
Abstract
The immune and hematopoietic systems play an important role in the normal homeostasis of blood and blood cells and for immune responses to endogenous and exogenous processes and insults. In order to interpret histopathologic changes in the immune and hematopoietic systems, it is important to understand the normal anatomy and histology of the thymus, spleen, lymph nodes, bone marrow, and other tissues. The thymus, spleen, and lymph nodes can be categorized by anatomical compartments, each of which contributes to specific immune functions. Lesions may be diagnosed by interpretive or descriptive (semiquantitative) methods. The interpretation of these tissues by lesion in anatomical compartments should allow for better understanding of these reactions and more definitive pathologic findings. Proliferative lesions may be difficult to differentiate from lymphomas and leukemias. The use of immunohistochemistry, compartmental pathology, and methods for the evaluation of clonality will make interpretation easier.
Keywords
Introduction
Pathologists who are not familiar with the histology and pathology of the immune and hematopoietic systems may find challenges in the diagnoses of lesions in these tissues (Germolec, Kashon et al. 2004; Germolec, Nyska et al. 2004). The background variation of lesions in rodent immune and hematopoietic tissues of controls also makes it more difficult to diagnose and interpret induced lesions. It is the aim of this manuscript to review the histology and histopathology of these tissues by compartment and with interpretative diagnoses.
Normal Anatomy and Histology
The normal anatomy and histology of the rodent hematopoietic and lymphoid systems has been described (Cesta 2006; Elmore 2006a; Linden et al. in press; Mebius and Kraal 2005; Ohtani and Ohtani 2008; Pearse 2006a; Suttie 2006; van den Broeck et al. 2006; Ward et al. 1999; Willard-Mack 2006). The lymph nodes of mice are generally very small, and routine sections can be at any angle through the node producing appearances of lesions when the node is actually normal. To avoid misdiagnosing normal tissue as a lesion, one should be familiar with the many possible cross- and tangential sections of the normal mouse node (Linden et al. in press). In rats, the nodes are larger, but the same problem can occur. Familiarity with normal histological variation in controls is essential. The rat spleen has a wide normal marginal zone, whereas the mouse marginal zone is narrower and varies in width by strain, stock, and line.
Reactive Lesions
Reactive lesions are any histological responses of cells and tissue to any insult or even a change in normal physiological processes. They can be categorized histologically by anatomical (compartmental) location in the spleen, lymph nodes, thymus, and Peyer’s patches. For lymph nodes, the compartments include the cortex (follicles, germinal centers, interfollicular areas), paracortex (T-cell–rich deep cortical unit), and medulla (medullary cords and sinuses; Willard-Mack 2006). For the spleen, the compartments include white pulp (B-cell–rich follicles, germinal centers and mantle zone, T-cell–rich periarteriolar lymphoid sheath, and macrophage rich marginal zone) and red pulp (sinuses and cords). The thymus is composed of the cortex and the medulla. Each compartment has its own types of cells, which may be unique for that zone or may be found elsewhere. Reactive lesions may be either hyperplastic or nonhyperplastic but rather composed of infiltrative cells. Cellular antigens are shown by immunohistochemistry to be expressed by the specific cell types (Rehg et al. in press; Ward 2006). The spleen, lymph nodes, and thymus also have capsules that may show reactive lesions. Bone marrow anatomy and histopathology have been described and will not be discussed in this paper (Travlos 2006).
The anatomical compartments of these tissues and their cellular components may react individually or collectively in response to insults to cells and tissues (Tables 1 and 2). The use of interpretative diagnoses (Ward 1990a; Ward 1990b; Ward et al. 1993) and of descriptive (semi-quantitative) terminology Elmore 2006a; Elmore 2006b; Elmore 2006c; Elmore 2010; Elmore, 2012; Haley et al. 2005; Kuper et al. 2000) are examples of the histopathologic evaluation of anatomical compartments for diagnoses. Interpretative diagnoses include terms such as atrophy and hyperplasia, whereas descriptive diagnoses indicate an increase or decrease of cellularity or area within a specific anatomic compartment (Elmore 2010; Elmore, in press). The descriptive diagnoses are especially helpful for immunotoxicity studies, which include various doses of the test chemical agent and time course sacrifices. Both methods can be graded for severity (Elmore, in press). Computerized image analysis may be used for more accurate interpretation of changes in lymphoid anatomy, morphology, and antigen expression. A good example of chemically induced lesions categorized by semiquantitative descriptive histopathological analysis has recently been published (Pearse et al. 2009).
Types of reactive lesions in lymph nodes of rodents.
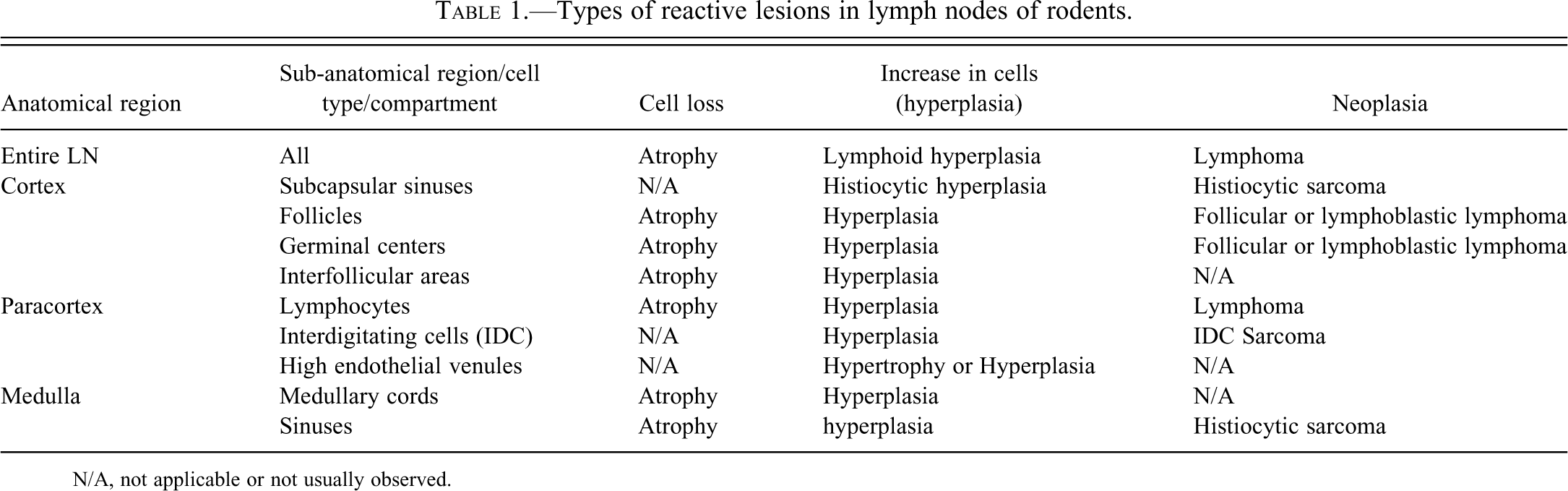
N/A, not applicable or not usually observed.
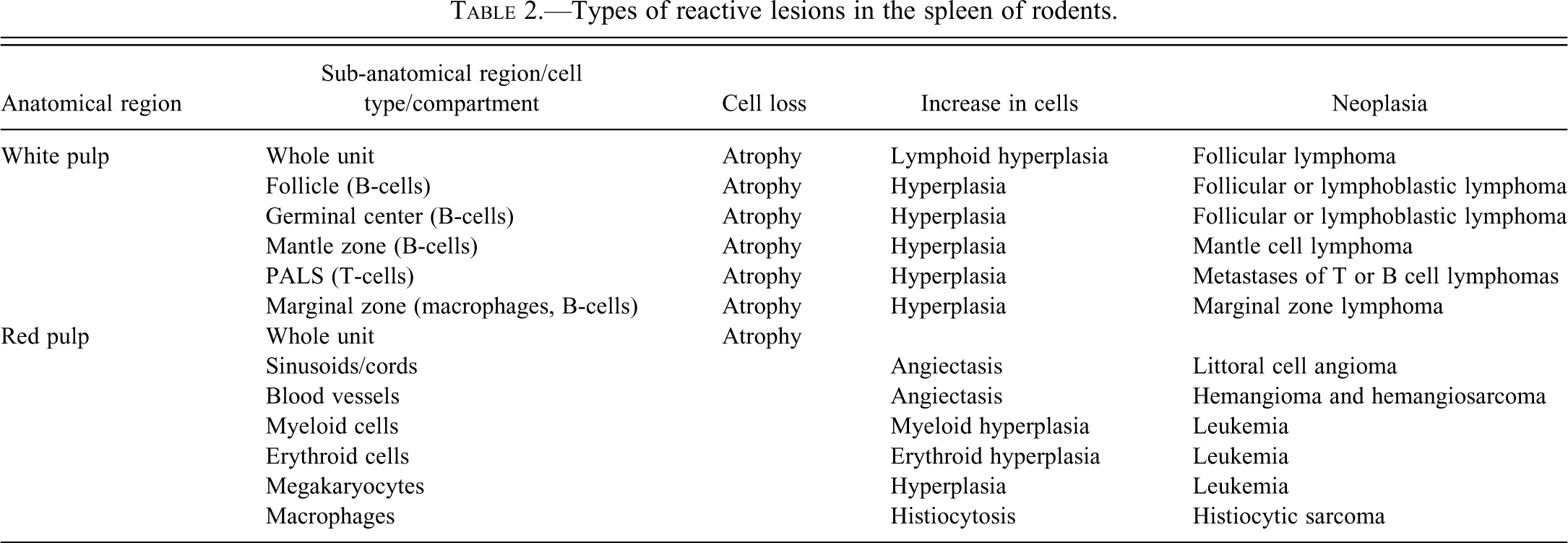
Types of reactive lesions in the spleen of rodents.
Age-associated changes in the lymphoid system may also be considered reactive to changes in the aging endogenous environment (hormonal, biochemical, molecular, genetic) and exogenous interactions (diet, air, water). Age-related lesions are common in the thymus and include atrophy (involution) in all species, glandular/epithelial hyperplasia in aging rats, and medullary and nodular hyperplasia in aging mice.
In humans, patterns of reaction in lymph nodes have been described as follicular and nodular, predominantly sinus, interfollicular, or mixed and diffuse (Jaffe et al. 2011). These patterns are well characterized for specific disease conditions in humans that are caused by genetic abnormalities, infectious agents, acquired disorders, and response to exogenous substances. The tissues of the rodent lymphoid system also respond in a similar manner, although the preferred terminology is slightly different.
Stepwise Histological Examination of Lymphoid Tissues
Optimal interpretation of a lymph node or spleen is best initiated with a review of the tissue at low magnification to observe the anatomical compartments and relationships to adjacent tissues (Table 3). All observations should be compared with those in normal or control tissues, and questions to be considered while reviewing the tissues are as follows. Are the normal anatomical structures in their usual order and of their usual sizes in relationship to each other, or is one compartment larger in comparison to others? Is more than one compartment abnormal in size or appearance? Is the node or spleen associated with local or systemic lesions? Can the histological appearance of the node be explained?
Stepwise histopathological analysis of the lymph nodes, spleen, and thymus.

Immunohistochemistry can aid in the interpretation of histological changes in lymph node, spleen, Peyer’s patch, thymus, or bone marrow. Each organ or tissue has unique normal antigen expression patterns because of its normal or abnormal cell populations. The normal cellular compartments in each organ can be compared with the affected tissue. For rodents, panels of antibodies have been validated for routine use in identifying specific cell types (Kunder et al. 2007; Mikaelian et al. 2004; Rehg and Sundberg 2008; Rehg et al. in press; Ward et al. 1993; Ward et al. 2006; http://tumor.informatics.jax.org/html/antibodies.html; http://www.niehs.nih.gov/research/atniehs/labs/lep/path-support/immuno/protocols.cfm; http://icg.cpmc.columbia.edu/cattoretti/Protocol/Mouse_IHC/Antibodies_for_mouse_IHC.html).
Proliferative Lesions of the Thymus
The thymus reacts in various ways to stress, toxins, and infectious agents (Pearse et al. 2006a,b). Atrophy (aging involution) and hyperplasia of anatomical and cellular compartments of the thymus are seen in aging rats and mice (Elmore 2006a; Pearse et al. 2006b). Epithelial cell and glandular hyperplasia are more commonly seen in aging rats, whereas aging mice develop medullary hyperplasia. When pronounced, thymic medullary hyperplasia, sometimes nodular, can mimic neoplasia in aging mice and in some lines of mice with autoimmune disorders. These hyperplastic lesions are not as large as tumor masses seen in thymic lymphomas, however, even when they are nodular (Figures 1–3). These hyperplastic foci may be composed of B-cells, which can form germinal centers and plasma cells (Figures 2 and 3).
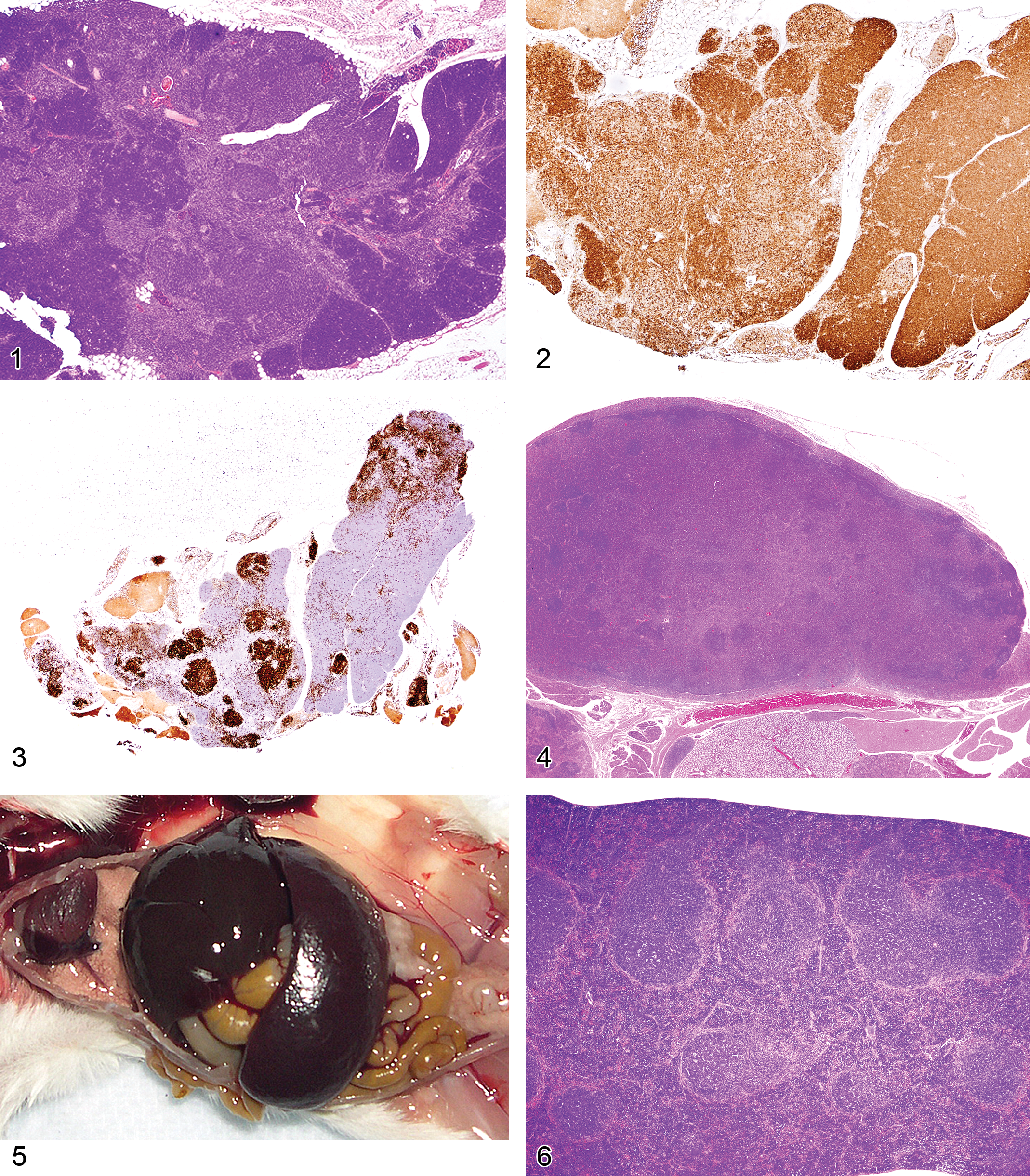
Figure 1 Thymus of an eighteen-month-old B6,129 mouse showing nodular medullary hyperplasia.
Genetically engineered mice with various altered genes may develop lymphomas that pass through histopathologic stages of unilateral or bilateral thymic cortical lymphocyte depletion, as well as medullary atypical hyperplasia (Dunnick et al. 1997; Ward et al. 1999). Atypical thymic medullary hyperplasia is characterized by unilateral or bilateral thymic lobular lesions and the appearance of immature lymphoid cells, including blasts with large nuclei. These lesions often progress into lymphoma with a grossly enlarged thymic lobe or lobes and metastases to the spleen, liver, and other tissues (Ward et al. 1999). The same histopathologic sequence can occur for thymic lymphomas induced by chemicals in mice and rats (Konishi et al. 1988), and by irradiation or viruses in mice.
Proliferative Lesions of Lymph Nodes
Lymph nodes often react to regional or systemic insults. The increase in lymph node size is often a result of enlargement of the various compartments. For example, the tissues affected by skin inflammation and ulceration drain via the lymphatics to the regional lymph nodes. The nodal responses may include inflammation with neutrophils, mast cells, germinal center hyperplasia, and the production of many plasma cells. Plasma cells may be abundant in chronic lesions, and the enlarged lymph node may be composed primarily of plasma cells and their precursors (Figure 4). Differentiation between plasma cell hyperplasia and a plasma cell lymphoma can be difficult. If only the regional node at the site of inflammation is involved, the change can readily be diagnosed as reactive hyperplasia. If generalized lymph node enlargement is seen, with many plasma cells in each enlarged node and no obvious etiology, assays for clonality can be performed (see below). Other reactive lesions in lymph nodes include germinal center hyperplasia associated with infectious diseases or other antigenic stimuli. This lesion is often accompanied by plasma cell hyperplasia in the medullary cords and cortex. T-cell hyperplasias in the paracortex may occur after viral infections. Unique lymphoproliferative disorders that involve the lymph nodes are also discussed below.
Proliferative Lesions of the Spleen
Proliferative splenic lesions can involve the white or red pulp, or both compartments at the same time. Lymphoid hyperplasia in the white pulp occurs after infections and inflammatory lesions in other tissues or as responses to tumors in any tissue. The hyperplasia may include germinal center hyperplasia, plasma cell hyperplasia, and general lymphoid hyperplasia of the many cell types that compose the white pulp. In experimental malaria infection of mice, spleens may attain 5 g in weight after seven to twelve days of infection because of a marked immune response in the spleen accompanied by erythroid hyperplasia (Waisberg et al. 2011). The white pulp is much enlarged, mostly because of germinal center and plasma cell hyperplasia (Figures 5–7). The red pulp is expanded by many erythroid precursors.
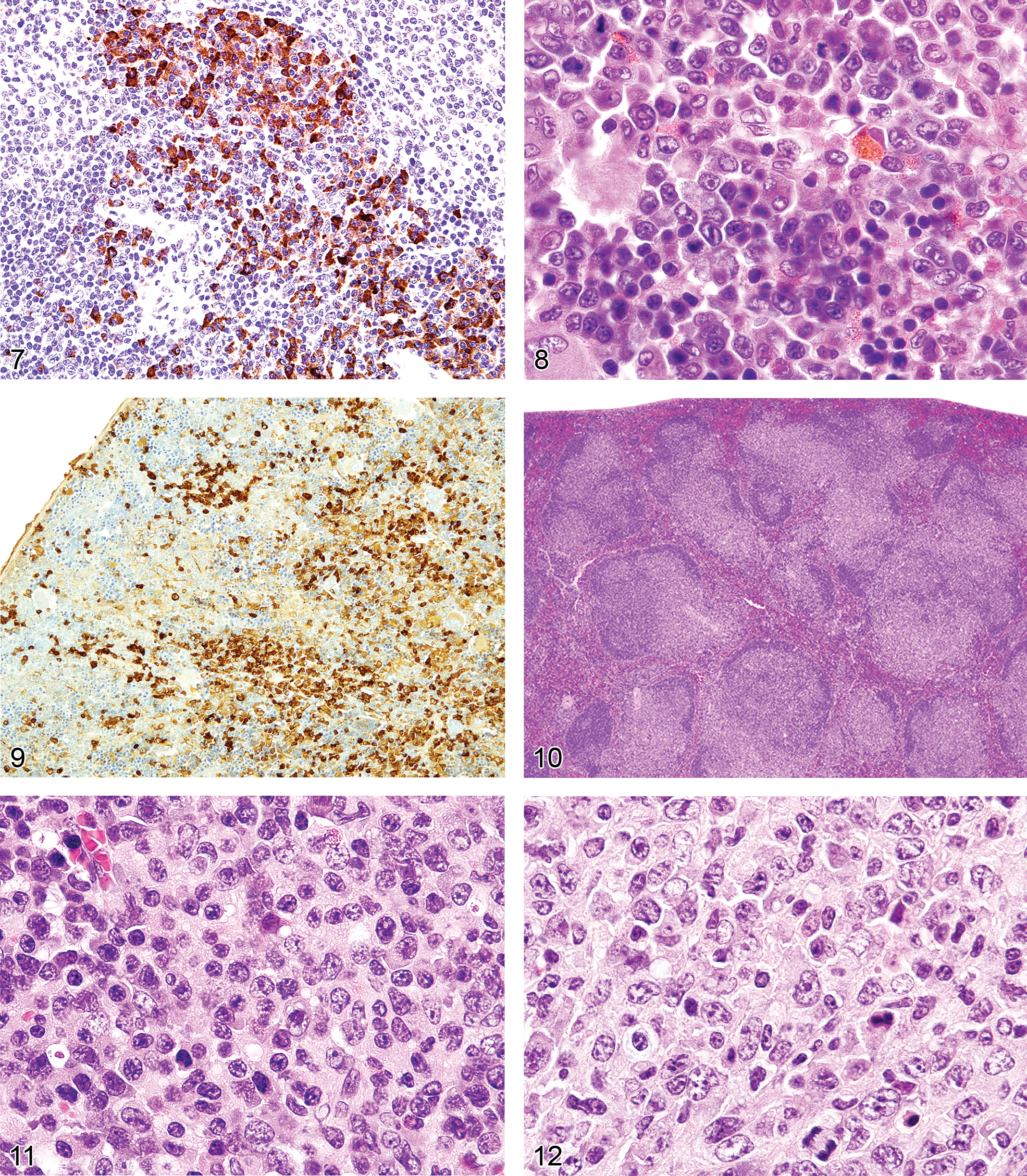
Figure 7. Spleen in a malaria-infected mouse with numerous plasma cells. Immunohistochemistry for human κ light chains expressed mostly in the plasma cells.
Splenic Myeloid and Erythroid Hyperplasia
The spleens of rats and mice are highly reactive to endogenous lesions, including inflammation and neoplasia. Inflammatory lesions in various organs, especially in the skin, lung, and intestine, can stimulate myelopoiesis in the bone marrow, spleen, and other tissues. The spleen is particularly susceptible to enlargement caused by myeloid and erythroid hyperplasia, which are often accompanied by megakaryocyte hyperplasia. Immature myeloid and erythroid precursors can predominate in such reactions, mimicking leukemia. These spleens often exhibit lymphoid hyperplasia in the white pulp, in response to antigens expressed by tumors or infectious agents. Evaluation of the anatomical appearance of the spleen and its compartments at low magnification is a first step in histological evaluation. The appearance under high magnification of mature and immature hematopoietic cell types may reveal differences in the ratio of the most immature cells to mature cells (Figure 8). These lesions are often accompanied by plasma cell hyperplasia (Figure 9). If a majority of the cells in a compartment are of a similar immature cell type, this finding may be indicative of neoplasia. If the cause of a splenic response (inflammatory and ulcerative skin lesions, tumor, or internal inflammatory lesions in other tissues) is found, the splenic hyperplastic state can be explained as a response to those lesions. Sometimes, a lesion cannot be found to explain the splenic reaction. Detailed suggestions for histopathological differentiation of hyperplasia versus leukemia have been reported (Long et al. 1986; Ward 1990). One should be conservative and not diagnose leukemia unless there is overwhelming evidence to support the diagnosis.
Lymphoproliferative Disorders
Lymphoproliferative disorders (LPD) have been described in humans (Swerdlow et al. 2008), monkeys (Schmidtko et al. 2002), and less often, in mice. In humans and monkeys, they may occur naturally (of genetic or other origin) or from inadvertently induced immune disorders (herpes viral infections or after drug-induced immunosuppression during organ transplants). In mice, they are of genetic origin or induced by experimental procedures such as viral infection. Lymphoproliferative disorders are characterized by a non-neoplastic proliferation of lymphocytes in one or more lymphocyte lineages in the various lymphoid and other tissues. Their non-neoplastic nature may be shown by laboratory assays demonstrating lack of clonality or other characteristics. These conditions may progress to lymphomas, especially in monkeys and humans, when caused by drug-induced immunosuppression and herpes virus infection (Swerdlow et al. 2008; Schmidtko et al. 2002). In mice, LPD has been described in the SJL/J strain (Tang et al. 1998) and in mice with naturally occurring mutations in Fas (lpr) and Fasl (gld; Cohen and Eisenberg 1991; Davidson et al. 1998), and it has also been induced by viruses including a mutant retrovirus, LP-BM5 murine leukemia virus (MuLV), which causes murine acquired immunodeficiency syndrome (MAIDS) (Hartley et al. 1989; Hartley et al. 2000; Klinken et al. 1988), the γ herpes virus, MHV68 (Barton et al. 2011), and mouse cytomegalovirus (CMV; Karupiah et al. 1998). Lymphoproliferative disorders have also been described in genetically engineered mice, including mice bearing transgenes for IL6 (Kovalchuk et al. 2002), Myc (Adams et al. 1985; Park et al. 2005), BCL6 (Cattoretti et al. 2005), and BCL10 (Li et al. 2009).
The gross lesions of lymphoproliferative disorders in mice vary from a massive lymph node enlargement to moderate enlargements of the lymph nodes and spleen. In the spleen, the white pulp usually expands (Figure 10) as a result of proliferation of a uniform population of mature lymphoid cells of a single lineage (T-cells, as in gld or lpr mice) or, more often, of a mixed population of mature B- and T-cells and plasma cells (Figure 11), especially in the early stages of MAIDS and the other disorders. This early stage can be followed by the appearance of a neoplastic population of blastic lymphoma cells with many mitotic figures (Figure 12). Although nonlymphoid organs can be involved, they need not be. The morphology and antigen expression patterns of the lymphoid cells can help differentiate the lesions from lymphomas.
Unique genetic disorders in mice, which result from the gld or lpr mutation, involve genes similar to those in humans causing similar LPD. They have an initial proliferative non-neoplastic stage composed of a uniform population of T-cells (Figures 13 and 14). The lesion appears morphologically as a lymphoma of medium-sized lymphocytes that express CD3 and CD45R/B220 but that are not clonal. With time, the lesions progress into a plasmacytoma composed of a pleomorphic population of B-cells producing immunoglobulins (Davidson et al. 1998).
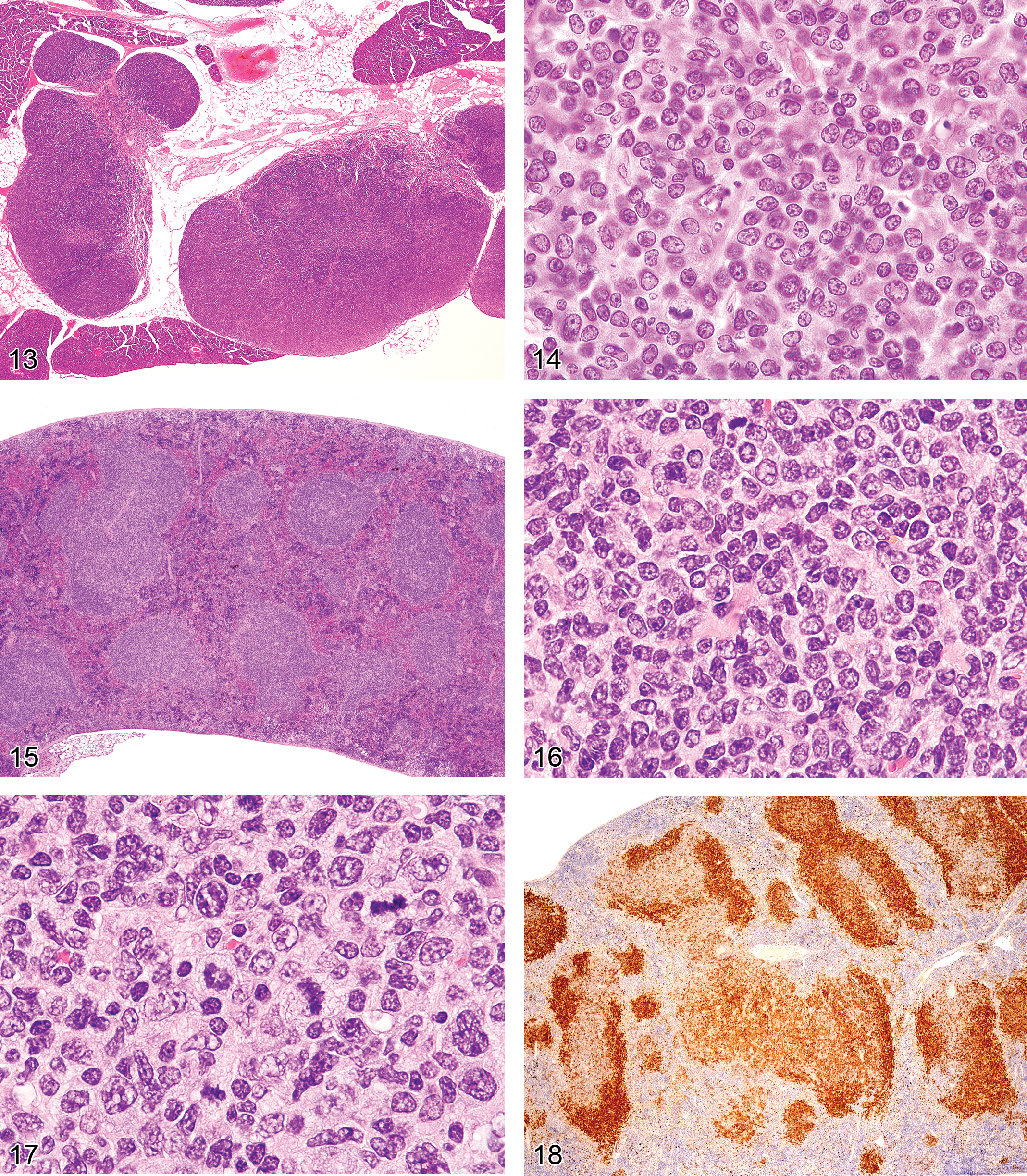
Figure 13. Greatly enlarged lymph nodes of a six-month-old C3H/lpr mouse with lymphoproliferative disease.
If a chemical induces a lymphoproliferative lesion that does not appear to be a lymphoma, one should try to characterize the nature and cause of the lesions. For example, in monkeys and humans, post-transplant LPD often occur one to six months after the initial immunosuppressive treatment. In many cases, herpes viruses have been shown to be responsible (Swerdlow et al. 2008; Schmidtko et al. 2002). The herpes virus– and retrovirus-induced LPD of mice are generally composed of several cellular components and can be differentiated from typical mouse lymphomas (Klinken et al. 1988).
With time, several of the LPD of mice that are initially polyclonal have been shown to evolve in the same lymphoid tissues into clonal B-cell lineage lymphomas, primarily of germinal center and postgerminal center origin. These include follicular and diffuse large B-cell lymphomas (SJL/J) and immature to fully differentiated plasma cell neoplasms (SJL/J, MAIDS, lpr, gld) and marginal-zone lymphomas (p53, BCL10).
Early Lymphomas in the Lymph Nodes, Spleen, and Peyer’s Patches
In aging control mice of various strains, stocks, and lines, lymphomas often arise in the splenic white pulp, mesenteric lymph node cortex, and/or Peyer’s patches (Ward 2006). These lesions can be seen in some genetically engineered mice, that develop B-cell lymphomas. These anatomical locations are also the sites of reactive lesions that can be similar to pre-lymphomatous lesions. The classification and nomenclature of mouse hematopoietic neoplasms is well described (Dunn 1954; Kogan et al. 2002; Morse et al. 2002).
It is important to differentiate the type of lymphomas that develop in controls from those that appear in treated mice (Ward 2006). The age of the mouse and the anatomical locations of the primary and metastatic or multicentric lymphomas are important, as well as phenotype (T-cell, B-cell, etc.). For example, it is unusual for lymphomas to arise in the thymuses of young mice, except for specific mouse strains such as AKR. Young CD1 mice and rats appear to have a low incidence of thymic lymphomas. Tumors induced in genetically engineered mice and by chemicals or irradiation often arise in the thymus (Ward et al.1999). If lymphomas are disseminated at the time of illness or death, it may be difficult to determine the origin of the lymphomas.
Early lymphomas in the splenic white pulp may be focal, multifocal, or diffuse (Ward 1990c). The occurrence of early lymphomas in the spleen, mesenteric or other lymph nodes, and Peyer’s patches may also be restricted to one organ or be multifocal or diffuse in several organs (multicentric). At low magnification, the splenic white pulp enlarges and appears less basophilic than the normal white pulp areas (Figure 15). The normal white pulp has areas with small lymphocytes (Figure 16), and in the germinal center, the centroblasts and centrocytes occupy different zones (polarization) and tingible body macrophages are present. Early lymphoma lesions in splenic white pulp may be seen within follicular areas or even in the periarteriolar lymphoid sheath. Although enlargement of white pulp is often seen in reactive splenic lymphoid hyperplasia, the morphology of immature lymphoid cells in early follicular lymphomas (often centroblasts and centrocytes) is unique (Figure 17). The early lymphoma cells can be blasts with round large nuclei, cells with cleaved (folded) nuclei, and other immature lymphoid cells in various proportions. Tingible body macrophages, a characteristic of reactive hyperplasia, are absent, and the centroblasts and centrocytes are randomly distributed. If plasmacytoid cells are present, they have to be differentiated from a plasma cell reaction.
Immunohistochemistry can identify different gene expression patterns for early lymphomas in the white pulp areas (Ward et al. 1999). The areas are larger than normal white pulp areas and often express more or less antigen (Figure 18). In lymph nodes and Peyer’s patches, spontaneous lymphomas may arise in the cortex from follicular areas and may also show a change in the normal B-cell antigen (CD45R, PAX5) expression pattern. Induced lymphomas of the spleen, Peyer’s patches, and lymph nodes may also less commonly arise from germinal centers.
Proof of Clonality (neoplasia)
There are several methods for determining whether presumed tumor populations are clonal. Proof of clonality would strongly suggest neoplasia rather than hyperplasia. For lymphoid disorders, analyses of DNA for the genomic organization of immunoglobulin heavy chain (IgH) and κ light chain (IgK) and T-cell receptor β chain (TCRβ) loci using Southern blot hybridization with locus-specific probes have been the gold standard (Davidson et al.1998; Fredrickson et al.1993; Hao et al. 2010; Hartley et al. 2000; Klinken et al. 1988). Infectious ecotropic murine leukemia viruses (MuLV) can contribute to transformation by insertional mutagenesis. Clonality can also be demonstrated by the presence of new somatically acquired proviral integrations identified by Southern blot hybridization of genomic DNA using an ecotropic MuLV-specific probe. This method is useful for examining nonlymphoid as well as lymphoid neoplasms (Morse et al. 2001). Clonal chromosomal aberrations, including translocations and aneuploidy, can be revealed using spectral karyotyping of metaphase spreads (Qi et al. 2010). This approach can be applied to analyses of lymphoid and nonlymphoid neoplasms. Also, TCRβ gene rearrangements can be amplified for sequence analysis using degenerate RT-PCR (Choi et al. 2009) or for IgH using primers that detect rearrangements of diversity and joining regions from IgH genes by PCR (Xiang et al. 2011).
Summary
Reactive lesions in lymphoid tissues commonly occur with aging and under experimental conditions. The lesions follow a pattern involving specific anatomical compartments or multiple compartments of the lymph nodes, spleen, and other tissues. An interpretive diagnostic scheme is presented. Differentiation of reactive (often hyperplastic) lesions from early neoplastic lesions may be a difficult task. Patterns of tissue and cell reactions, LPD, early neoplasia, and neoplasia can be readily defined. Clonality assays can be important. This review provides information for differentiating reactive from neoplastic lesions.
Footnotes
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: in part, by the Intramural Research Program of the NIH, National Institute of Allergy and Infectious Diseases.