
Abstract
Histopathological examination of the nasal passages requires a standardized approach for recording lesion distribution patterns. Nasal diagrams provide guidance to map the lesions. Information on lesions exists for rodents, dogs, and monkeys, which all have been used in inhalation studies. Recently, minipigs have garnered interest as an inhalation model because minipigs resemble humans in many features of anatomy, physiology, and biochemistry and may be a good alternative to monkeys and dogs. The present work explored the microanatomy and histology of the nasal passages of Göttingen minipigs from postnatal day 1 until 6 months of age. Six nasal levels were selected, which allow examination of the squamous, transitional (nonciliated) and ciliated respiratory, and olfactory epithelia; the nasopharynx; and relevant structures such as the vomeronasal organ, olfactory bulb, and nasal/nasopharynx-associated lymphoid tissue.
The upper respiratory tract, especially the nasal passages, is an important target site for inhaled xenobiotics and drugs (Haschek, Witschi, and Nikula 2002). The mucous membranes of the nasal passages are the first tissues exposed to inhaled substances, and detailed information of the anatomy and histology of these tissues is needed. Minipigs and humans have 2 nasal passages that are essentially separate, a feature not seen in other species commonly used in safety assessment. To a certain extent, each animal could thus serve as its own control by exposing only 1 nostril (Bode et al. 2010). Like humans, pigs can breathe through the nose as well as the mouth, although they breathe mainly through the nose (habitual nose breathers; Martinez 2010). The size of the nose allows the use of the same device and dose as intended in humans. Permeation and absorption of substances through minipig nasal tissues demonstrated a good correlation with these processes in man (Rankløve et al. 2006). Some structural differences are found as well between minipigs and humans, such as the size of the vestibule, which will affect the microanatomy and histology of the nasal passages. Pigs in general have excellent smell, and therefore minipigs are expected to have a more extensive area of main olfactory epithelium than the vision-oriented humans.
Until now, minipigs have been used only sparingly for inhalation studies (Franco-Cereceda et al. 1995; Sparrow 1996; Koch et al. 2001; Nunoya et al. 2007; Windt et al. 2010). Studies on nasal tissues have been even rarer, and the histologic examination in these few cases was limited to a few samples of the mucosa (Marini et al. 1998; Rankløve et al. 2006). Careful mapping of induced nasal lesions in frequently used animal species such as mice, rats, dogs, and monkeys has been very helpful to establish the mode of action and facilitate extrapolation of the findings to humans (Young 1981; Woutersen et al. 1987; Boorman, Morgan, and Uriah 1990; Morgan and Monticello 1990; Harkema 1991; Mery et al. 1994; Herbert and Leininger 1999; Harkema, Carey, and Wagner 2006; Carey et al. 2007; Craven et al. 2007). At the same time, the lesion distribution patterns have provided the basis to recommend the proper number and location of levels to be examined routinely in guideline-driven safety studies, for example, according to OECD412 and OECD413 regulations for inhalation toxicity. Because deposition patterns of inhaled substances are largely unknown for minipigs, information from other species should guide selection of nasal levels.
The purpose of the present work was to describe the normal microanatomy of the nasal passages of the Ellegaard Göttingen Minipig and to provide guidance for a standard method to sample the minipig nasal passages. The interest in juvenile safety testing is rapidly growing; therefore, the development of the nasal passage and its associated structures were studied in minipigs of different ages, from day 1 up to 6 months of age, which represents the age of sexual maturation.
Materials and Methods
Animals
Male Göttingen Minipigs (Ellegaard, Göttingen Minipigs, Denmark) were used, 1 day to 6 months of age (Table 1). The pigs younger than 5 weeks were born at TNO from pregnant sows, shipped from Ellegaard. Juvenile to young adult minipigs (approximately 2, 3, and 6 months of age) were obtained directly from the animal provider. All procedures and handling were in accordance with the institutional, governmental, and European guiding principles in the use and care of animals.
Age and developmental stage of the minipigs examined.

a According to Beck et al. (2005).
Animal Husbandry
The animals were kept in indoor pens, according to European guidelines for housing of laboratory minipigs. The room was ventilated with approximately 10 air changes per hour and was targeted at a temperature of 20°C to 24°C with a relative humidity of 40% to 70%. Lighting was artificial with a sequence of 12 hr light and 12 hr dark. The animals were fed measured amounts of a commercial minipig diet (SMP (E) SQC; SDS Special Diets Services, Witham, England). Piglets were offered a solid piglet diet, 30 g twice a day per piglet, from PN day 14 until weaning (piglet diet delivered by Ellegaard). The 6-week-old piglets were fed a combination of piglet diet and SDS minipig diet. Feed and apples in limited amounts were also used to socialize the animals. Tap water suitable for human consumption (quality guidelines according to Dutch legislation based on EC Council Directive 98/83/EC) was provided ad libitum. Piglets were born after a gestation period of 113 days, which is normal for minipigs.
Necropsy and Histology
Prior to necropsy, the animals were sedated by Domitor (Pfizer Orionpharma, medetomidine hydrochloride, 1 mg/ml) 0.5 ml/10 kg body weight i.m. + Dormicum (Roche, midazolam, 5 mg/ml) 0.5 ml/10 kg body weight i.m. and anesthetized with Na-pentobarbital (60 mg/ml, ca 1 ml/kg body weight, intracardially) until deep anesthesia and euthanized by exsanguination via the abdominal aorta (piglets until the age of day 7) or via the axillary artery (older animals). The head was collected and the under jaw was removed. The nasal passages were flushed with a 10% neutral phosphate-buffered formalin through the buccal opening of the nasopharyngeal duct and thereafter immersed in formalin. Subsequently, the nose was cut into slices of about 0.4 cm with a band saw (Exact B10 CP Macroband manufactured by Exact, Germany, and supplied by Klinipath, the Netherlands), using the landmarks (mainly dental landmarks and in addition the rostral and caudal ends of the palate) provided in the diagram (Figure 1; gross structures per level, see Figure 2). The level with nasal/nasopharynx-associated lymphoid tissue (nasopharyngeal tonsil; NALT; level 6/J) was sampled differently: first, a cut was made through the buccal opening of the nasopharynx, which was parallel to the cuts of the other levels; NALT was then visible in the caudal part of the nasopharynx, and a slice was taken through NALT. The nose of the 3-day-old and of the 4-, 8-, and 13-week-old animals was cut completely, leading to 10 to 16 levels (more levels in older animals; Figure 1: levels in letters for the 3-day-old animals). Based on these cross sections, a selection of 8 levels was taken from the nasal passages of the other animals. From these, 6 levels were selected for recommendation, leaving out the level anterior to level 1 and the level with the palatine tonsil (Figure 1: levels in numbers). After fixation of 2 to 3 days, the slices were decalcified in 10% formic acid, with the exception of the nose of the very young animals, which did not need decalcification. The slices were decalcified for 3 days, during which the solution was changed twice. Before processing, the slices of the older animals needed to be trimmed, and those from the central part of the nose (ethmoturbinates and olfactory bulb) were cut in half or into more pieces in order to fit the cassettes. The slice at level 5 could be embedded in total (Figures 2 and 4). The tissues were embedded in paraffin wax, sectioned at 5 µm, stained with hematoxylin and eosin, and examined microscopically.
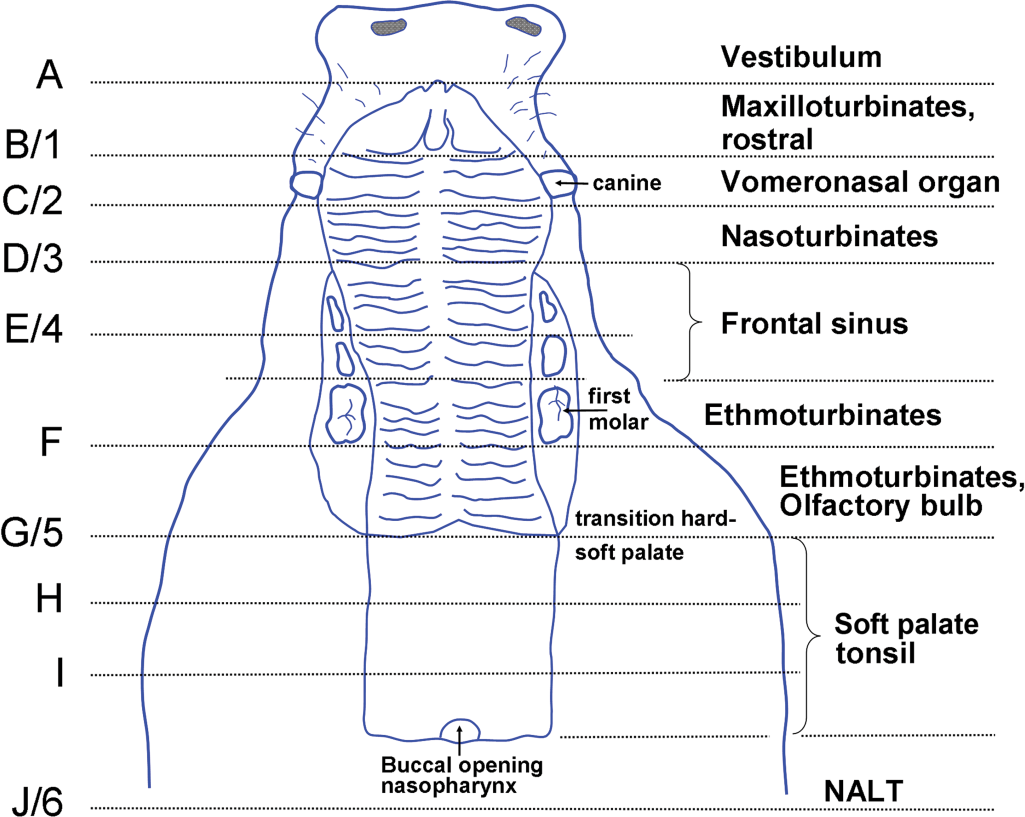
Major landmarks on the dorsal buccal cavity of the minipig for cross/transverse section level identification. The teeth are not depicted, except for the canine teeth. Scheme according to the anatomy observed in a 6-month-old animal, with the first molar present. At 6 months of age, the temporary canine teeth are less conspicuous, as shown in this scheme. Recommended level 1 is before the canine teeth, level 2 behind these teeth, level 3 before the first premolar, level 4 behind the premolar, level 5 at the transition between the hard and the soft palate, and level 6 at the caudal side of the slice taken from the buccal opening of the nasopharynx to more caudally.
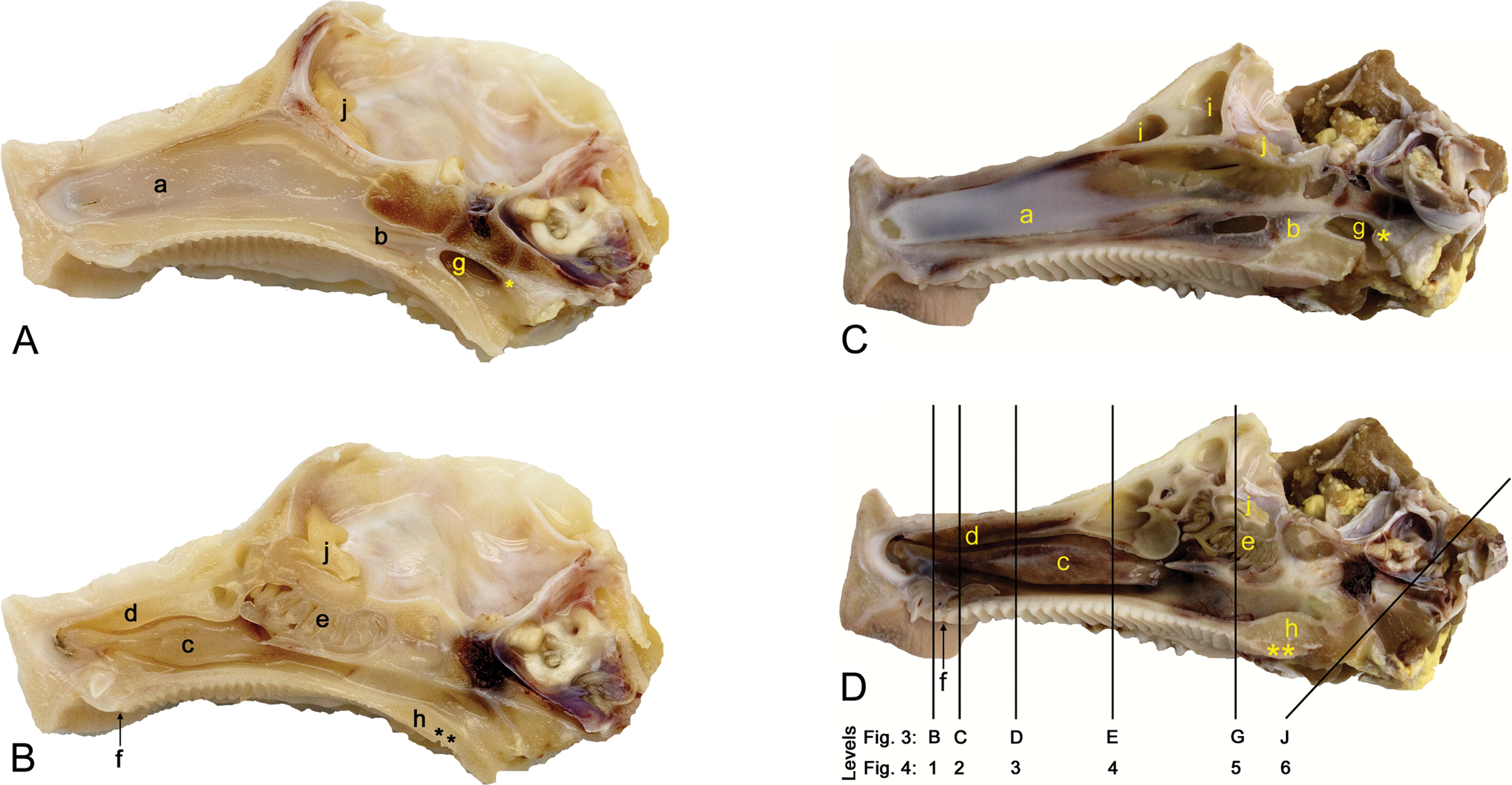
(A-D) Midsaggital section of the nasal passages of a minipig aged 3 days (A, B) and 6 months (C, D) before (A, C) and after (B, D) removal of the septum. The actual size of the 6-month-old animal was about 2.5 times greater than the 3-day-old animal (the entire length of the specimen—from nostril to the right end—was 7 cm for the 3-day-old animal and 18 cm for the 6-month-old animal). Shown here are the nasal septum (a) and the nasopharyngeal septum (b; the oval window in C is an artifact) and the area of NALT (*), the maxilloturbinates (c), the nasoturbinates (d), the ethmoturbinates (e), the nasopalatine duct (f), the nasopharynx (g), the soft palate (h) with the area of the soft palate tonsil (**), the frontal sinus (i), and the olfactory bulb (j).
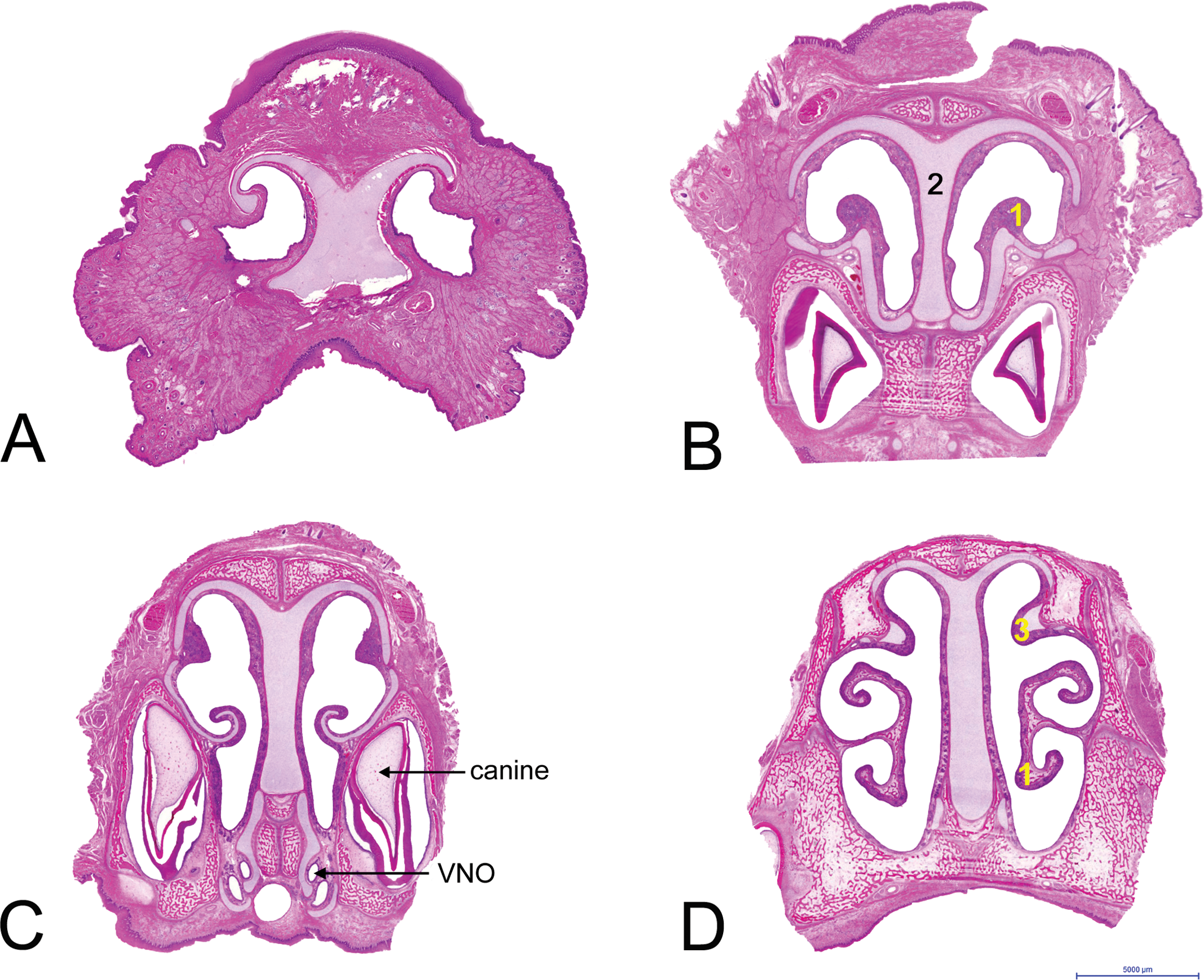
(A–J) Ten cross/transverse sections through the nasal passages of the 3-day-old minipig, from rostral to caudal. Level A is the vestibulum, lined by squamous epithelium. Level B includes the rostral part of the maxilloturbinates (1). The nasal septum (2) and the other parts of the passages are lined by squamous epithelium with some transitional epithelium on the maxilloturbinates. Level C includes the VNO. The passages are lined by transitional epithelium with squamous epithelium at the tips of the nasoturbinates and the opposing sides of the nasal septum, while the dorsal part of the maxilloturbinates is lined by respiratory epithelium. Levels D and E (level with sinus in nasoturbinates) are lined predominantly by respiratory epithelium, including the sinus in the nasoturbinates (3) at level E. The main arches at the dorsal meatus are covered by transitional epithelium in level D and with olfactory epithelium at level E. Levels F and G contain the olfactory bulb (4) and the ethmoturbinates (5) and are lined almost completely by olfactory epithelium, except for the choana/nasopharynx (6), which is lined by respiratory epithelium and divided into two by a septum. Levels H and I are at the nasopharynx above the soft palate, divided into two by a septum in level H. The soft palate contains the soft palate tonsil (7). Level J contains the nasopharynx with NALT, which is covered by lymphoepithelium (8). Hematoxylin and eosin–stained sections (approximately 3× enlarged).
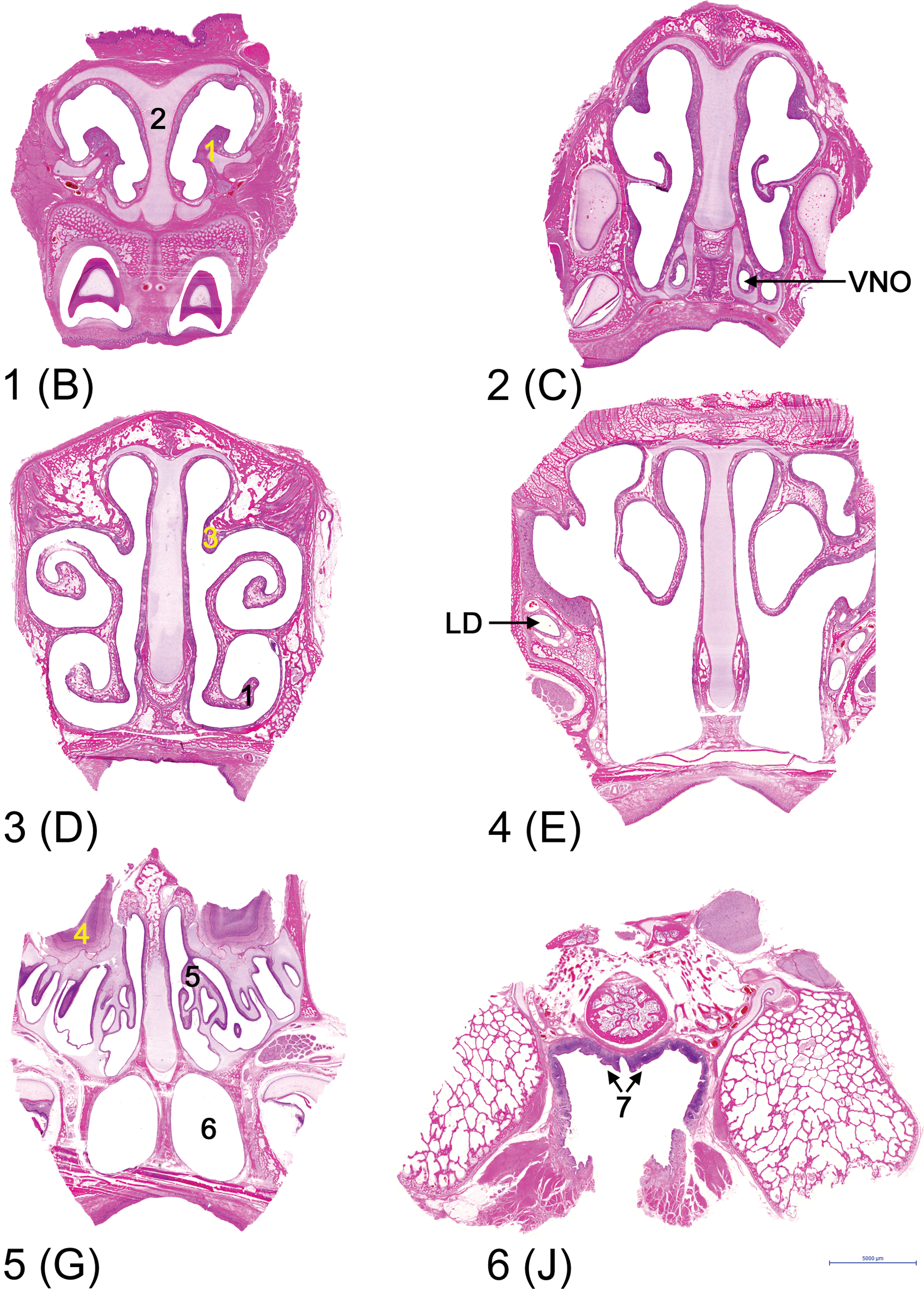
Cross sections at the 6 recommended levels through the nasal passages of the 8-week-old minipig, from rostral to caudal. The letters between brackets indicate the corresponding levels of the 3-day-old minipig depicted in Figure 3. (1) Maxilloturbinates, (2) nasal septum, (3) nasoturbinates, (4) olfactory bulb, (5) ethmoturbinates, (6) choana/nasopharynx, (7) NALT.
Results
Cross Sections of 3-Day-Old Minipig
The first cross section of the 3-day-old animal shows the vestibule (A in Figure 3), which is rather long in minipigs (Figures 1 and 2). At this level, the nasal cavity is supported by cartilage at the dorsal side only, allowing the tip of the nose to be quite flexible. The vestibulum was completely lined by squamous epithelium, with a few nasal glands in the mucosa of the lateral wall. Cross section 3B contained the rostral part of the maxilloturbinates and incisors. It was lined predominantly by squamous epithelium (Table 2), with some nasal glands in the mucosa of the lateral wall (Steno’s glands), the septum, and the turbinates. Cross section 3C contained the maxilloturbinates, the vomeronasal organ or organ of Jacobson (VNO), and the canine incisors. It was mainly lined by transitional (unciliated respiratory) epithelium, with slightly more nasal glands than in 3B. The maxilloturbinates were most prominent in level 3D, which included furthermore the rostral part of the nasoturbinates. The passages were lined mainly by (ciliated) respiratory epithelium. The amount of glandular tissue was comparable with cross section 3C. The microanatomy of the nasal passages from cross section 3D to the ethmoturbinates did not undergo significant changes except for the folding of the maxillo- and nasoturbinates, the increasing area of olfactory epithelium in the dorsal meatus, and the sinus within the nasoturbinates (Figure 3E). The sinus was lined by respiratory epithelium. Cross sections 3F and 3G included the ethmoturbinates, lined mainly by olfactory epithelium, and the nasopharynx, which was divided by a septum. A few Bowman’s glands were observed in the olfactory mucosa. The nasopharynx was lined by respiratory epithelium. From here on, the hard palate ended: cross sections 3H and 3I contained the soft palate with the soft palate tonsil and the nasopharynx, divided by a septum (3H) or undivided (3I). The nasopharyngeal septum ended just before the buccal opening of the nasopharynx. Cross section 3J included the nasopharynx with a dorsally located single NALT. NALT was covered by lymphoepithelium, bordered by respiratory epithelium.
Main histologic characteristics of the nasal passages of the minipigs at the recommended levels. a

a Applies to minipigs of all ages examined (1 day old to 6 months old).
Recommended Nasal Levels
Six levels were selected for standardized examination of the nasal passages of minipigs (levels depicted in Figure 4, 8-week-old minipig). Four levels were selected close to each other in the part caudal to the vestibulum, because the nasal passages demonstrated some distinct changes over this area. Level 1 (comparable to 3B) allows examination of the squamous epithelium. Level 2 (comparable to 3C) includes the VNO and a large area of transitional epithelium. Levels 3 (comparable to 3D) and 4 (comparable to 3E) contain mostly respiratory epithelium; level 4 contains also the most rostral part of the olfactory epithelium. Level 5 (comparable to 3G) includes the ethmoturbinates with olfactory epithelium, the olfactory bulb, and the nasal end of the choana/nasopharynx. The equivalent of 3G, rather than 3F was selected, because in older animals, the slice represented by 3F was too large to be embedded in the total. Level 6 includes the nasopharynx with NALT. Cross-cuts through the lacrimal duct were found at the base of the maxilloturbinates at level 3 to level 5 (3F–3G). At level 5 (3F–3G), a more longitudinal cut through the ducts was found in the lateral walls, but it was often trimmed in the older animals in order to fit the block into the cassettes.
Developmental Changes
Grossly, the nose lengthened (Figures 2A and B versus 2C and D) with age, and calcification increased. The height of the nasal passages relative to the width increased as well. The temporary deciduous canines were present at birth but showed distinct signs of wear in the 6-month-old animals. The microanatomy of the nasal passages did not change with age (1-day-old to 6-month-old animals), as illustrated in Figures 3 (3-day-old animal) and 4 (8-week-old animal), with the exception of the following aspects.
Until 8 weeks, the ethmoturbinates were cartilaginous (Figure 5A–D). Thereafter they ossified, which was completed at week 13. In addition, the caudal nasal septum (level 5) ossified, a process that was ongoing in 8- and 13-week-old animals, but 26-week-old (6-month-old) animals had a fully ossified septum at level 5.
The number of goblet cells in the respiratory epithelium was generally low, especially in the animals up to 4 weeks of age, but it increased slightly with age. Steno’s (lateral nasal) and Steno-like glands (septal and turbinate glands) were present in the mucosa of the squamous and respiratory epithelium, especially in the anterior part of the nasal passages of the minipigs. The amount of glandular tissue increased with age and appeared increasingly more active from day 14 onward (Figures 6A and B). Bowman’s glands or olfactory glands were found in the mucosa of the olfactory epithelium. In the 1- and 3-day-old piglets, ductlike structures appeared predominant (Figures 6C and D). In the older animals, the acini became abundant.
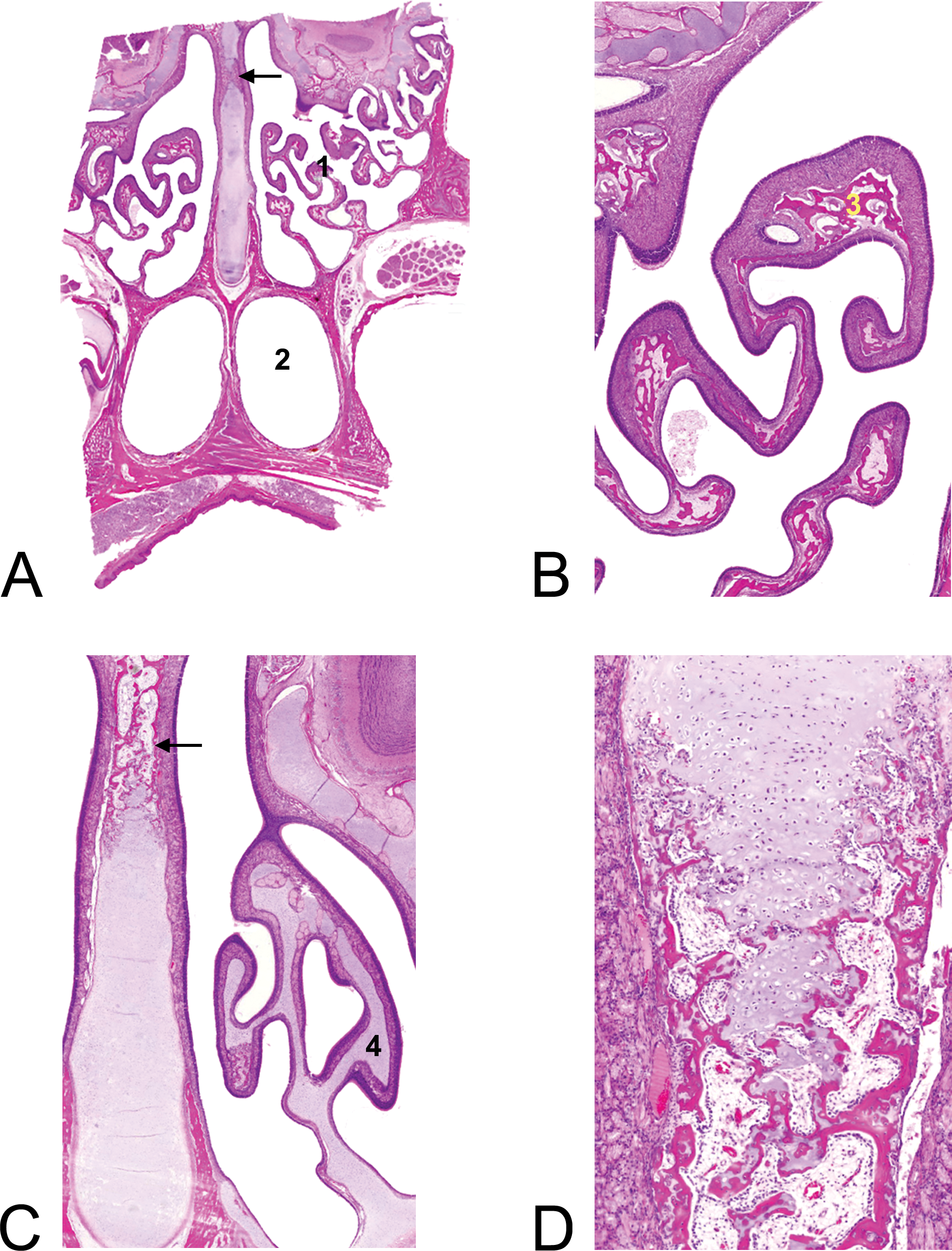
Ossification of nasal turbinates and septum with age. (A) Overview of nasal passages (level 5, comparable with Figure 3G and Figure 4-level 5) of a 13-week-old animal with ossified ethmoturbinates (1), nasopharynx (2), and ossification of septum (arrow). (B) Detail of the ossified (3) ethmoturbinates of the 13-week-old animal (20× magnification). (C) Cartilagenous (4) ethmoturbinates of an 8-week-old animal and ossification of septum (arrow). (20× magnification). (D) Detail of ongoing ossification of the septum, 8-week-old animal (50×). Hematoxylin and eosin–stained sections.
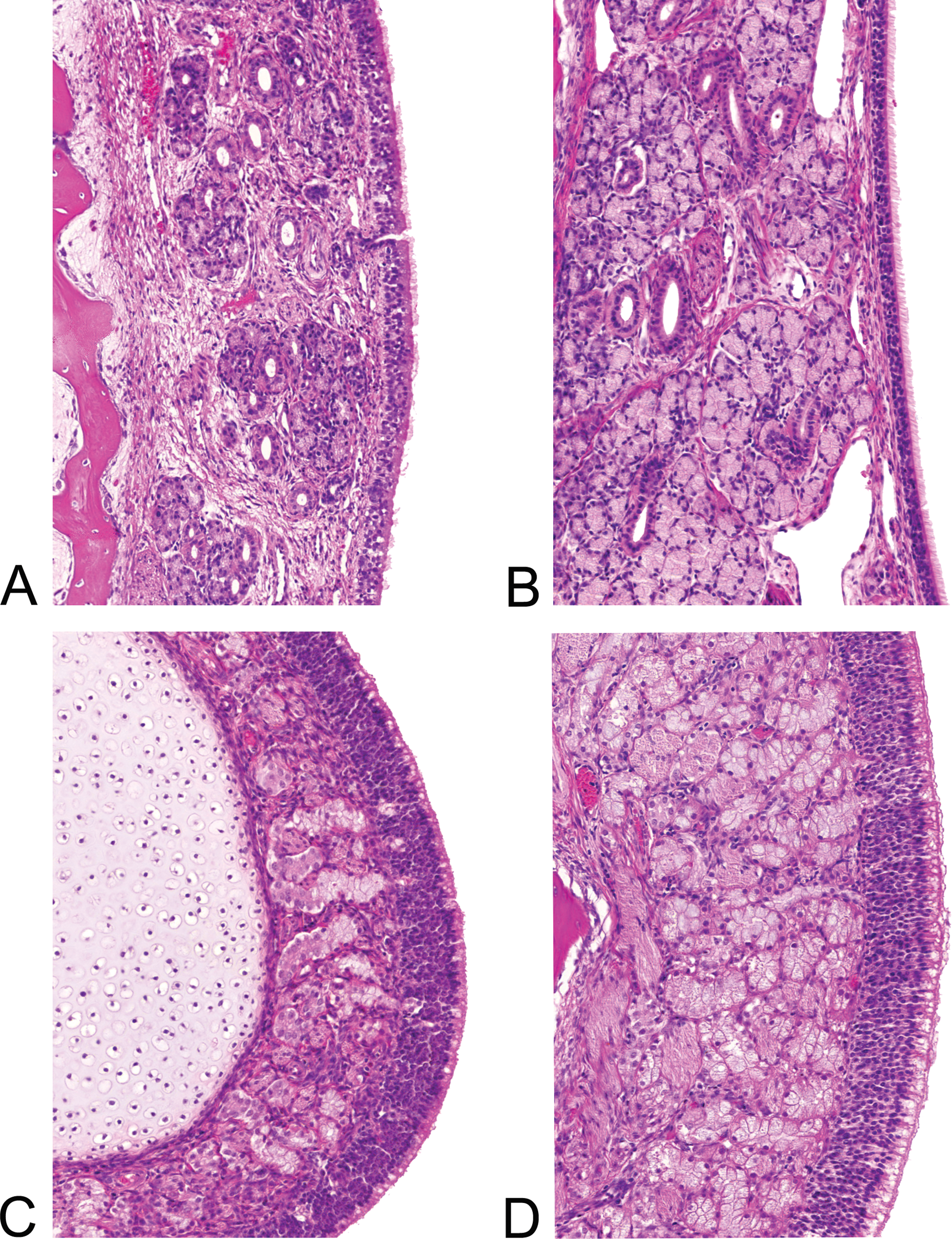
(A–D) Mucosal glands. Lateral nasal (Steno’s) glands in the lateral wall of the 3-day-old animal (A) and abundant glands in the 13-week-old animal (B). Bowman’s glands in development in the olfactory mucosa of the 3-day-old animal (C) and abundantly in the 26-week-old (6-month-old) animal (D). Hematoxylin and eosin–stained sections, 200× magnification.
The VNO was enclosed by a cartilaginous structure (Figures 3D and 4-2; detailed in Figure 7). More rostral in the nasal passages, the VNO was connected with the oral cavity via the nasopalatine duct, which was lined by stratified squamous epithelium. The VNO was lined laterally with respiratory epithelium and medially by receptor epithelium, as in other mammalian species, including man. Untill 6 months of age the size of the VNO itself and its glands increased. The VNO glands were scarce and not well developed before day 14, but they became extensive with age.
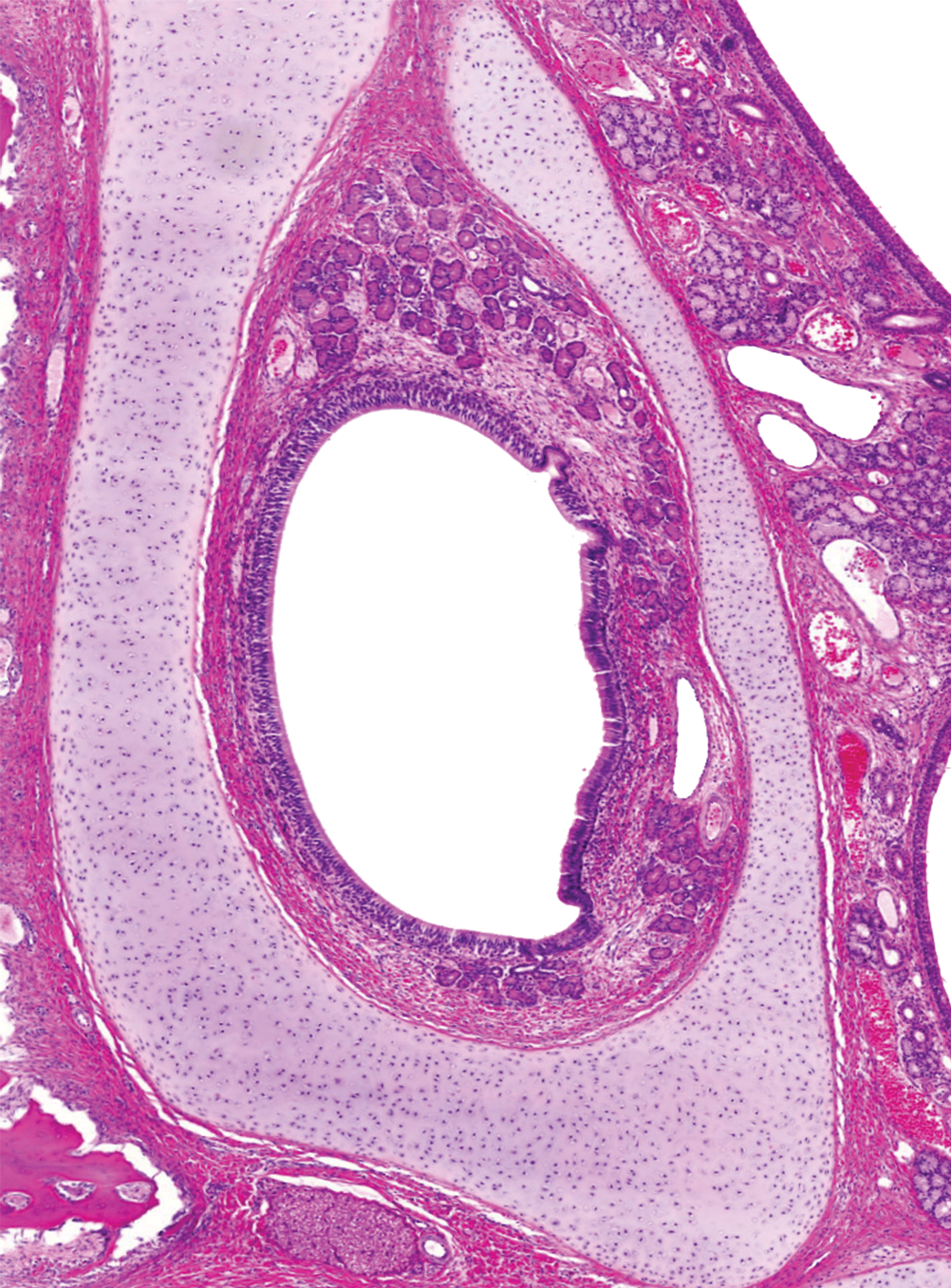
VNO in a 13-week-old animal showing the delicate cartilaginous structure surrounding the organ, the VNO glands, and the receptor epithelium at the medial side (left side in the picture). Hematoxylin and eosin–stained sections, 50× magnification.
NALT in minipigs is a single organ, located at the roof of the nasopharyngeal duct (Figure 3J). NALT was already present in the 1-day-old piglet, as a few scattered lymphoid cell accumulations. It rapidly increased in size and became activated with prominent germinal center development in the 3-day-old animal. The bulk of lymphoid tissue continued to increase in absolute size until the oldest, 6-month-old animal examined, although the cellular density decreased somewhat. The soft palate tonsil was present at birth as well and increased markedly with age.
Discussion
The nasal passages were examined of Ellegaard Göttingen minipigs, with ages ranging from 1 day to 6 months. Dental elements and palatal structures were used as landmarks to identify cross sections. The temporary deciduous canines were present at birth. A precise account of dental eruption is given by Weaver, Jump, and McKean (1969). Pigs are born normally with 8 sharp needle teeth, comprising the 4 canines and the third incisor. The sequence in which tooth eruption occurs depends on the strain, and within the strain there is variation among litters and individuals (Tucker and Widowski 2009), but the location of the dental elements is already visible at birth.
Six levels were selected for routine use. The selection was based primarily on the presence of the main nasal (accessory) structures and representative areas of the different epithelia in combination with the knowledge of target sites in other species. A cross section right through the vestibulum can be of interest in nasal application studies and in inhalation studies with highly reactive substances but is otherwise expected to show little or no histopathology (Morgan 1991). Therefore, cross sections in this area were not selected for routine use. Level 1 (Figure 4) provides information on the squamous epithelium; this epithelium was absent in the more caudal levels. Level 1 resembles the first level, proposed by Pereira, Macri, and Creasy (2011) for rabbits, except that in the minipigs, the nasoturbinates were not included at this level. Level 2 allows examination of the transitional epithelium. In rats, this epithelium was sensitive to aldehydes and ozone (Feron et al. 2001; Harkema and Hotchkiss 1994). Levels 3 and 4 were selected for the respiratory epithelium. Level 4 contained the most rostral part of the olfactory epithelium, which was found to be a sensitive site in the rat (Harkema, Carey, and Wagner 2006; Renne et al. 2007). Levels with the soft palate tonsil were left out, because the soft palate tonsil is associated with the oral cavity, rather than with the nasal passages. Level 5 contained the ethmoturbinates with olfactory epithelium and the rostral part of the choana/nasopharynx, while level 6 included NALT. Nasopharynx and NALT were included in the revised OECD412 guideline on 28-day inhalation toxicity (OECD 2009).
The elaborate ethmoturbinate and its relatively large olfactory bulb suggest that olfaction is superior to that of humans and monkeys with their simple ethmoturbinates and small olfactory bulb (Harkema, Carey, and Wagner 2006). The relative size of the olfactory bulb was largest in the young minipigs (Figure 2).
The nose enlarged and lengthened, and ossification of the turbinates and caudal part of the septum increased with the age of the animal. Enchondral ossification of the posterior part of the nasal septum has been observed also in man during the postnatal period (Verwoerd et al. 1989). The amount of glandular tissue increased with age and appeared increasingly more active. This is in accordance with the abundance of anterior glands in pigs, as described in the excellent overview of Bojsen-Møller (1967). Bowman’s glands or olfactory glands were found in the mucosa of the olfactory epithelium. In young animals, ducts were prominent, while acini were predominant in older animals. The VNO is a blind-ending tubular structure. It is enclosed by a cartilaginous structure as described by Salazar, Quinteiro, and Cifuentes (1997). Like in other mammalian species, it was lined laterally with respiratory epithelium and medially by receptor epithelium. A similar structure was observed in hematoxylin and eosin (H&E)–stained sections of VNO in man, but no immunohistochemical characteristics for receptor cells were found in adults (Troitier et al. 2000). Therefore, it is questionable how effects on VNO in laboratory animals should be translated to man. Apart from size, there were no distinct developmental changes until 6 months of age, like clustering of the receptor cells at the boundaries with the respiratory epithelium as occurs in rats from PN21 (Weiler, McCullouch, and Farbman 1999). The only distinct change was an increase in the VNO glands with age.
The NALT in minipigs is a single organ, located at the roof of the nasopharyngeal duct, comparable to the adenoid or nasopharyngeal tonsil in humans and dogs, although dogs reportedly have a far less conspicuous nasopharyngeal tonsil (Billen et al. 2006). This is in contrast to rodents, in which NALT is a paired organ, located at the ventral aspects of the walls of the nasopharyngeal duct, at the nasal entrance (Kuper et al. 1992). NALT in monkeys has been described as located on both the lateral and septal walls of the proximal nasopharynx (Harkema, Carey, and Wagner 2006). NALT was already present in the 1-day-old piglet, as a few scattered lymphoid cell accumulations; even a stillborn piglet already showed some lymphoid accumulations (unpublished observations). The bulk of lymphoid tissue continued to increase in absolute size until the oldest, 6-month-old animal examined. The soft palate tonsil was observed just anterior to NALT. Like NALT, it was present at birth. The entire ring of tonsils in the oronasopharynx is called Waldeyer’s ring; an extensive description of the tonsils in domestic and laboratory animals is given by Casteleyn et al. (2011).
In summary, a standardized method to examine the nasal passages of the minipig of different ages is feasible, using anatomic landmarks that are independent of the age of the minipig. Postnatal developmental changes are mainly confined to growth, ossification, and an increase in the amount of glandular acini.
Footnotes
Acknowledgments
The excellent technical assistance of Daryl Leydekkers and Tim van Olmen and the discussions with Dr. Marcel Wijnands are greatly appreciated.
The authors declared the following potential conflict of interest: the minipigs were received for free from Ellegaard.
The authors received no financial support for the research, authorship, and/or publication of this article.