
Abstract
Propane-oxidizing bacteria in surface soils are often used to indicate the position of oil and gas reservoirs. As a potential replacement for the laborious traditional culture-dependent counting method, we applied real-time fluorescent quantitative polymerase chain reaction detection as a quick and accurate technology for quantification of propane-oxidizing bacteria. The propane monooxygenase gene was set as the target and the assay is based on SYBR Green I dye. The detection range was from 9.75 × 108 to 9.75 × 101 gene copies/µl, with the lowest detected concentration of 9.75 copies/µl. All coefficient of variation values of the threshold cycle in the reproducibility test were better than 1%. The technique showed good sensitivity, specificity, and reproducibility. We also quantified the propane-oxidizing bacteria in soils from three vertical 250 cm profiles collected from an oil field, a gas field, and a nonoil gas field using the established technique. The results indicated that the presence of propane monooxygenase A genes in soils can indicate an oil or gas reservoir. Therefore, this technique can satisfy the requirements for microbial exploration of oil and gas.
Keywords
Introduction
With the extensive exploration of oil and gas reservoir, the distribution of remaining oil and gas resources increasingly dispersed, and reservoirs are getting smaller and most of the reservoirs belong to atectonic reservoirs. Therefore, it is difficult and costly to explore the reservoir with the geophysical and geochemistry exploration only (Yuan, 2011). Gaseous hydrocarbons present in subsurface reservoirs can migrate to the surface soil through upward diffusion and effusion in response to the formation pressure (Saunders et al., 1999). Some microbes in soil can exclusively use these hydrocarbons as the carbon source for their metabolic activities and growth (Mogilewskii, 1938). As a result, there is a positive correlation between hydrocarbon concentration and hydrocarbon-oxidizing bacteria in the soil (Hitzman et al., 2002). These light hydrocarbon-oxidizing bacteria have been exploited for petroleum exploration for a long time (Rasheed et al., 2011). The microbial prospecting methods can give us the cheap and effective early exploration to reduce drilling risks. The methods can also grade the hydrocarbon contents in various geologic structures. Especially, it is of great significance in the prediction of subtle and deep reservoir (Liu et al., 2014).
Methane is the dominant component of light hydrocarbons in oil and therefore methane-oxidizing bacteria have been intensively investigated (Xu et al., 2013). However, in addition to thermogenic methane, biogenic methane originates from the decomposition of organic matter by methanogens in anaerobic environments and can occur under appropriate substrate and environmental conditions regardless of the presence of oil or gas reservoirs (Schoell, 1988). As a result of the multifactorial nature of methane formation, methanotrophs alone are not reliable indicators of petroleum reservoirs (Brisbane and Ladd, 1965). In contrast, propane, another important light hydrocarbon, is appropriate for petroleum reservoir prospecting because biologically produced propane is much less prevalent than propane originating from seepage from geological sources (Ashraf et al., 1994).
There are two different traditional methods of determining the abundance of gaseous hydrocarbon-oxidizing bacteria: the plate method and the maximum probability number method (including the culture flask test) (Yi et al., 2006). The current two microbiological techniques used to explore for oil and gas, the Microbial Oil Survey Technique and Microbial Prospecting for Oil and Gas, are examples of a plate method and a maximum probability number method, respectively (Chan, 2011). Both methods can be called culture-dependent microbial approaches, which are laborious and time consuming and cannot represent the uncultured microorganisms that account for more than 99% of the microorganisms naturally occurring in the environment (Hugenholtz et al., 1998).
The development of a technology that detects a population of bacteria without culturing is widely preferred. Molecular techniques offer speed and specificity that culturing cannot. Molecular techniques related to functional genes have been proven to be effective as a basis for understanding the microbial diversity in environmental communities (Anderson and Cairney, 2004). Methods to indicate the presence of propane-oxidizing microorganisms based on polymerase chain reaction (PCR) technology have been developed (Chan, 2011). However, there is currently no culture-independent quantification method for propane-oxidizing microorganisms. Fluorescent quantitative real-time PCR (RT-PCR), an important molecular technique, can quantify the specific functional genes from environmental samples within a matter of hours (Bustin et al., 2005). Molecular techniques, including RT-PCR, which can detect methanotrophic bacteria are currently available and have been intensively used in petroleum reservoir prospection (Miqueletto et al., 2011; Xu et al., 2013; Zhang et al., 2010). However, as an important indicator of oil and gas reservoirs, the quantification of propane-oxidizing bacteria has not been studied in detail.
Bacterial propane oxidation is catalyzed by a reduced nicotinamide adenine dinucleotide-dependent multicomponent enzyme system belonging to the family of dinuclear-iron oxygenases. There are three components in this dinuclear-iron-containing monooxygenases, including a hydroxylase, a reductase, and a coupling protein. Hydroxylation of propane occurs at a dinuclear iron of the large subunit (propane monooxygenase A (prmA)) accompanied by the small subunit (propane monooxygenase C). The propane monooxygenase B and propane monooxygenase D correspond to the reductase and coupling protein, respectively. These enzymes are responsible for the first reaction that can oxidize the propane to propanol (Kotani et al., 2003). Therefore, prmA gene is the key gene that encodes the propane oxidation enzyme and cannot be knocked out when metabolizing the propane (Sharp et al., 2010). In some bacteria, the products of these prmA genes also involve in other gaseous n-alkane, some halohydrocarbon, and ether oxidation (Kotani et al., 2006; Steffan et al., 2003; Vanderberg and Perry, 1994). Generally speaking, the origin of the other gaseous n-alkane is also oil reservoir in natural condition (Saadoun and Al-Ghazawi, 2005). The halohydrocarbon and others are closely related to human activities and may migrate into soils, which can be avoided when sampling. From the above, we may find that the quantification of prmA gene may ravel the relative abundance of propane-oxidizing bacteria in soils and has the potential for oil and gas exploration.
In this study, we developed the RT-PCR technique for the quantification of propane-oxidizing bacteria based on the prmA gene. We also evaluated the accuracy, sensitivity, reproducibility, and specificity of the technique. Furthermore, soils collected from an oil field, a gas field, and a nonoil gas field were used to examine whether the technique could be used in oil and gas exploration.
Materials and methods
Sample collection and DNA preparation
Gas field soil samples were taken from the Xiaoquan-Xinchang gas exploration area in western Sichuan Province, China. This gas field is a composite field that was composed of multiple Jurassic secondary gas pools and has a shallow burial depth (Ren and Xia, 2006). Oil field soil samples were taken from the Yangxin oil exploration area in the Huabei oil field, China. The site is located above an anticlinal structural reservoir, which is an anticlinal structural trap that was formed by the gravitational effects and the differential compaction of Paleogene Shahejie formation sediments that were deposited above the early formed horst. The nonoil gas field samples were collected from a farmland in Zhengding County, Hebei Province, China. It is a contact zone of the Shanxi platform and the Bohai depression. The sampling sites are pinpointed in Figure 1.
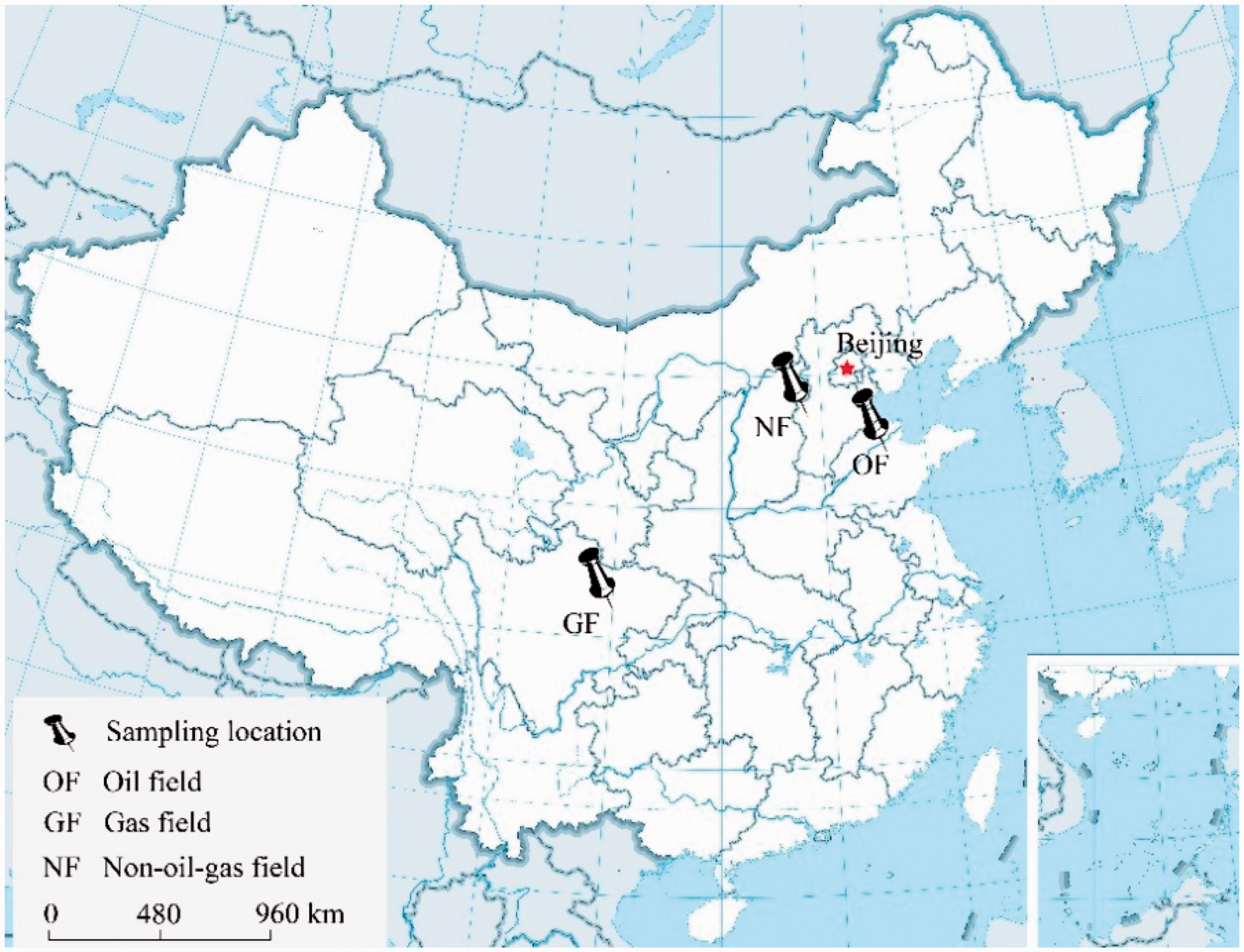
The locations of the sampling sites.
In the oil and gas fields, a vertical profile in the vadose zone was collected, 50 m from the oil or gas well to avoid disturbance during well completion. A vadose zone vertical profile was also chosen in the nonoil gas field in an area identified as least disturbed by human activities. Eight soil samples retrieved from depths of 0.05, 0.2, 0.5, 1.0, 1.5, 2.0, and 2.5 m were collected from each profile.
When sampling, the soil cores were initially taken out using Luoyang shovel which is a special hand auger and is used for drilling soil profiles and sampling the soil. Subsequently, a part of the soil samples was collected from the core center using a sterile knife and placed in aluminum boxes previously sterilized at high temperatures. Collected samples were immediately stored in an incubator with an ice pack and then transferred to the laboratory and stored at −70°C prior to extraction of the genomic DNA within two days. Another subsample of the soils was stored in plastic automatic sealing bags for analysis of physical and chemical characteristics. In addition, the soils at 150 cm depth were chosen for light gaseous hydrocarbon analyses. These soil samples were placed into a 1000 ml plastic bottle, which was prefilled with saturated brine and then sealed with a screw cap.
Genomic DNA from approximately 0.5 g of fresh soil for each of the samples was extracted using the PowerSoil® DNA Isolation Kit (Mo Bio Laboratories, USA) according to the manufacturer’s instructions. The obtained DNA was quantified with a Biodropsis BD-2000 spectrophotometer. The DNA extracts were stored in Tris–EDTA buffer (pH 7.0) at −20°C for further analysis.
The soil temperature was measured using a mercurial thermometer and the pH was measured using a portable pH meter (Leici PHB-4, China). The soil water content was determined by drying soil overnight at 105°C. The soil lithology was determined according to the Standard Test Method for Particle-Size Analysis of Soils (ASTM D, 2009). The salinity was estimated based on the conductivity of the soil–water system (Lu, 2000). The light gaseous hydrocarbons adsorbed in soil particles were extracted by heating the bottle and analyzing on a gas chromatograph (Lakshmi et al., 2013) at the National Research Center for Geoanalysis, Chinese Academy of Geology Sciences.
Primer design and standard plasmid preparation
The forward and reverse primers of 955F25 (TGG CAC CGG TGG ATC TAC GAC T) and 1517R22 (GCG CGA TCA GCG TCT TGC CGT C) were designed using conserved nucleotide sequences in genes encoding the large hydroxylase subunit of propane monooxygenases, prmA (Chan, 2011).
The genomic DNA from the oil field at 50 cm soil depth was chosen as the template for the PCR procedure. The PCR was performed in an Eppendorf, 5333 PCR Thermocycle Instrument (Eppendorf AG) and followed the optimized protocol for 50 µl reactions (Chan, 2011). The PCR reaction products were examined by 1.0% agarose gel electrophoresis (DYY-6C, Beijing Liuyi, China) and visualized with the Gel Documentation and Analysis System (WD-9413B, Beijing Liuyi, China). If the PCR products were verified as the target fragment, prmA, then they were purified using the Gel Extraction Kit (Promega). The purified DNA fragments were ligated into pGEM-T Easy vectors and then the vectors were transformed into Escherichia coli DH5α and incubated at 37°C. The positive clones were picked for sequencing. The clones that had the same sequence were collectively called an operational taxonomic unit (OTU). The clone of the OTU with the most members was chosen as the target clone and was used to provide the standard plasmid for quantitative PCR. The standard plasmid was extracted using the High-purity Plasmid Mini-preparation Kit (spin-column) (Bioteke, DP1001) and the extracted plasmid was quantified by Biodropsis BD-2000 spectrophotometer and then stored at −20°C for further use. The copy numbers of the prmA gene were calculated from the DNA sequence, the number of inserted DNA fragments, and the extracted plasmid concentration.
Optimization of the RT-PCR reaction system
First, according to the procedures and reaction system of the SYBR® Premix Ex Taq™ kit, the prmA gene fragments of soil DNA were amplified by PCR reactions with primers 955F25 and 1517R22 in the StepOne Real-Time PCR System (Life Technologies). The amplification curve and the melting curve were then obtained. The primer concentration and annealing temperature were adjusted to optimize the RT-PCR reaction system based on the curves. Finally, the optimal reaction system and amplification procedures were determined as follows:
The total volume of the reaction mixture system was 20 µl, including 2× SYBR Premix Ex Taq™ (10 µl), 10 µM PCR Forward Primer (0.2 µl), 10 µM PCR Reverse Primer (0.2 µl), 50× ROX Reference Dye (0.4 µl), DNA (2 µl), and dH2O (7.6 µl). The mixtures were processed at 94°C for 30 s, followed by 40 cycles of 5 s at 94°C, 30 s at 66°C, and 60 s at 72°C and then a read step at 88°C for 15 s. Subsequently, in the melt stage, the temperature was held for 15 s at 95°C and for 60 s at 60°C, and then the temperature was gradually increased to 95°C (for every 0.3°C increase, the fluorescence intensity was read for 15 s).
Sensitivity evaluation
Tenfold serial dilutions of standard plasmid DNA, acting as templates in duplicate, were used as a RT-PCR assay to prepare a standard curve for the enumeration of propane-oxidizing bacteria. The relationship between prmA gene concentrations and the threshold cycle (CT) values was analyzed to evaluate the sensitivity of detection of the method.
Repeatability evaluation
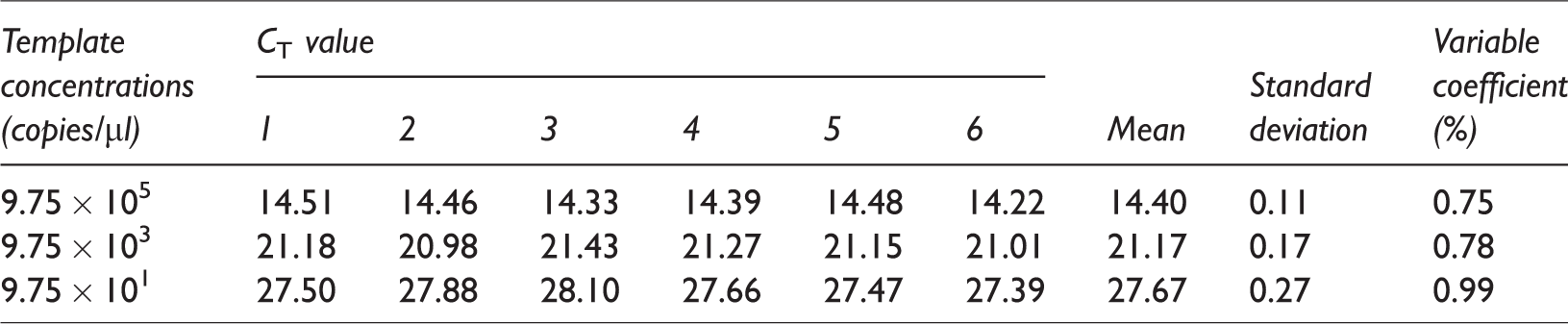
The standard plasmid DNA was diluted to three concentrations from high to low concentrations (9.75 × 105, 9.75 × 103, and 9.75 × 101 copies/µl). Each of the three concentrations was measured six times. The standard deviation and coefficient of variation of the CT value and the prmA concentrations were then calculated.
Specificity evaluation
The specificities of the technique were tested with template DNA from strains not containing propane monooxygenase from our laboratory culture collection, which include aniline-degrading bacteria, quinoline-degrading bacteria, ethyl sulfonic acid-degrading bacteria, and oil-degrading bacteria. Plasmid DNA at the concentration of 9.75 copies/µl and sterile water were set as the positive and negative controls, respectively. Subsequently, the CT and ΔRn (the magnitude of the fluorescence signal generated during the PCR at each time point) and the shape of the amplification and melting curves were used to evaluate the specificity of the assay.
Further, the diversity of propane-oxidizing bacteria in the RT-PCR product was examined and the phylogenetic tree was generated using clone library analysis (Jones et al., 2009).
Quantification of the prmA gene in soil samples
The purified DNA fragments from soil samples collected from the oil field, gas field, and nonoil gas field were used as template to perform the RT-PCR amplification to quantify the prmA gene. After the prmA gene concentration data were obtained, the technique was evaluated as to whether it could be used in oil and gas exploration. All samples were processed in duplicate.
To determine the relative abundance of prmA in all bacteria, the concentrations of 16S ribosomal RNA (16S rRNA) genes in soils were also quantified using the RT-PCR technique as a reference (Shu et al., 2016). The ratio of prmA to 16S rRNA, which might indicate the activity of propane-oxidizing bacteria, was then calculated.
Results
Physical and chemical characteristics
The results of physical and chemical characteristics of the soil samples are shown in Table 1. All samples have similar salt content ranging from 0.02 to 0.07%. The gas field samples had the highest temperature by a small margin. The oil field samples had higher pH values (8.92–9.38) than the gas field samples (7.61–8.08). The surface samples had lower pH values than samples at depth in both fields. The moisture content of gas field samples was nearly double that of the oil field samples, which might be because of the climate (Chen et al., 2005). The surface soil samples (0–50 cm) in the gas field are silty light clay while in the oil field they are silty light loam. At the depth of about 200 cm, the gas field samples are medium sand while the oil field samples are sandy silt. In general, the Yangxin oil field soil is characterized by weak alkaline pH, low moisture, and low temperature.
The physical and chemical characteristics of the soil samples.
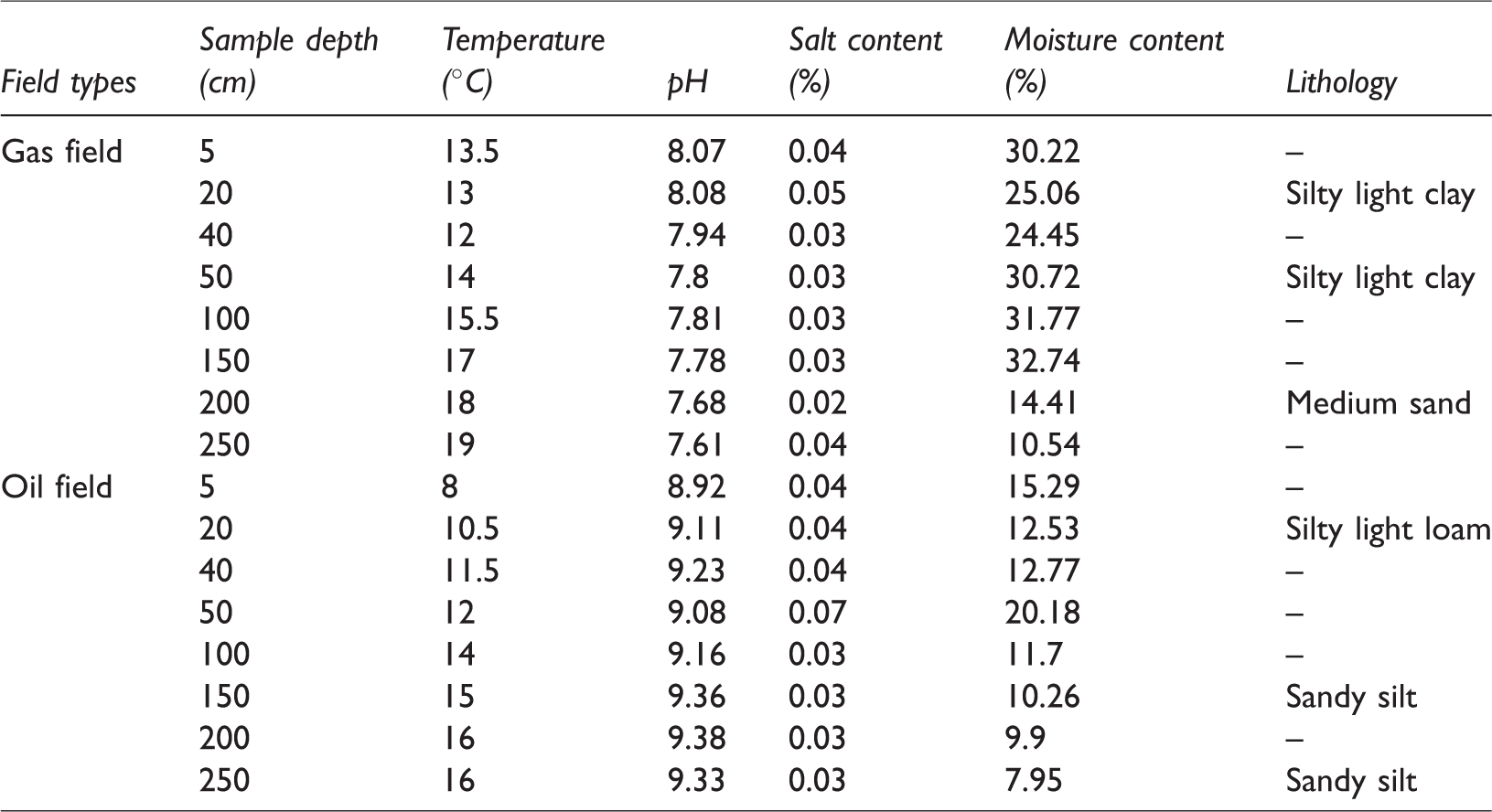
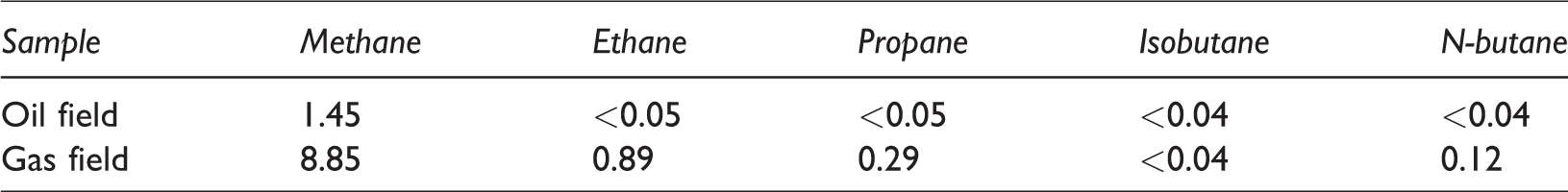
Table 2 shows the concentrations of light gaseous hydrocarbons adsorbed in soil particles at 150 cm depth. We found that the soil in the gas fields had more light gaseous hydrocarbons than those in the oil field. This is because the gas reservoir might effuse more propane than the oil reservoir and the surface clay soil might retard the gas seeping out of the soil in the gas field.
The concentrations of C1–C4 hydrocarbons in soil samples’ particles µl/l.
Standard plasmid preparation
The PCR reaction was performed using the template DNA from the oil field soil and the product was determined to be the target fragment, the prmA gene (see Supplementary Material). After the PCR product was purified, ligated, transformed, and cultured, the optimal clone was selected, amplified, and the plasmids were extracted as the standard plasmid (Supplementary Figs. 1–3). The concentration of the plasmid was quantified to be 39 ng/µl, which is equivalent to 9.75 × 109 copies/µl.
Sensitivity evaluation
When the concentrations of standard plasmid ranged from 9.75 × 108 to 9.75 × 101 copies/µl, the prmA gene amplified well (Figure 2), and the melting curves showed specific peaks. The propane-oxidizing bacteria detection standard curve, whose vertical axis is CT values and horizontal axis is standard plasmid concentrations in denary logarithm form, was established (Supplementary Fig. S4). The regression equation of the curve was as follows
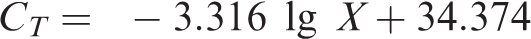
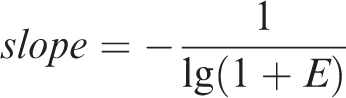
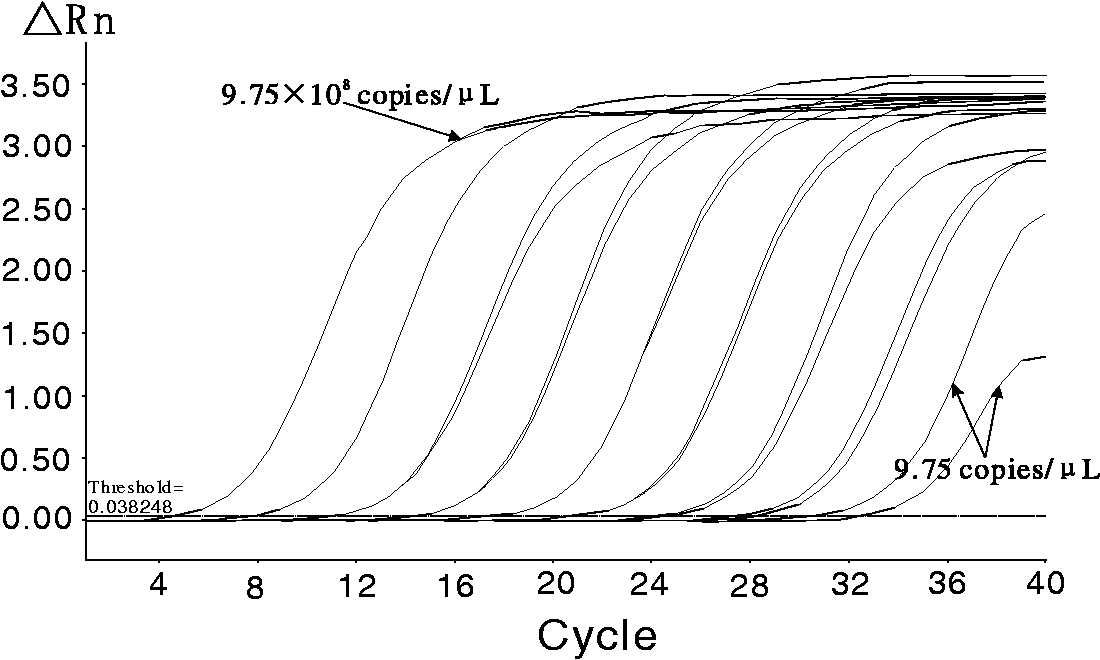
Amplification curves of standard plasmid with prmA gene.
As a result, the measurement range of this method was shown to be from 9.75 × 108 to 9.75 × 101 copies/µl.
We also found that the amplification curve could still be detected even if the concentration of the template was only 9.75 copies/µl. Therefore, the limit of detection of the method was 9.75 copies/µl.
Repeatability evaluation
The CT values of the repeatability evaluation experiment are shown in Table 3, and the list of concentrations is given in Table 4. The standard deviation and coefficient of variation of the CT values and measured concentrations were calculated and are also presented in Tables 3 and 4. With the decrease in template concentration, both the standard deviation and coefficient of variation of CT values increased gradually, but the coefficient of variation values were less than 1%. For the measured concentrations, the standard deviation decreased and the coefficient of variation increased as the template concentration decreased. However, the coefficient of variation values were less than 20%. The errors of the coefficient of variation values are all within the allowable range, which implies that the repeatability of the method was satisfactory regardless of the prmA gene concentrations. Therefore, for this method, the maximum error was identified as 20 and 1% for the prmA gene concentrations and CT values, respectively.
The CT values in repeatability evaluation.
The template concentrations measured in repeatability evaluation.

Specificity evaluation
The CT values of various bacteria in the specificity evaluation experiment are shown in Figure 3. We found that the CT values of the negative control and the non-propane-oxidizing bacteria samples were all more than 27, which is beyond the scope of determination of the standard curve and indicated that these samples had few or no prmA genes. Moreover, irregular amplification curves and nonspecific melting curves also indicated the same result (data not presented).
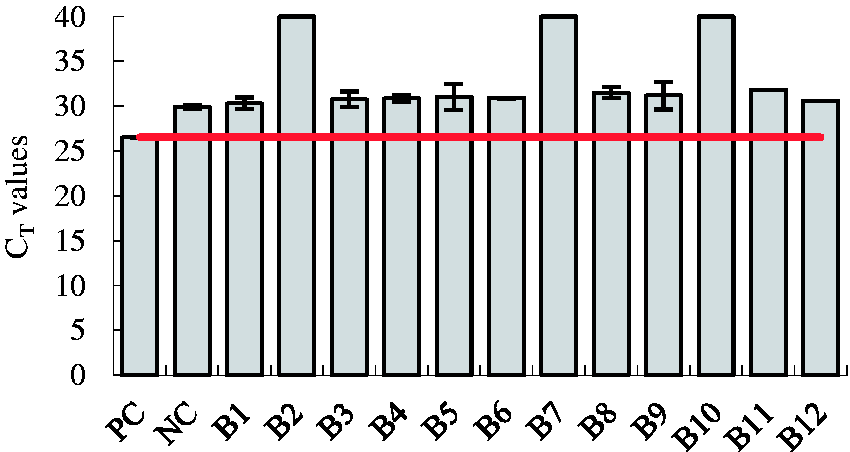
The CT values of non-propane-oxidizing bacteria. PC represents the positive control, NC represents the negative control and B1–B12 represents the 12 different non-propane-oxidizing bacteria. The line is the limit of determination of the CT value.
In addition, we examined the diversity of propane-oxidizing bacteria in the RT-PCR product using clone library analysis of the soil sample collected at 50 cm soil depth in the Yangxin oil field (Figure 4). There were 145 clones in the library, all of which contained the propane monooxygenase gene. Most of these clones have the same sequence as BYX-1, whose sequence is similar to an uncultured soil bacterium, clone SL01 (JX045657.1), propane monooxygenase gene, according to the NCBI database. The percentage identity value was 90%. The SL01 clone was obtained during an investigation of the activity and diversity of the microbial community in the presence of light hydrocarbons. The BYX-13, BYX-53, BYX-66 clones have similar sequences to Rhodococcus sp. RR1 (HM209445.1), Pseudonocardia sp. TY-7 (AB250941.1) and Gordonia sp. TY-5 (AB112920.1), respectively, and were all identified during research focusing on the metabolism of propane. These results indicate that the RT-PCR method is specific for the quantification of propane-oxidizing bacteria.
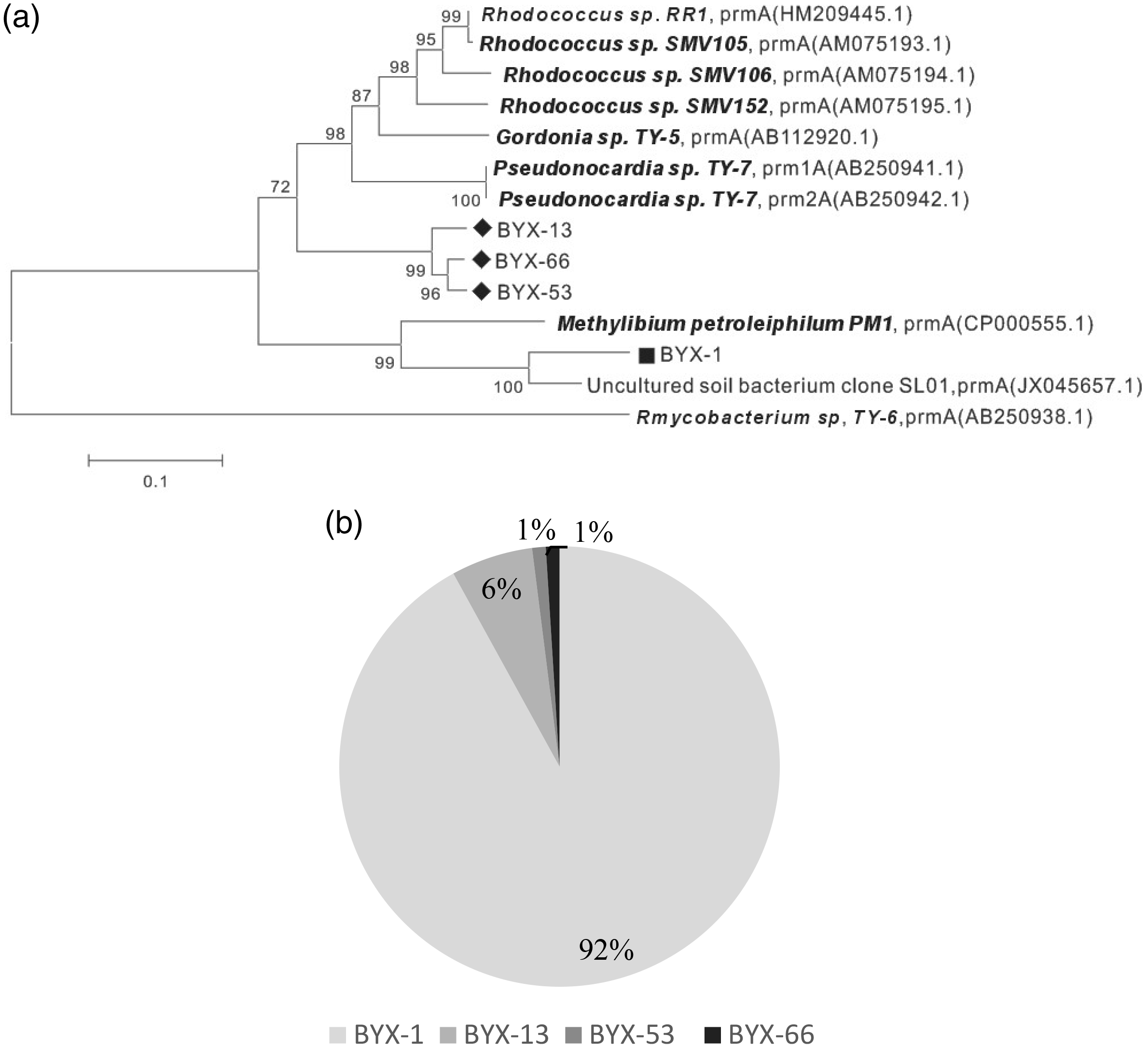
The diversity of bacteria in the RT-PCR products amplified from Yangxin oil field soil. Part a is the phylogenetic tree constructed through the clone library. Part b indicates the percentage of each clone that has the same sequences in the clone library. BYX-1, BYX-13, BYX-53, and BYX-66 are clones that have different sequences in the clone library. prmA: propane monooxygenase A.
Quantification of the prmA gene content in various soil samples
The CT values of various soil samples from the oil field and nonoil gas field are presented in Figure 5. The nonoil gas field samples, except for the sample collected at 5 cm depth, had CT values greater than 27, which indicates that the soils below 5 cm have no prmA genes above the detection limit and thus are unlikely to contain propane-oxidizing bacteria. This in turn reveals that there are no oil reservoirs beneath the sample field, which is consistent with reality. However, the oil field soil samples had CT values less than 27, indicating that the soils in the oil field contain considerable amounts of prmA gene and are thus rich in propane-oxidizing bacteria. This reveals that there are potential oil reservoirs beneath the sample field, which is also consistent with reality.
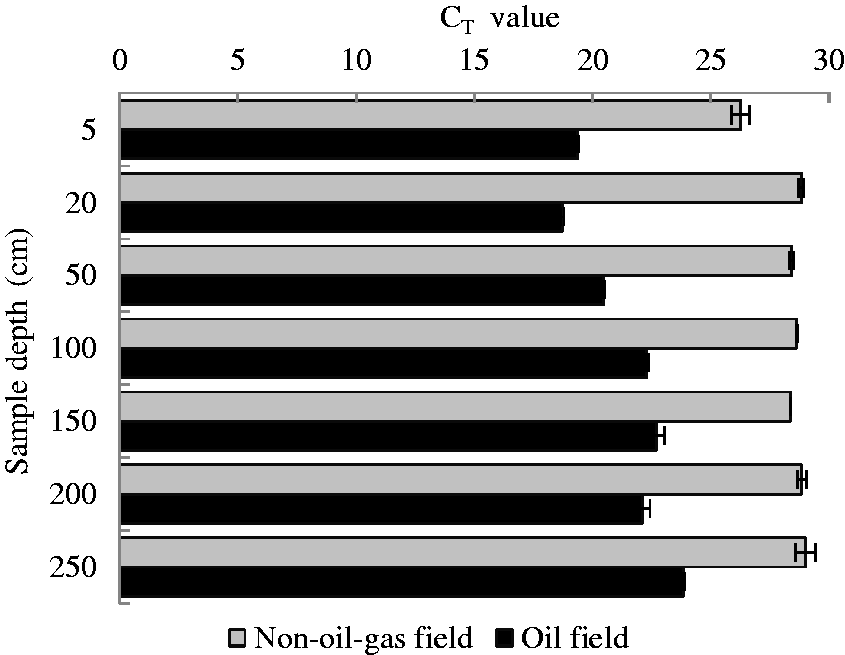
The CT values of soil samples from the oil field and the nonoil gas field.
According to the above comparison and analysis, we found that the prmA RT-PCR method has the potential for oil reservoir prospecting. Moreover, we also studied whether the method could be used in gas reservoir prospection. The soil samples were collected from the gas field and evaluated according to the test procedure. The prmA gene concentrations of the oil field and gas field samples are illustrated in Figure 6. Although gas field samples had lower concentrations of the prmA genes than the oil field samples, the levels of prmA genes detected in gas field samples were markedly different from those in the nonoil gas field, indicating that gas fields could also be detected using this method.
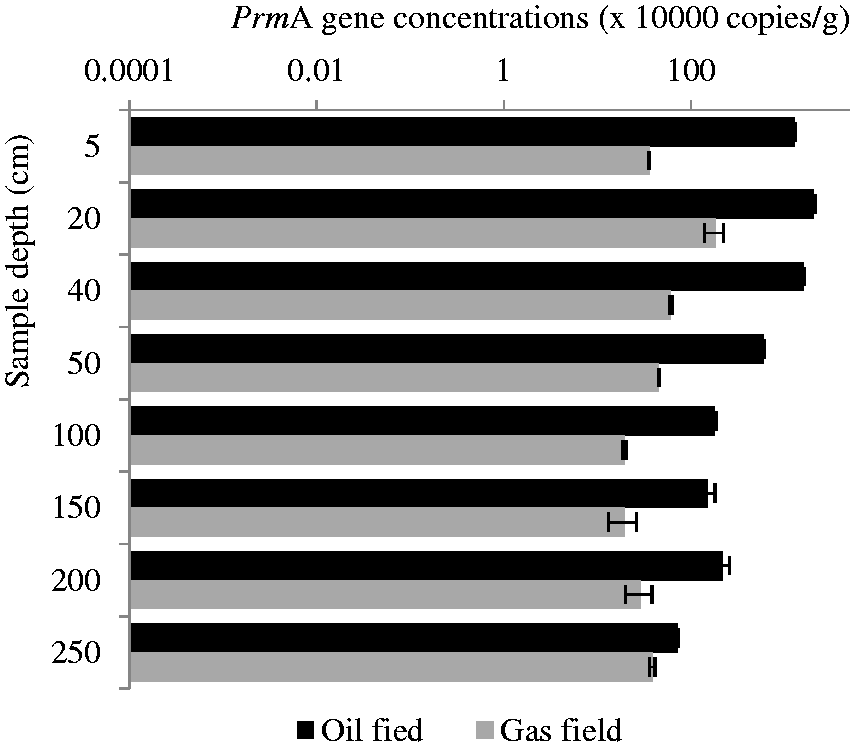
prmA gene concentrations of soil samples from the oil field and the gas field. prmA: propane monooxygenase A.
The ratio of the prmA to 16S rRNA gene concentrations of the soils sampled in both oil and gas fields is shown in Figure 7. It is clear that samples from the gas field have low relative abundance of the prmA gene, which indicates that propane-oxidizing bacteria in this area have lower activity compared with other bacteria.
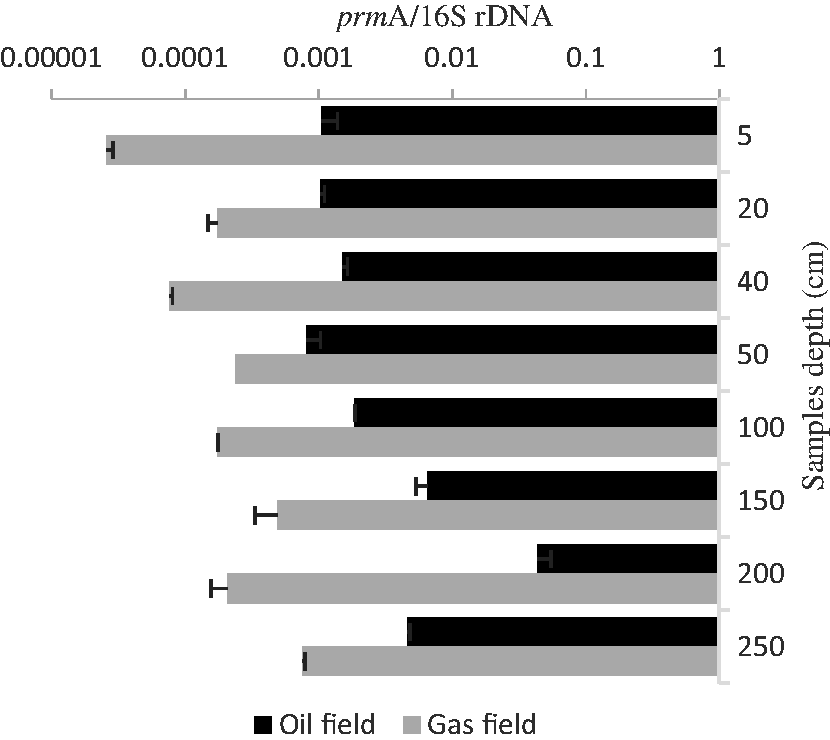
The ratio of prmA gene to 16S rRNA gene concentrations in the oil and gas field samples. prmA: propane monooxygenase A; 16S rRNA: 16S ribosomal RNA.
Discussion
The site selected in the Yangxin oil field is located above an anticlinal structural reservoir, which is an anticlinal structural trap that was formed by the gravitational effects and the differential compaction of Paleogene Shahejie formation sediments that were deposited above the early formed horst. The trap has the advantage of allowing the easy transportation, aggregation, and preservation of the oil and gas around it. The underground oil reservoir is 1390–2570 m beneath the surface. Furthermore, mature oil is predominantly distributed in the oil reservoir in this area (Yang et al., 2003). Mature oil has the characteristics of low density and viscosity, which means that the short-chain hydrocarbon content is high and light hydrocarbon gas can easily escape into the surface soils. Therefore, we detected relatively high concentrations of the prmA gene in this field.
The gas field from which the samples were collected is called the Xiaoquan-Xinchang gas field, which is composed of multiple secondary gas reservoirs in the Jurassic system. The Xiaoquan-Xinchang structure is an anticline, which formed a northeast-east-striking Palaeohigh belt. This belt is the migration zone for natural gas in the subsidence formation. Moreover, there are good conditions for the formation of structural-lithologic composite traps, producing the optimal conditions for the formation of a productive gas field in the area (Ren and Xia, 2006). From the field sample, we detected high concentrations of propane according to the headspace gas chromatography of soil samples (Table 2). However, the gas field had lower concentrations of the prmA gene than the oil field. This led us to examine the possible causes for the finding of high propane but low prmA gene prevalence.
The ratio of functional gene to 16S rRNA genes might solve the problem. 16S rRNA genes are present in all bacteria, while the prmA gene is only present in propane-oxidizing bacteria. The ratio of prmA:16S rRNA genes might indicate the activity of propane-oxidizing bacteria. It is clear that samples from the gas field have low relative abundance of the prmA gene (Figure 7), which indicates that propane-oxidizing bacteria in this area have lower activity compared with other bacteria, such as methane-oxidizing bacteria. These bacteria are likely to have the competitive advantage because of the optimal growth environment for them but suboptimal conditions for propane-oxidizing bacteria. The temperature, pH, nutrient content, moisture, grain size, and other soil characteristics affect the growth of microorganisms (Roszak and Colwell, 1987). The optimal conditions for most propane-oxidizing bacteria are mesophilic temperature, neutral pH, and aerobic environment (Ashraf et al., 1994), which is consistent with the overwhelming majority of microorganisms. However, the pH, moisture, and temperature might influence the growth of propane-oxidizing bacteria less than the other microorganisms (Levin and Bergstrom, 2000). As a result, there are high prmA gene concentrations in the Yangxin oil field, where the soil was characterized as weakly alkaline, low moisture, and low temperature. Moreover, the pores in silt and silty loam allow free air and moisture exchange with the environment, which is optimal for the growth of propane-oxidizing bacteria. In contrast, the clay present at the Xinchang gas field would retard aeration of the underlying soil and result in reduced numbers of propane-oxidizing bacteria. In addition, the clay layer might reduce the transfer of propane to the atmosphere, resulting in the accumulation of propane within the soil.
The geological background of the nonoil gas field at the site indicated that there were no oil or gas reserves and covering strata. As a result, all samples in the 2.5 m profile collected in this area, except for the 5 cm deep soil, had no prmA gene detected, which is consistent with the geological conditions. The detection of prmA genes in the 5 cm deep soil sample is probably the result of human activity above the surface soil. It has been reported that propane-oxidizing bacteria can also degrade several other types of organic matter, such as hydrohalocarbons, trichloroethylene, and tert-butyl ether components (Steffan et al., 1997, 2003; Streger et al., 1999; Wackett et al., 1989). These materials are often used by humans as chemical materials and might migrate into surface soils through crop dusting, the precipitation of vehicle exhaust, or wastewater irrigation, which might have influenced the test result. Moreover, the plough layer (usually the top 20 cm) contains increased organic matter from manure, root systems, and other humic substances, which might affect the quality of the DNA extracted, and lead to a less credible result. Therefore, an optimal depth should first be defined to distinguish oil or gas fields from nonoil gas fields.
As indicated above, it is important to determine the optimal sampling depth for prospecting oil and gas reservoirs. If the sampling depth is too shallow, numerous factors such as human activities could influence the result. However, if the depth is too deep, sampling would become a laborious process. Furthermore, as shown in Figure 6, prmA gene concentrations typically decreased with sampling depth in the oil and gas fields (except for the samples collected at 5 cm), which might be caused by the decreasing oxygen concentration with depth (Liu et al., 2014).
To avoid the disturbance of human and plant activities as much as possible, we should not select depths above the 50 cm level, as these soils would be included within the plough and cultivated layer and might further be contaminated by organic pesticide and fertilizers and root exudations (Hutsch, 1998; Seghers et al., 2005; Stiehl-Braun et al., 2011). Therefore, to minimize the economic and resource costs of sampling work and prevent prmA concentrations that are too low to detect in the gas or oil fields, we suggest 50 cm as an optimal choice for the sampling depth.
When this newly established technique is used with samples at 50 cm depth to quantify propane-oxidizing bacteria in oil or gas fields, the prmA gene is detectable, whereas it is undetectable in nonoil gas fields. The geological backgrounds and soil textures of different sites might give differing results. Therefore, in a local field in which all the soil and rock has similar characteristics, the prmA gene data might indicate whether there would be an oil or gas reservoir and where the oil or gas is most abundant. However, when the study area is across different regions with different rock and soil types, it would only be possible to determine the potential oil or gas fields from the nonoil gas fields.
To investigate a specific field, the presence of an underlying gas or oil reservoir based on the presence of the prmA gene should be examined from multiple 50 cm deep samples. Multiple samples should be taken from each site and the sampling density should follow the standard of geochemical prospecting. To accurately locate the gas or oil reservoir, geochemical and geophysical data should be combined with the prmA gene data.
The RT-PCR technique could get the more accurate results than the geochemical data. For example, the geochemical data showed that there was no propane in the soil particles sampled in Xiliu field (Table 2), but there is an oil reservoir, while the prmA gene data were consistent with the real situation. This is because the propane-oxidizing bacteria are more sensitive to propane than the laboratory apparatus used in geochemical techniques.
Conclusion
During this study, we established a robust method based on the real-time fluorescent quantitative PCR detection technology to detect the abundance of propane-oxidizing bacteria in oil and gas field soil. We considered 50 cm to be the optimal sampling depth for microbial prospecting of oil and gas reservoirs. This method can easily distinguish nonoil gas fields from those potentially containing oil or gas reservoirs.
Since these microorganisms are sensitive to hydrocarbons, the established PCR technique might detect the abnormal presence of these microorganisms in the field when gaseous hydrocarbon concentrations are below the detection limit. Compared with traditional culture-dependent microbial approaches, this technique has the advantage of saving labor, materials, and time and has more accurate results. Although we cannot identify whether the field is an oil field or a gas field through this procedure alone, the combination of the RT-PCR technique with geochemical and geophysical data could help us to do this. Therefore, the prmA RT-PCR technique has promising potential for application to oil and gas exploration.
Supplemental Material
Supplemental Material: Figures
Supplementary Figures, supplementary_material for Development of a prmA genes quantification technique and assessment of the technique’s application potential for oil and gas reservoir exploration by Zhuo Ning, Ze He, Sheng Zhang, Miying Yin, Yaci Liu and Cuiyun Zhang inEnergy Exploration & Exploitation
Footnotes
Acknowledgments
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was sponsored by China Geological Survey project (No. 1212011120268) and the National Natural Science Foundation of China (No. 41402233).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.