
Abstract
Seven-week-old female BALB/c mice received a single intraperitoneal injection of N-ethyl-N-nitrosourea (ENU) (50, 100, 200, 400, or 600 mg/kg), and retinal damage was evaluated after 7 days. Sequential morphological features of the retina and retinal apoptosis, as determined by the TUNEL assay, were analyzed 6, 12, 24, and 72 hr and 7 days after treatment with 600 mg/kg of ENU. Moreover, older mice (25 to 34 weeks of age) received an intraperitoneal injection of 600 mg/kg ENU and were sacrificed 7 days later. All animals were necropsied, and both eyes were examined histopathologically. Two of the 5 mice that received 600 mg/kg ENU died during the experimental period. Histopathologically, all mice that received 600 mg/kg of ENU experienced retinal degeneration characterized by the loss of photoreceptor cells (disappearance of the outer nuclear layer and photoreceptor layer) in both the central and peripheral retina within 7 days. One of 5 mice treated with 400 mg/kg ENU exhibited retinal damage that was restricted to the central retina. Older mice treated with 600 mg/kg ENU exhibited retinal damage that was similar to the retinal damage in younger mice. In the 600 mg/kg ENU-treated mice, TUNEL-positive photoreceptor cells peaked 72 hr after ENU treatment. Retinal thickness and the photoreceptor cell ratio in the central and peripheral retina were significantly decreased, and the retinal damage ratio was significantly increased 7 days after treatment. In conclusion, ENU induces retinal degeneration in adult mice that is characterized by photoreceptor cell apoptosis.
Introduction
N-ethyl-N-nitrosourea (ENU) is an alkylating agent that is particularly effective in inducing congenital malformations (Diwan 1974; Nagaishi 1982), including anophthalmia, microphthalmia, and bulged or open eyes; and tumors in various locations, including the nervous tissue, gastrointestinal tract, pancreas, respiratory tract, skin, mammary gland, kidney, and lymphoreticular tissues, in rodents (WHO IARC 1998). Prenatal exposure to ENU is particularly effective in producing tumors of the nervous system, such as glioma (Barth and Kaur 2009). Moreover, ENU mutagenesis has been used to create a large number of random point mutations in the genomic DNA of mice and zebrafish, and it is a powerful tool for creating disease models (Justice et al. 1999; Probst and Justice 2010; Wagner et al. 2010), such as morphological defects in eyes (Baird et al. 2002; Favor and Neuhauser-Klaus 2000; Wagner et al. 2010) and retinal degeneration in the progeny (Maaswinkel, Mason, and Li 2003; Liu et al. 2010; Sato et al. 2010).
Inherited night blindness is a fairly widespread disease in humans, and it affects approximately 1 in 5,000 individuals worldwide. A common form of inherited blindness is retinitis pigmentosa, which is a degenerative pigmentary retinopathy that is noninflammatory, bilateral, and progressive. The loss of photoreceptors is followed by perivascular pigment deposition within the retina (Hartong, Berson, and Dryja 2006; Shintani, Shechtman, and Gurwood 2009). Animal models of retinal degeneration are important for elucidating the mechanism of human retinitis pigmentosa (Rivas and Vecino 2009; Sancho-Pelluz et al. 2008) and exploring potential treatments (Shintani, Shechtman, and Gurwood 2009; Tsubura et al. 2010). Mutant mouse models for retinitis pigmentosa have been described (Dalke and Graw 2005), and mice carrying the rd (rodless retina or retinal degeneration) gene or rds (retinal degeneration slow) gene experience a final common cell death feature, namely, photoreceptor apoptosis (Chang, Hao, and Wong 1993; Sancho-Pelluz et al. 2008). Rodent eyes are highly sensitive to toxic substances. The retinal degeneration (photoreceptor cells loss) induced by N-methyl-N-nitrosourea, an alkylating agent (Gnewuch and Sosnovsky 1997), has been well studied (Tsubura et al. 2010; Yoshizawa et al. 1999; Yuge et al. 1996). In contrast, few studies have focused on ENU-induced retinal damage in adult rodents.
The aim of the present study is to elucidate the effect of ENU on the photoreceptor cells in adult BALB/c mice. The involvement of photoreceptor cell apoptosis stimulated by ENU is also discussed.
Materials and Methods
Animals
Fifty female SPF/VAF mice (BALB/c AnNCrlCrlj) were purchased at 6 weeks of age from Charles River Japan (Osaka, Japan). Mice were maintained in specific pathogen-free conditions and received free access to a commercial diet (CMF 30Gy; Oriental Yeast, Chiba, Japan) and water. Animals were housed in plastic cages with paper-chip bedding (Paper Clean, SLC, Hamamatsu, Japan) in an air-conditioned room at 22 ± 2°C and 60 ± 10% relative humidity with a 12-h light/dark cycle. The illumination intensity was below 60 lux in the cages. All procedures were in accordance with the guidelines for animal experimentation at Kansai Medical University.
Chemical and Dose Formulation
ENU (ISOPAC; chemical formula, C3H7N3O2) was purchased from Sigma-Aldrich Japan (Tokyo, Japan) and kept at –80°C in the dark. ENU was dissolved in physiologic saline just prior to use.
Experimental Procedures
Seven-week-old mice received an intraperitoneal (ip) injection of vehicle (physiological saline) or ENU at a dose of 50, 100, 200, 400, or 600 mg/kg (5 mice per dose). Seven days after the injection, mice were anesthetized with isoflurane (Forane; Abbot Japan, Tokyo, Japan) and sacrificed by exsanguination from aortic transection. In another experiment, 20 mice received a single ip injection of 600 mg/kg of ENU, and 4 randomly selected mice were sacrificed at 5 time points (6, 12, 24, and 72 hr and 7 days) after treatment. The control group consisted of 4 mice that were treated with vehicle only. To examine the susceptibility to ENU in relationship to aging, 6 female mice at 25 and 34 weeks of age received an ip injection of 600 mg/kg ENU or vehicle and were sacrificed 7 days after treatment. All mice were observed daily for clinical signs of toxicity and were weighed at the time of ENU treatment and on the day of sacrifice. Both eyes were quickly removed at the time of sacrifice, and complete necropsies were conducted on all animals.
Tissue Fixation and Processing
One eye from each mouse was fixed overnight in 10% neutral buffered formalin, and the other eye was fixed overnight in methacarn. Subsequently, the eyes were embedded in paraffin, sectioned at a thickness of 4 µm, and stained with hematoxylin and eosin (HE). Ocular sections were cut along a line parallel to the optic axis and nerve (including the ora serrata). To examine the effects of ENU on immunosuppression and hematopoiesis, the thymus and spleen were fixed in 10% neutral buffered formalin, sectioned to prepare HE-stained slides, and examined histopathologically.
TUNEL Staining
Formalin-fixed sections were used for cell death analysis. Cell death was evaluated by terminal deoxynucleotidyl transferase (TdT)–mediated dUTP digoxigenin nick end-labeling (TUNEL) using an in situ apoptosis detection kit (Apop-Tag; Millipore, Billerica, MA, USA) according to the manufacturer’s instructions.
Morphometric Analysis of Retinal Thickness, Photoreceptor Cell Ratio, and Retinal Damage Ratio
HE-stained sections and TUNEL-stained sections of the retina were scanned with a high-resolution digital slide scanner (NanoZoomer 2.0 Digital Pathology, Hamamatsu Photonics, Hamamatsu, Japan) to prepare digital images. The ndpi image files were opened in color mode with NDP.view software (Hamamatsu Photonics). The total retinal thickness (from the internal limiting membrane to the pigment epithelium), inner retinal thickness (from the internal limiting membrane to the outer plexiform layer), and outer retinal thickness (from the outer nuclear layer to the pigment epithelial cell layer) were individually measured from methacarn-fixed HE slides using NDP.view, as described in our previous reports (Yoshizawa et al. 1999, 2009). The measurements were conducted at the central retina (approximately 400 µm from the optic nerve) and peripheral retina (approximately 400 µm from both sides of the ciliary bodies). To further evaluate the photoreceptor cell loss, the photoreceptor ratio [(outer retinal thickness/total retinal thickness) × 100] was calculated. To determine the area of retinal damage, the entire length of the retina and the length of the damaged area in HE preparations were measured. Damaged retina was designated as the presence of less than 4 rows of photoreceptor nuclei in the outer nuclear layer (Yoshizawa et al. 1999, 2009), and the retinal damage ratio was calculated as (length of damaged retina/whole retinal length) × 100. The apoptotic index was calculated from TUNEL-stained slides by determining the number of apoptotic nuclei per microscopic field at three central locations and three peripheral locations of the retina. The images were captured with a 40-fold objective magnification. Histopathological and morphometrical evaluations were performed by a toxicologic pathologist certified by the Japanese Society of Toxicologic Pathology (K.Y.) and an ophthalmologist certified by the Japanese Ophthalmological Society (M.K.), according to the previously defined histopathological terminology and diagnostic criteria (Yoshizawa et al. 1999, 2009).
Statistical Analysis
All discrete values, expressed as the mean ± standard error (SE), were analyzed using the two-tailed independent Student’s t-test for unpaired samples after confirming the homogeneity of variances. The results presented below include comparisons between ENU-treated mice and vehicle-treated mice. P-values < .05 were considered to show statistical significance.
Results
General Remarks
The mortality and body weight of ENU-treated mice were affected in a dose-dependent manner. Mice treated with 600 mg/kg ENU exhibited decreased locomotor activity and closed eyes from the day of treatment; 2 of the 5 mice in this group died within 5 days. Although ENU did not cause weight loss in the 7-week-old mice, weight gain (the growth rate) was inversely related to the ENU dose; there was a statistically significant decrease in the growth rates of the 400 and 600 mg/kg ENU groups as compared to the control group (data not shown). The older mice (25 and 34 weeks old) treated with ENU displayed similar clinical signs and a decrease in body weight, but none of these mice died during the experimental period.
Morphological and Morphometric Analysis of Dose-Dependent Retinal Damage
Retinal histology of ENU- and vehicle-treated mice was compared 7 days after ENU treatment. The retinas of vehicle-treated mice contained more than 10 layers of photoreceptor nuclei in the central retina and more than 8 layers of cells in the peripheral retina (Figure 1 ). In contrast, all mice treated with 600 mg/kg ENU had 0 or only a few rows of photoreceptor cell nuclei in both the central and peripheral retina (Figure 1). The remaining photoreceptor nuclei in ENU-treated mice 7 days after the treatment were densely stained and contained clumped chromatin, and some large basophilic bodies were observed between the inner nuclear cells and Bruch’s membrane. One of the 5 mice that received 400 mg/kg ENU had fewer rows of photoreceptor cell nuclei at the central retina than the controls (Figure 1), while the retinas of the other 4 mice appeared to be normal. The ENU-induced changes were restricted to photoreceptor cells; all other layers of the retina remained intact. The retinas of all mice that received 200 mg/kg ENU or less were histopathologically intact (Figure 1). Seven days after treatment with 600 mg/kg ENU, the total retinal thickness and outer retinal thickness were significantly decreased in both the central and peripheral retina, as compared with the vehicle-treated control mice (Figure 1). In mice treated with 400 mg/kg ENU or less, the mean total retinal thickness and the outer retinal thickness at both the central and peripheral retina were not significantly different than in control mice, although damage did occur to the central retina of one mouse treated with 400 mg/kg ENU. No changes to the inner retinal thickness were observed in any of the ENU-treated groups (data not shown). To further evaluate the ENU-induced effects on retinal thickness, the photoreceptor cell ratio was calculated (Figure 2a and b). The photoreceptor cell ratio at the central and peripheral retina was 52% and 54%, respectively, for vehicle-treated mice but only 21% and 5% (p < .01), respectively, for 600 mg/kg ENU-treated mice. The photoreceptor cell ratio at the central and peripheral retina was not significantly different in mice treated with 400 mg/kg ENU or less (46% and 55% in 400 mg/kg group, 52% and 55% in 200 mg/kg group, 51% and 54% in 100 mg/kg group, and 49% and 55% in 50 mg/kg group, respectively).
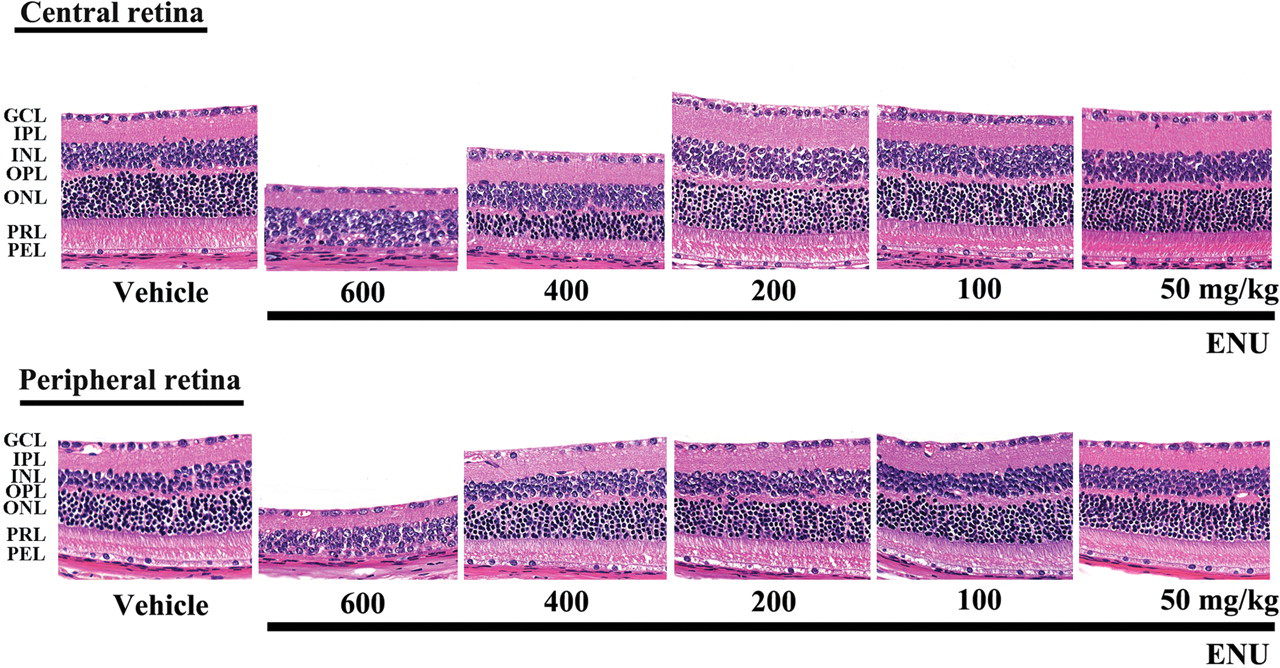
Retinal change in adult mice 7 days after a single intraperitoneal (ip) injection of ENU. The outer nuclear layer and photoreceptor layer disappeared in both the peripheral and central retina of mice that received 600 mg/kg ENU. Moderate retinal degeneration was detected only in the central retina of one 400 mg/kg ENU-treated mouse. GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; PRL, photoreceptor cell layer; and PEL, pigment epithelial cell layer. HE staining, ×400.
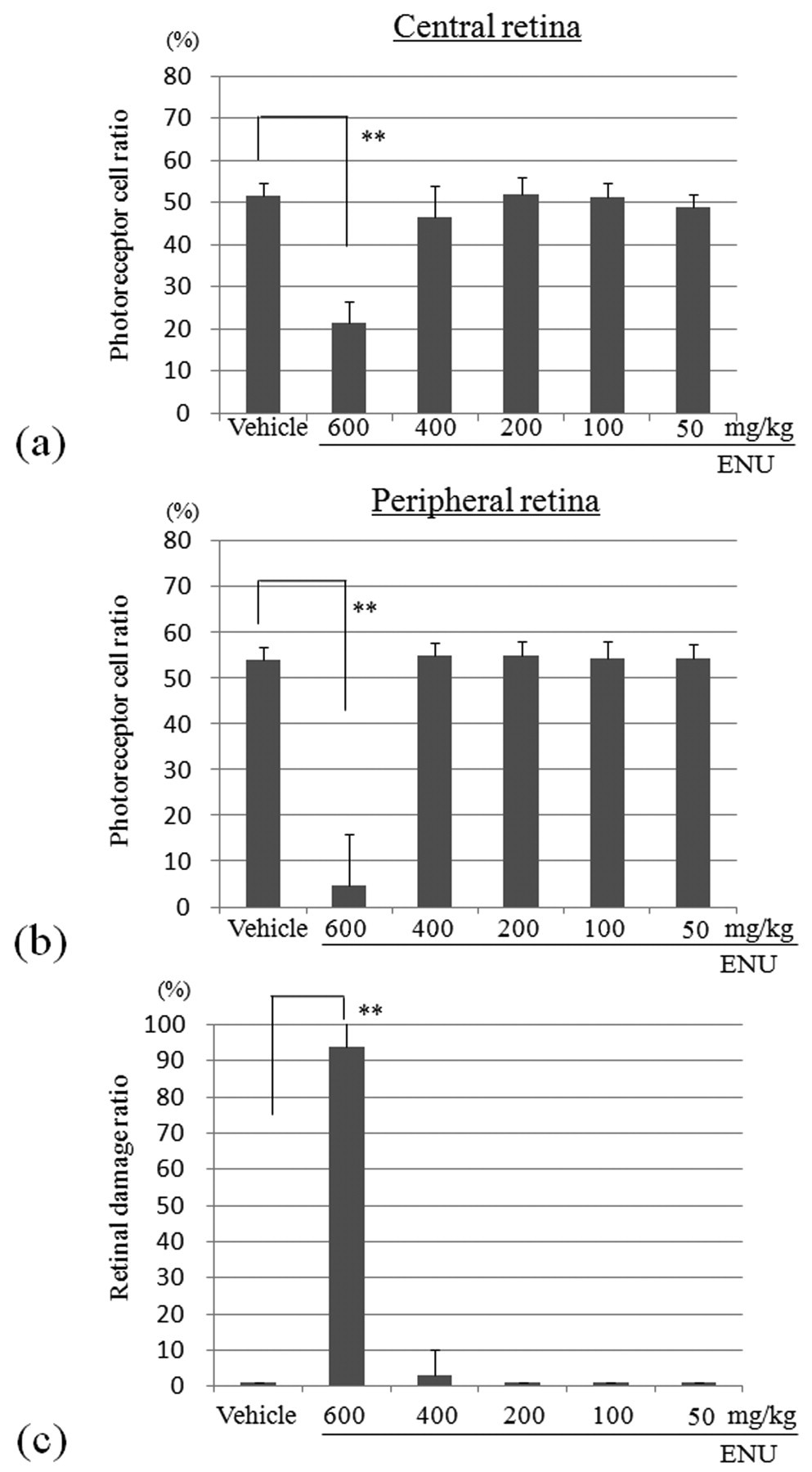
Photoreceptor cell ratio in the central retina (a) and peripheral retina (b) 7 days after a single intraperitoneal (ip) injection of ENU into adult mice. Mice treated with 600 mg/kg ENU had a statistically significant decrease in their photoreceptor ratio at the central and peripheral retina, as compared to vehicle-treated controls. The index was calculated as [(outer retinal thickness/total retinal thickness) × 100]. (c) Retinal damage ratio in ENU-treated adult mice. The index was evaluated as [(length of retina composed of less than four photoreceptor cells/whole retinal length) × 100]. The mean ± SE of the 5 mice in each treatment group is shown. **p < .01.
To evaluate the degree of disease progression, the retinal damage ratio was compared among the groups (Figure 2c). The 600 mg/kg ENU-induced damage covered almost the entire retina (94%), while it covered only 3% in the 400 mg/kg ENU group and 0% in the 200, 100, and 50 mg/kg ENU-treated groups (not significant). As anticipated, the ratio in vehicle-treated mice was 0%.
In the other organs examined, the most striking features were the dose-dependent suppression of hematopoiesis and the atrophy in the thymus and spleen of the 400 and 600 mg/kg ENU-treated groups (data not shown).
Morphological and Morphometric Analyses of Sequential Changes in Retinal Damage
The first evidence of retinal degeneration in mice treated with 600 mg/kg ENU was condensation and fragmentation of some photoreceptor cell nuclei that occurred 12 hr after treatment. The disruption and loss of the photoreceptor cell nuclei progressed at 24 hr after treatment, and widespread destruction of photoreceptor segments had occurred at 72 hr (Figure 3 ). Then the photoreceptor cell nuclei were almost lost in both the central and peripheral retina 7 days after treatment. As a result, the thickness of the total retina and the outer retina of ENU-treated mice decreased progressively (Figure 3). During the disease course, no inflammatory cell infiltration was seen in the retina.
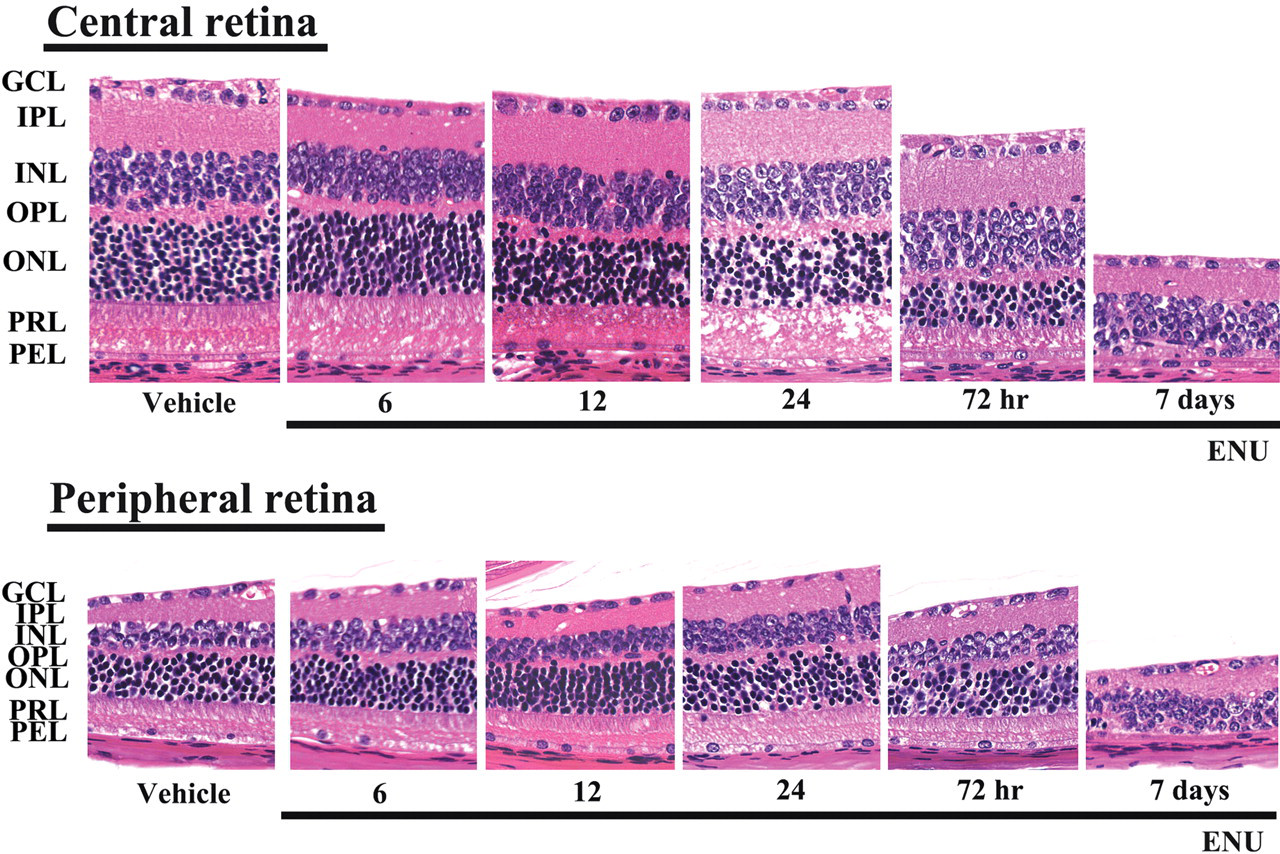
Sequential retinal changes in adult mice after a single intraperitoneal (ip) injection of 600 mg/kg ENU. Retinal cross sections were collected 6, 12, 24, and 72 hr and 7 days after ENU treatment. The retina of a saline-treated control mouse is also shown. GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; PRL, photoreceptor cell layer; and PEL, pigment epithelial cell layer. HE staining, ×400.
Aging Effect on ENU-Induced Retinal Damage
When 600 mg/kg ENU was administered to 25- to 34-week-old mice, retinal photoreceptor cell loss occurred in the central and peripheral retina, and the degree of retinal degeneration was similar to that observed in younger mice (Figure 4 ).
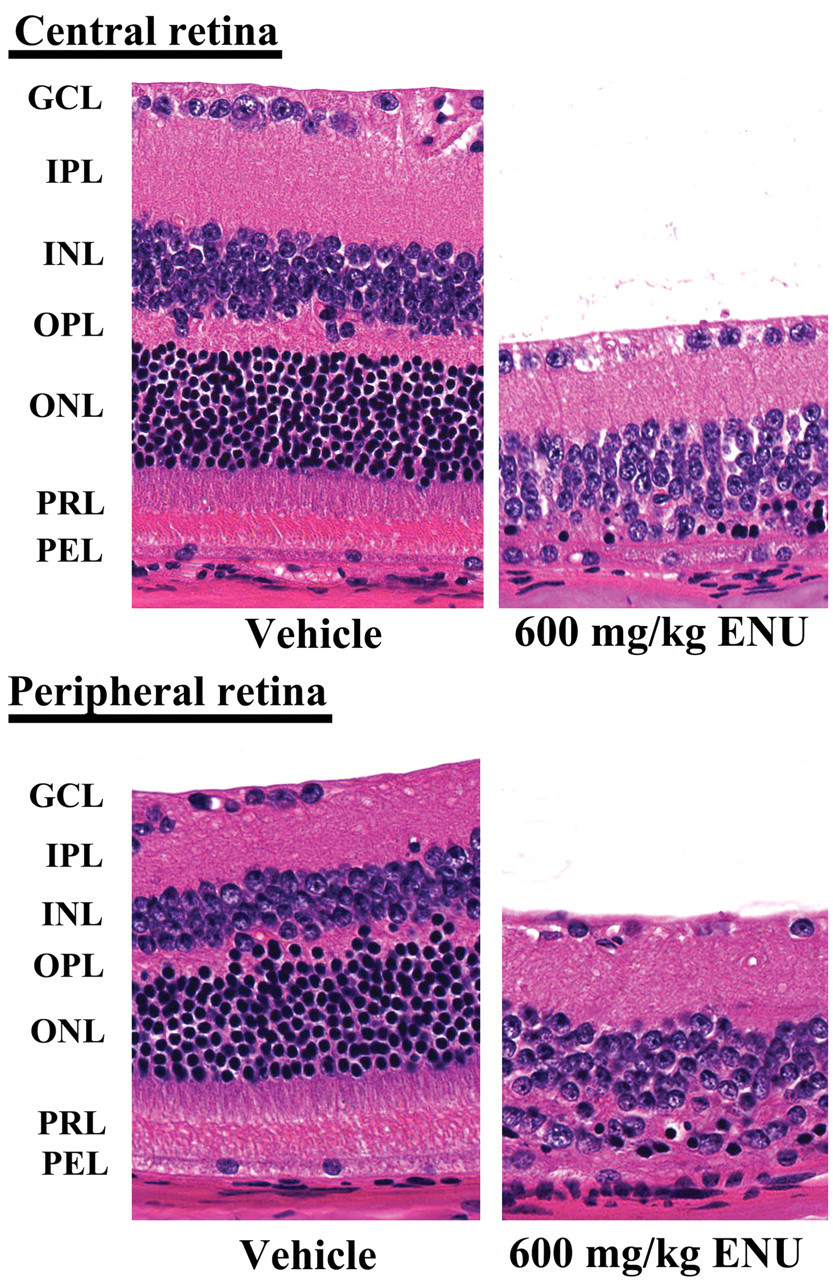
Retinal change 7 days after one intraperitoneal (ip) injection of 600 mg/kg ENU into mice at 34 weeks of age. The outer nuclear layer and photoreceptor layer almost disappeared in both the central and peripheral retina, as compared to the vehicle-treated control mouse. The remaining photoreceptor nuclei were densely stained, and their chromatins were clumped. GCL, ganglion cell layer; IPL, inner plexiform layer; INL, inner nuclear layer; OPL, outer plexiform layer; ONL, outer nuclear layer; PRL, photoreceptor cell layer; and PEL, pigment epithelial cell layer. HE staining, ×400.
Apoptotic Index
In both the central and peripheral retina of 600 mg/kg ENU-treated mice, the TUNEL signal appeared selectively in the photoreceptor cell nuclei, and many pyknotic photoreceptor cell nuclei at 12 and 24 hr showed TUNEL positivity without obvious nuclear destruction (Figure 5a ). The sequential changes in the apoptotic index at 0, 6, 12, 24, and 72 hr and 7 days were 0.3, 0.8, 14.1, 40.6, 51.8, and 25.3, respectively, in the central retina and 0.3, 1.1, 8.0, 31.3, 42.8, and 15.7, respectively, in the peripheral retina (Figure 5b).
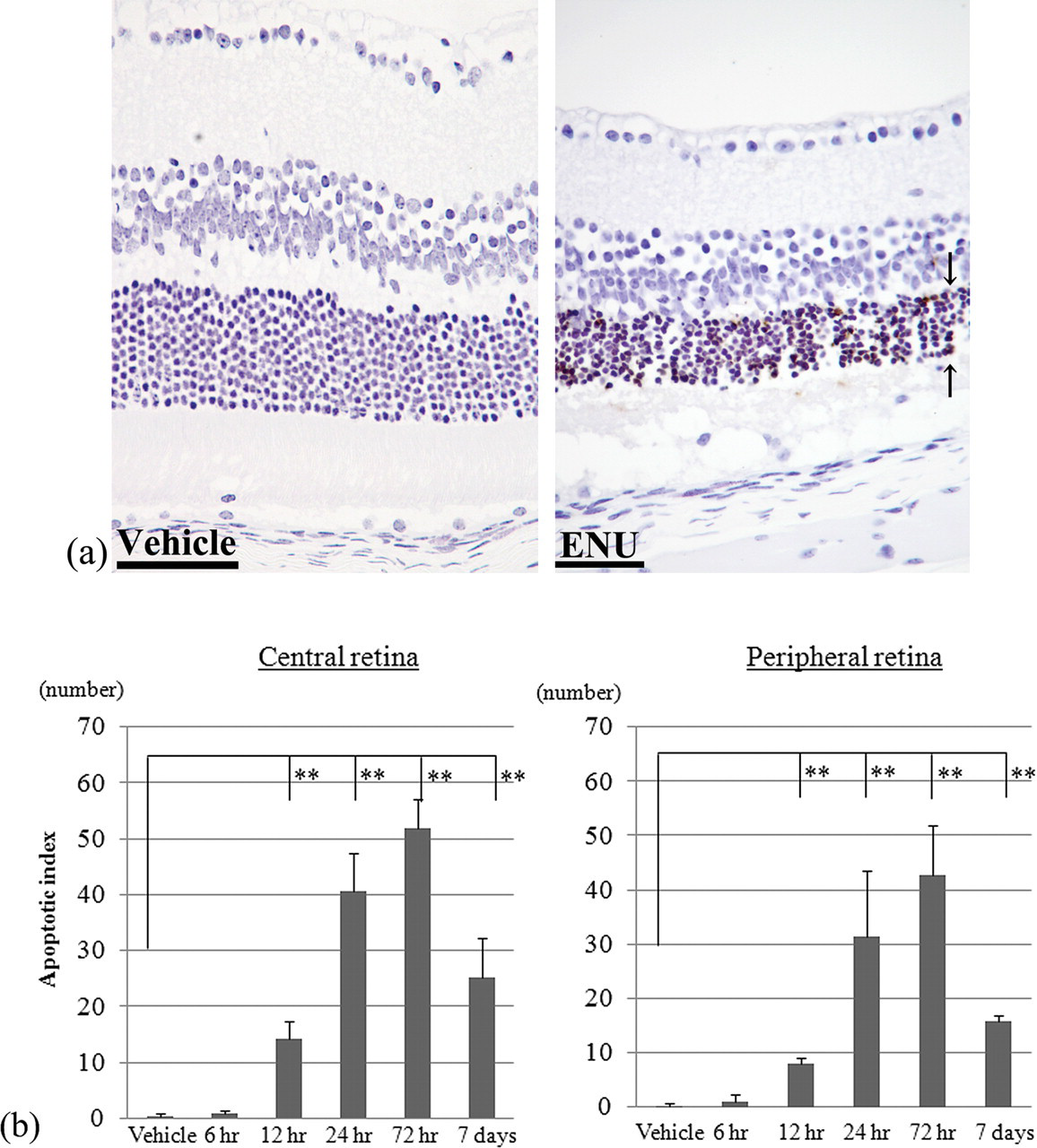
(a) Sequential changes in the expression of TUNEL- positive photoreceptor cells in the outer nuclear layer. The signals are seen in many photoreceptor cell nuclei (arrows). TUNEL staining, ×400. (b) The apoptotic index was evaluated in the central retina and peripheral retina, respectively. Mean ± SE, each bar represents 4 mice. TUNEL, terminal deoxynucleotidyl transferase (TdT)–mediated dUTP digoxigenin nick end-labeling. **p < .01.
Discussion
The present study provides new evidence of retinal changes in response to ENU. Retinal degeneration characterized by the loss of the outer nuclear layer and photoreceptor layer in the central and/or peripheral retina was detected in ENU-treated adult mice 7 days after a single ip injection of 400 mg/kg ENU or more. The outer nuclear layer and photoreceptor layer of ENU-treated mice decreased progressively due to photoreceptor cell apoptosis, which was characterized by the presence of TUNEL-positive nuclei restricted to photoreceptor cells. Photoreceptor cell apoptosis was evident at 12 hr and peaked at 72 hr after ENU treatment, followed by extensive photoreceptor cell loss at 7 days. Morphometrical indexes, such as the photoreceptor cell ratio in the central and peripheral retina and the retinal damage ratio, reveal the degree of retinal lesions induced by chemicals (Yoshizawa et al. 1999, 2009). In the present study, a morphometric analysis showed that photoreceptor cell damage occurred similarly in both the central and peripheral retina of mice treated with 600 mg/kg ENU, while 400 mg/kg ENU caused only low-magnitude damage in 1 out of 5 mice that was restricted to the central retina. Therefore, the central retina may be more sensitive to ENU-induced damage. In the 25- to 34-week-old mice treated with 600 mg/kg ENU, the retinal photoreceptor cell loss was similar to that of 7-week-old mice; thus, the age at exposure did not influence the ENU-induced retinal damage in adult mice.
Most inbred strains of mice cannot tolerate 400 mg/kg ENU administered by intraperitoneal injection, while BALB/c mice can tolerate this dose of ENU (Justice et al. 2000). In the present study, a single dose of 400 mg/kg ENU to female BALB/c mice caused hematopoietic toxicity and immunotoxicity but without mortality. ENU is a stem-cell mutagen, and it also affects hematopoietic stem cells and immune cells, and animals are highly susceptible to infection after injection of ENU (Justice et al. 2000). Thus, the cause of death in the 600 mg/kg dose group may be due to hematopoietic toxicity and immunotoxicity.
N-methyl-N-nitrosourea, which is a DNA-alkylating agent that is similar to ENU, causes DNA adduct formation in photoreceptor nuclei, followed by apoptosis via the down-regulation of Bcl-2, up-regulation of Bax, and activation of caspase families (Yoshizawa et al. 1999, 2009; Tsubura et al. 2010). In the present study, ENU induced an increased number of TUNEL-positive photoreceptor cells starting 12 hr after treatment and peaking 3 days after treatment. Further studies of the cascade of photoreceptor apoptosis are necessary to understand the detailed pathogenesis of ENU-induced retinal degeneration.
ENU is the most potent known mouse mutagen, and it can be used to generate point mutations throughout the mouse genome. The progeny of ENU-mutagenized male mice can be screened for autosomal dominant phenotypes, or they can be used to generate multigeneration pedigrees to screen for autosomal receive traits (Justice et al. 1999; Probst and Justice 2010). Various mouse models for human diseases have been generated by a systematic, large-scale, genome-wide phenotyping screen in the past decade (Wagner et al. 2010). Morphological defects in eyes, such as lens opacity, microphthalmia, corneal adhesion, corneal opacity, pink eye, iris anomalies, coloboma, and preretinal blood vessels (Baird et al. 2002; Favor and Neuhauser-Klaus 2000; Wagner et al. 2010), as well as retinal degeneration in the progeny (Maaswinkel, Mason, and Li 2003; Liu et al. 2010; Sato et al. 2010), have been reported. In addition, a single ip injection of 100 mg/kg ENU on day 14 of gestation or of 200 mg/kg ENU on day 3 after birth induces supraependymal cell clusters and rosette-like structures (dysplasia) in the retinas of fetal and neonatal rats, respectively (Maeda et al. 1985; Nagaishi 1982). The defect in the outer limiting membrane induced by the cytotoxic effect of ENU is an essential factor for the formation of these lesions. Few reports have been published concerning the effect of ENU on the retinal morphology of adult rodents, although ENU has been used as a carcinogen and mutagen in many animal toxicity and carcinogenesis studies. To the best of our knowledge, the present study is the first report on ENU-induced retinal injury in adult rodents.
In conclusion, retinal degeneration occurred in adult mice after a single intraperitoneal injection of 400 mg/kg ENU or more. The degrees and quantities of lesions were more severe in mice treated with 600 mg/kg ENU, which is the lethal dose in adult mice. ENU-induced retinal degeneration was related to photoreceptor apoptosis, which was more severe in the central retina than in the peripheral retina. Cell loss via apoptosis is increasingly recognized as a frequent component in the course of different human retinal diseases including retinitis pigmentosa, as well as in mutant mice models, and apoptosis has been considered as a final common pathway of photoreceptor cell death (Chang, Hao, and Wong 1993; Hartong, Berson, and Dryja 2006; Rivas and Vecino 2009; Sancho-Pelluz et al. 2008). Thus, the manipulation of factors that influence the apoptotic pathway may be of potential value for retinal disease therapy. Investigations of the detailed mechanisms of ENU-induced retinal degeneration are needed for a better understanding of the pathogenesis of retinal degeneration in humans.
Footnotes
Acknowledgments
The authors thank Ms. T. Akamatsu for her excellent technical assistance and Ms. A. Shudo for preparing the manuscript.
The authors declare that we have no competing financial interests. This research was supported in part by a Grant-in Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (22591954).