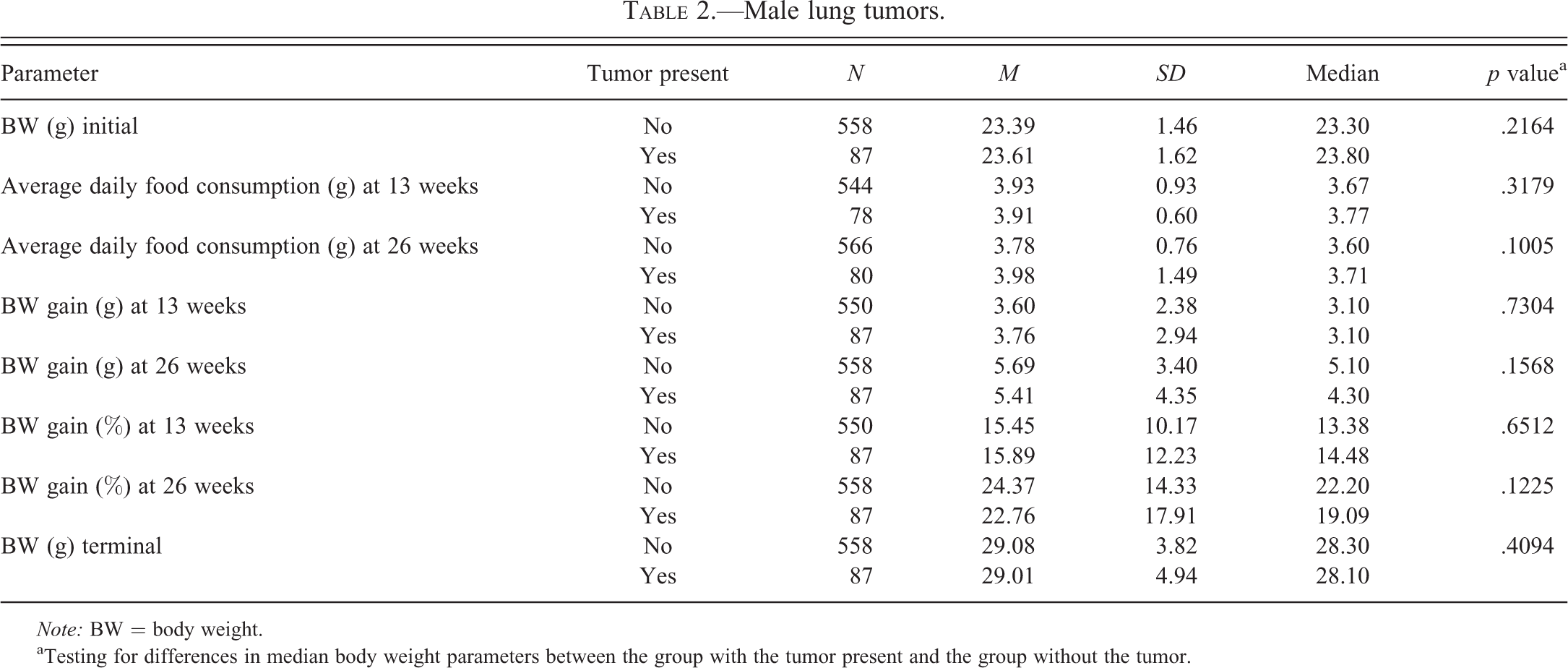
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
Male splenic hemangiosarcomas.
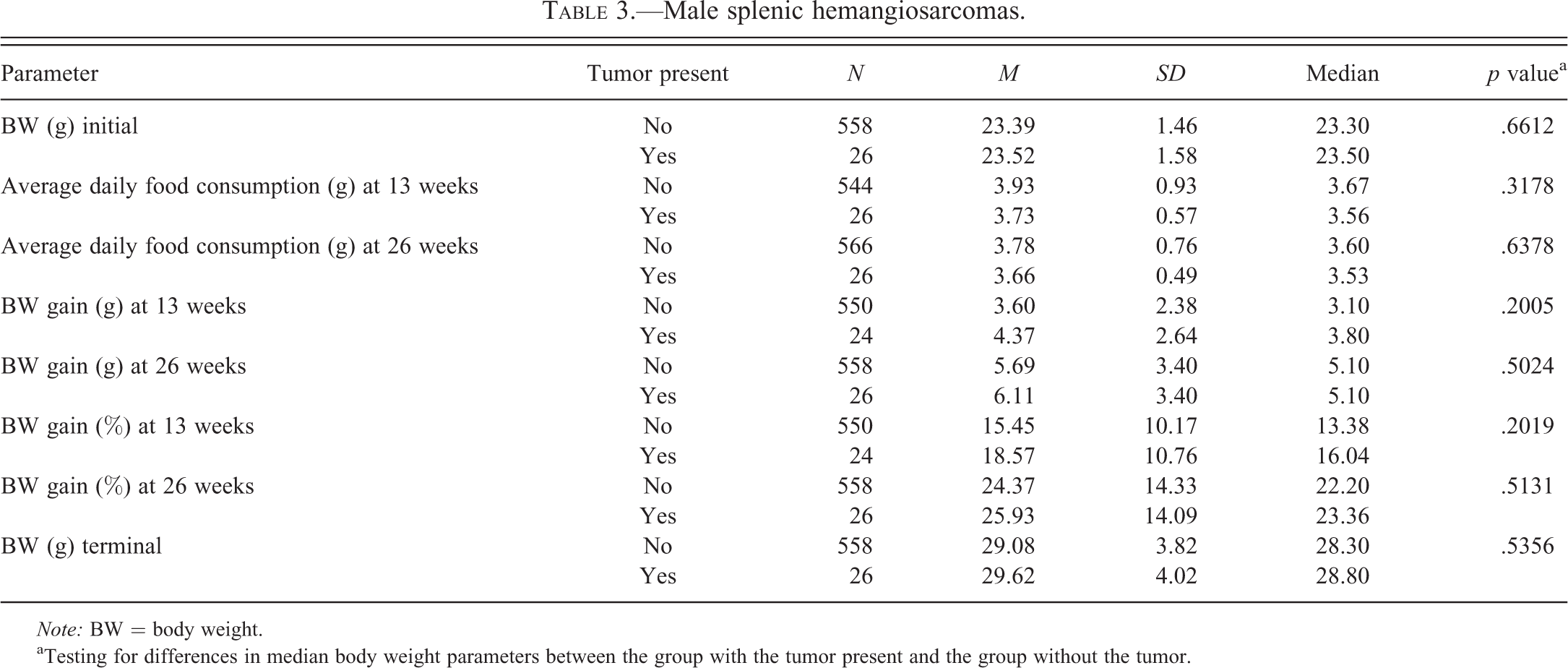
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
Male nonsplenic hemangiosarcomas.
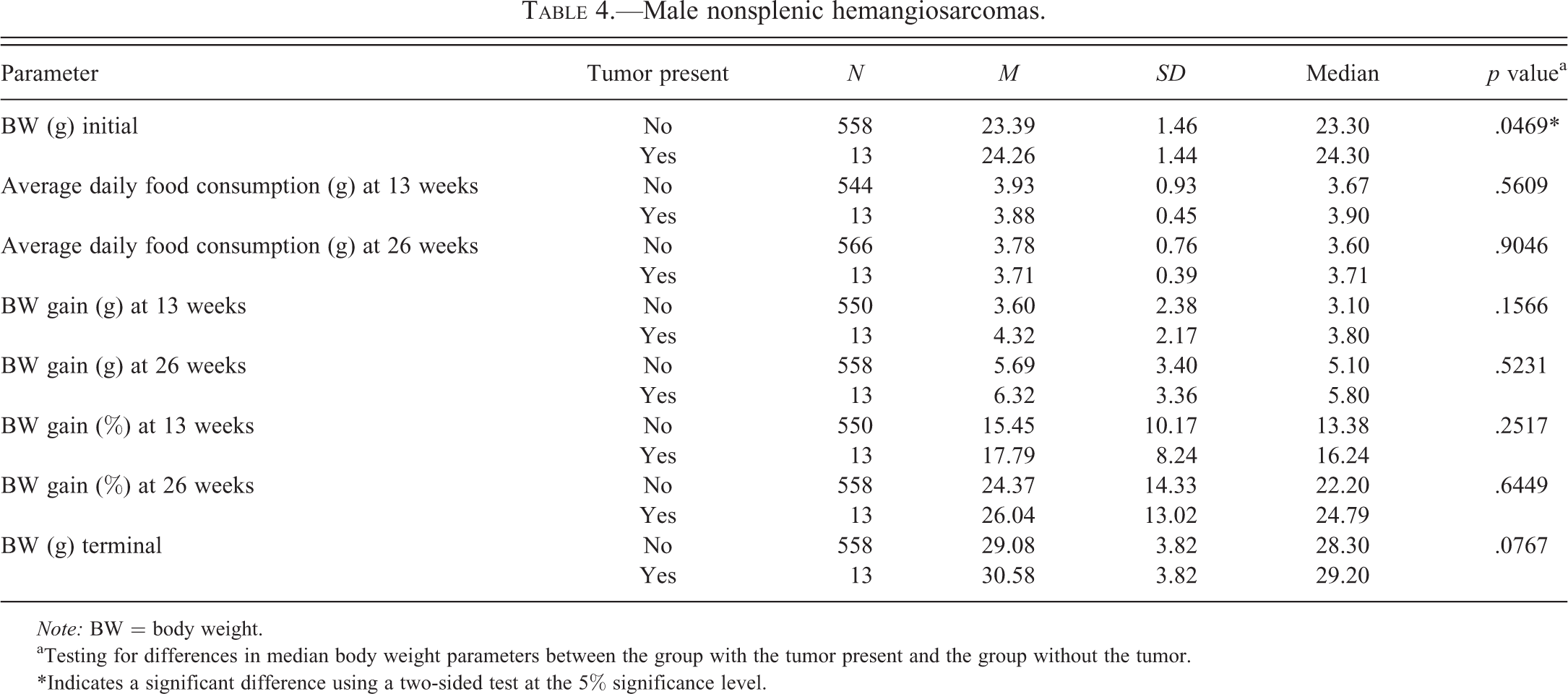
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
*Indicates a significant difference using a two-sided test at the 5% significance level.
Male splenic and nonsplenic hemangiosarcomas combined.
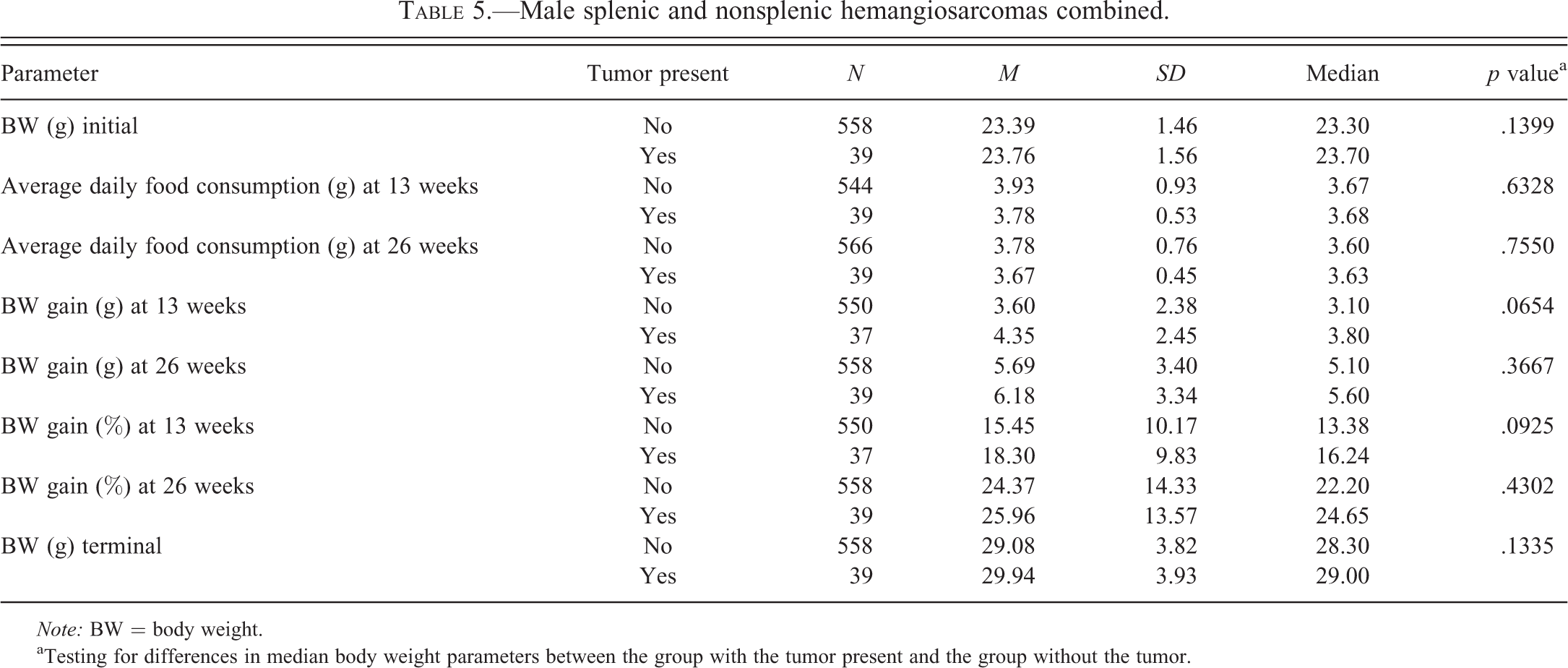
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
Male Harderian gland tumors.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
For males and females, the average initial body weights and total tumor counts for each tumor category were analyzed to determine the correlation of coefficient. The results of this analysis are presented in Table 12. These results indicated that in male mice there was no relationship between the total number of tumors for each tumor category with higher initial body weights and thereby possibly higher starting ages. In the female mice, there was also no such relationship between lung tumors, Harderian gland tumors, and splenic hemangiosarcomas. However, in the female mice, there was a statistically significant relation between the nonsplenic hemangiosarcomas and combined incidence of all hemangiosarcomas with the higher initial body weights.
Female lung tumors.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
Female splenic hemangiosarcomas.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
*Indicates a significant difference using a two-sided test at the 5% significance level.
Female nonsplenic hemangiosarcomas.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
*Indicates a significant difference using a two-sided test at the 5% significance level.
Female splenic and nonsplenic hemangiosarcomas combined.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
*Indicates a significant difference using a two-sided test at the 5% significance level.
Female Harderian gland tumors.
Note: BW = body weight.
aTesting for differences in median body weight parameters between the group with the tumor present and the group without the tumor.
Relationship between initial body weights and tumors.

*Indicates statistical significance.
Discussion
The average initial body weight of the Tg.rasH2 male mice in our studies expressed as M ± SD was 23.41 ± 0.86 g and that of female mice was 18.74 ± 0.60 g, when the mice were placed on the study between 6 and 10 weeks of age. The body weights of the Tg.rasH2 mice are 80 to 90% of their wild-type counterparts, namely C57BL/6 and the BALB/c mice (Taconic information 2013; Yamamoto et al. 1997). The initial body weights of CD1 and B6C3F1 mice when placed on study at 6 to 8 weeks of age are about 20 to 25% more than the Tg.rasH2 mice (Charles River information 2013; Harlan information 2013). Thus, in general, the Tg.rasH2 mice are smaller than mouse strains commonly used in conventional 2-year studies. The initial body weights are tied to the age of the mice at the start of the study. At the start of the study, these mice are 6 to 10 weeks old and by the completion of the study these mice would be 32 to 36 weeks old. It is important to start the study in the range of 6 to 10 weeks of age, as at around 40 weeks of age the mortality in these mice substantially increases, more so in females than in males, due to increased development of spontaneous tumors (Yamamoto et al. 1998).
The body weight parameters did not have any statistically significant relationship with any of the tumor types in both sexes of mice at 13 weeks. This suggests that at least at 13 weeks these body weight parameters are less likely to have any influence on future development of these tumors noted at 26 weeks. In male mice, these body weight parameters did not have any statistically significant relationship with any of the tumor types at 26 weeks. At the 26 weeks time point, the female mice did not have any statistically significant relationship with lung and Harderian gland tumors. However, in female mice at the 26 weeks time point, increased terminal body weights did have a statistically significant relationship with the incidence of splenic hemangiosarcomas, nonsplenic hemangiosarcomas, and the combined incidence of all hemangiosarcomas. In addition, at the 26 weeks time point in female mice, increased body weight gains in grams and percentages had statistically significant relationship with the combined incidence of all hemangiosarcomas, but not with the splenic and nonsplenic hemangiosarcomas separately. At our facility, although the starting age of the mice between studies has varied between 6 and 10 weeks, every attempt was made to initiate the study at the starting age of 8 weeks. However, the starting age depended on the breeding schedules of the supplier, Taconic. Nevertheless, in any given study, the starting age of the males and females was similar. The data analyzed to determine the coefficient of correlation showed that there was no positive correlation between the number of tumors for each tumor category with higher initial body weights and thereby higher starting ages in male mice. This analysis also showed that there was no positive relationship between the lung tumors, Harderian gland tumors, and splenic hemangiosarcomas in female mice. However, there was a statistically significant relationship between the higher initial body weights and nonsplenic hemangiosarcomas as well as combined incidence of all hemangiosarcomas in female mice. Clearly, the positive relationship between higher initial body weights and combined incidence of all hemangiosarcomas in female mice is due to nonsplenic hemangiosarcomas, as the splenic hemangiosarcomas did not demonstrate this relationship.
The cause for having a relationship with the splenic and nonsplenic hemangiosarcomas in female mice only but not in male mice is not clear. However, it needs to be pointed out that in terms of body weight gain in percentages, the female mice (25.79%) showed only slight gain compared to the male mice (25.62%). Also the average daily food consumption expressed as M ± SD in grams in female mice (3.79 ± 0.72) was more than the male mice (3.47 ± 0.25). These differences may be important because despite the initial body weights of female mice being 4 to 5 g lower than the males, both the food consumption and the body weight gain in percentages were higher in female mice. Given the fact that the incidence of splenic hemangiosarcomas was identical in both sexes at 16%, it is not likely that these splenic hemangiosarcomas would have any influence on terminal body weights and weight gain in percentage in female mice alone and not in male mice. The incidence of nonsplenic hemangiosarcomas was double in female mice (14%) compared to the male mice (7%). The majority of the nonsplenic hemangiosarcomas involved the reproductive system in both male and female mice; however, the hemangiosarcomas involving the reproductive system of female (uterus, vagina, and ovary) were double in number (Paranjpe, Elbekai, et al. 2013) compared to the male reproductive organs (testes, epididymides, and seminal vesicles). Also the nonsplenic hemangiosarcomas involving the skin/subcutis were much higher in number in female mice as compared to the male mice (Paranjpe, Elbekai, et al. 2013). We have consistently observed enlargement of spleens and therefore increase in their weights due to extramedullary hematopoiesis in mice of both sexes whenever nonsplenic hemangiosarcomas were present in the same animals (Paranjpe, Shah, et al. 2013). Because the incidence of nonsplenic hemangiosarcomas is double in female mice than the male mice, the number of female mice with splenic enlargement and therefore increased splenic weights due to extramedullary hematopoiesis are also expected to be double in number compared to the male mice. Based on all these factors, it is likely that the association noted between the increased terminal body weights and body weight gain with the hemangiosarcomas in female mice only but not in male mice may have been due to a combination of factors such as more average daily food consumption, more body weight gain in percentages, increased incidence of nonsplenic hemangiosarcomas involving the reproductive system and skin/subcutis, and secondary splenic enlargement of the spleen due to extramedullary hematopoiesis. In addition, the association between the increased initial body weights and nonsplenic hemangiosarcomas in female mice is noteworthy.
The incidence of lung tumors in male mice (53%) was considerably higher than in female mice (33%); whereas the incidence of Harderian gland tumors in male mice (7%) was much lower than in the female mice (15%). However, neither lung nor Harderian gland tumors demonstrated any statistically significant relationship with any of the body weight parameters in both sexes.
The mechanistic relationship between increased food consumption, increased body weights, hormonal perturbations, increased incidence of tumors, and increased mortality that eventually complicate the interpretation of 2-year rodent studies has been widely reported for the 2-year F344 rats, Sprague-Dawley rats, CD-1 mice, and B6C3F1 mice (Allaben et al. 1996; Biegel et al. 2001; Hart et al. 1995; Haseman et al. 1997; Haseman, Hailey, and Morris 1998; Keenan and Soper 1995; Keenan et al. 1995; Leakey, Seng, and Allaben 2003; Nohynek et al. 1993; Nunez et al. 2008; Rao 1996; Roe 1981, 1993; Seilkop 1995; Sistare et al. 2011; Turturro et al. 1996). Rather than mechanisms that involve alterations in body weight parameters and hormonal perturbations in 2-year rodent models, the mechanism of tumor production in Tg.rasH2 mouse involves overexpression of C-Ha-ras gene product, p21 (Maruyama et al. 2001; Saitoh et al. 1990; Sekiya et al. 1985; Tamaoli 2001; Usui et al. 2001; Yamamoto et al. 1998). These mechanistic differences also explain the differences in the types of spontaneous tumors in 2-year rodents and Tg.rasH2 mice. To name a few, the common tumors in 2-year rodents include tumors of pituitary, thyroid, testes, liver, and mammary glands; all of which are influenced by the combination of mechanisms such as increased food consumption, increased body weights, and hormonal perturbations (Allaben et al. 1996; Biegel et al. 2001; Hart et al. 1995; Haseman, Bourbina, and Eustis 1994; Haseman 1995a, 1995b ; Haseman et al. 1997; Haseman, Hailey, and Morris 1998; Kari and Abdo 1995; Keenan and Soper 1995; Keenan et al. 1995; Leakey, Seng, and Allaben 2003; Nohynek et al. 1993; Nunez et al. 2008; Rao 1996; Rao, Piegorsch, and Haseman 1987; Roe 1981, 1993; Seilkop 1995; Sistare et al. 2011; Thurman et al. 1994; Turturro et al. 1996; Witt, Sheldon, and Thurman 1991). In contrast, the common tumors in Tg.rasH2 include lung tumors, hemangiosarcomas, and Harderian gland tumors (Nambiar, Turnquist, and Morton 2012; Paranjpe, Elbekai, et al. 2013), none of which are likely to be influenced by hormonal perturbations. Among all of the nonsplenic hemangiosarcomas, the incidence of hemangiosarcomas involving the male and female reproductive system was higher (Paranjpe, Elbekai, et al. 2013); however, these hemangiosarcomas of the reproductive system in Tg.rasH2 mice are not considered to be hormonally induced.
In summary, at both 13 weeks and 26 weeks, the body weight parameters did not have any statistically significant relationship with the spontaneous tumors in the Tg.rasH2 male mice, namely lung and Harderian gland tumors and splenic and nonsplenic hemangiosarcomas. These parameters also did not have any significant relationship with any of the tumor types in female mice at the 13 weeks time point. At the 26 weeks time point, these parameters also did not have any significant relationship with lung and Harderian gland tumors in female mice. However, at the 26 weeks time point, these parameters did have a statistically significant relationship with splenic hemangiosarcomas, nonsplenic hemangiosarcomas, and combination of all hemangiosarcomas in female mice. The causes of this relationship in the female mice alone may be multifold and may include a combination of factors such as: Increased daily food consumption in female mice compared to male mice Increased body weight gains in percentage in female mice compared to the male mice Increased incidence of nonsplenic hemangiosarcomas in female mice compared to the male mice, particularly those involving the female reproductive system and skin/subcutis The tumor, hemangiosarcoma, itself contributing to the higher terminal body weights in tumor-bearing animals Increased initial body weights that may have contributed to the increased incidence of nonsplenic hemangiosarcomas in female mice Secondary extramedullary hematopoiesis that causes enlargement of spleens in mice with nonsplenic hemangiosarcomas.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.