
Abstract
Heat shock protein 70 (HSP70) genes play essential roles in guarding plants against abiotic stresses, including heat, drought, and salt. In this study, the SlHSP70 gene family in tomatoes has been characterized using bioinformatic tools. 25 putative SlHSP70 genes in the tomato genome were found and classified into five subfamilies, with multi-subcellular localizations. Twelve pairs of gene duplications were identified, and segmental events were determined as the main factor for the gene family expansion. Based on public RNA-seq data, gene expression analysis identified the majority of genes expressed in the examined organelles. Further RNA-seq analysis and then quantitative RT-PCR validation showed that many SlHSP70 members are responsible for cellular feedback to heat, drought, and salt treatments, in which, at least five genes might be potential key players in the stress response. Our results provided a thorough overview of the SlHSP70 gene family in the tomato, which may be useful for the evolutionary and functional analysis of SlHSP70 under abiotic stress conditions.
Introduction
Climate change not only has a detrimental impact on plant survival and growth but also declines crop yield and production. 1 To adapt to unfavorable life conditions, plants have evolved a defense mechanism that enables them to grow and develop. Recent studies have shown that heat shock proteins (HSPs) are involved in responses to unfavorable environmental conditions. 2 Among them, HSP70 was one of the first and the most well-known model for adaptation study. 3 The structure of the HSP70 protein composes of a 44kDA N-terminal nucleotide-binding domain, which has ATPase activity, and a 30 kDa peptide-binding domain, (which included an 18 kDa substrate-binding domain (SBD) and a 10 kDa C-terminal lid.4–6 HSP70 was found in the majority of organelles, demonstrating the importance of this gene for survival and growth of the organism. 4
HSP70 proteins are highly conserved in a wide range of organisms, including bacteria, plants, and humans.4,7 In plants, HSP70 is resistant to abiotic elements such as extreme temperatures, drought, and salinity, as well as biotic factors such as viruses or bacteria.8–10 The expression of HSP70 genes has been recorded to be quite diverse for abiotic factors. The accumulation of HSP70 proteins in the nucleus increased heat tolerance in transgenic Arabidopsis. 11 Heat treatment of Arabidopsis plants lacking HSP70.15 protein increased plant mortality, suggesting that the HSP70.15 gene plays an important role for Arabidopsis plants in resisting heat stress. 12 Increasing expression of the BiP gene, a member of the HSP70 family, reduced the stress on the endoplasmic reticulum and improved the drought tolerance of tobacco.13,14 The mRNA content of the HSP70 gene was much higher after exposure to the cold stress of mangroves. 15 However, in other species, such as tomatoes, the roles of the HSP70 genes have not been identified.
Tomato (Solanum lycopersicum) belonged to the Solanaceae family, and is the second essential vegetable widely cultivated and consumed around the world. 16 S. lycopersicum is also an excellent model for molecular biology research of fleshy fruit because of its small genome and body size, short life cycle, and abundant seed ability. 17 With the approximate size of 900 Mb, divided into 12 chromosomes, the genome of the tomato was fully sequenced. 18 To improve tomato fruit yield, potential genes for resistance to abiotic stresses have been investigated. However, the biological functions of the tomato HSP70 (SlHSP70) genes under stress conditions are still poorly understood. Despite the response of SlHSP70 genes under metal, temperature and drought stress have begun to be addressed, the knowledge of the SlHSP70 gene family remains limited and uncomprehensive.
In this study, we identified 25 SlHSP70 genes based on public data analysis using bioinformatic methods to further analyze their evolution. The response of SlHSP70 genes under abiotic stress as heat, drought and salt stresses were analyzed. These results provided solid fundamentals for further evolutional and functional analysis of the SlHSP70 gene family.
Materials and methods
In silico identification of SlHSP70 genes in tomato
The tomato HSP70 candidate genes were searched from the Solanum lycopersicum ITAG2.4 genome on the Phytozome database. 19 All protein sequences were subjected to MOTIF Finder web tool on https://www.genome.jp/tools/motif/ for conserved HSP70 domain identification. Gene structure was analyzed and visualized using a Gene Structure Display Server 2.0 (GSDS). 20 The molecular weight (mW, kDa) and theoretical isoelectric point (pI) of the SlHSP70 proteins were analyzed using the ExPASy-ProtParam tool (https://web.expasy.org/protparam/). 21 The protein subcellular localizations were predicted using the WoLF PSORT online tool (https://wolfpsort.hgc.jp/). The protein conserved domains were characterized by MEME software (https://meme-suite.org/meme/tools/meme) with default parameters. 22
The amino acid sequences of tomato and other plants (potato, Arabidopsis, and rice, downloaded from Phytozome13 database) were aligned by ClustalW. 23 The phylogenetic analysis was evaluated using the neighbor-joining method, 24 and an unrooted phylogeny tree was constructed by MEGA 11.0.1 software, with 1000 replicates for bootstrap testing. 25
The duplication of the SlHSP70 genes was identified using the Plant Genome Duplication Database (PGDD), then the ratio of nonsynonymous substitutions number per nonsynonymous site (Ka) and the rate of synonymous substitutions number per synonymous site (Ks) of duplication genes were acquired. 26 The ratio Ka/Ks was calculated for investigating the conservation level of the gene. 27 The approximate time for the duplication event occurrences was estimated as a “million years ago’ (Mya) by formula T = Ks/2λ, in which λ = 1.5 × 10–8 substitutions per synonymous site per year was defined as the mean value of clock-like rates of synonymous substitution in tomato. 28
Promoter analysis
The 1500 bp promoter sequences from the start codon (ATG) of all SlHSP70 genes were downloaded from the Phytozome database v13.0. The cis-elements relating to stresses and hormone responses were identified by PlantCARE (http://bioinformatics.psb.-ugent.be/webtools/plantcare/html/). 29
Expression pattern of the SlHSP70 genes in different tissues
The expression profiles of SlHSP70 genes were analyzed using RNAseq data from the tomato eFP browser database (http://bar.utoronto.ca/efp_tomato/cgi-bin/efpWeb.cgi) for vegetative tissues (seedling, root, stem, leaf, vegetative meristem) and productive tissues (flower, developing fruit, and mature fruit).18,30 The relative expression levels were visualized by heat map with the log2 of FPKM value, using Graphpad prism 9.0.
Expression profiles of SlHSP70 under various stress conditions
The expression analyses of SlHSP70 genes under abiotic stresses using the public RNA-seq on GEO databases, with accession number GSE148353 for salt stress and GSE151277 for heat and drought stress. Under heat treatment, the 28-day-after-sowing plants were treated in 42°C heat stress conditions for 0 h (Heat-0 h), 2 h (Heat-2 h), and 4 h (Heat-4 h). The same-age plants were used for drought treatments by stopping watering for 0 days (Drought-0 days), 2 days (Drought-2 days), and 5 days (Drought-5 days). The tomato plants were NaCl irrigated for 0 h (Salt-0 h), 2 h (Salt-2 h), and 8 h (Salt-8 h) in the salt stress treatment.
Tomato plant growing conditions and abiotic stress treatments
The Micro-Tom tomato was used in this study. The seeds were initiated sterilized with 70% ethanol, and sown in the mix of soil and coconut coir (1:1, v/v) in a growth chamber at 24 ± 1°C with 16 h/8 h of light/dark period and watering every day with Hyponex solution (1/1000 dilution). The 28-day-old seedlings were treated with heat, drought, and salt stresses. For heat stress, the seedlings were transferred to 42°C incubator for 0, 2, and 4 h, respectively. For drought stress, watering was stopped for 2 and 5 days for the seedlings. For salt stress, the seedlings were irrigated with 250 mM NaCl for 2 and 8 h. After treatment, the seedling leaves were harvested and frozen immediately in liquid nitrogen and stored at −80°C until further analysis.
Expression analysis of the SlHSP70 genes under heat, salt, and drought stress conditions
Total RNA was extracted from all samples using IQeasy™ plus Plant RNA Extraction Kit (Intron Biotechnology, South Korean). Trace contaminated DNA was treated by DNase I (Thermo Scientific™) digestion before RNA quality testing on 1.2% Agarose gel. The RNA quantity was evaluated using NanoDrop™ 2000/2000c Spectrophotometers (Thermo Scientific™). 1 µg of total RNA was reverse transcribed into cDNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific).
0.5 µL cDNA was added to a master mix consisted 10 µL SYBR™ Green PCR Master Mix (2X) and 10 pmol each primer. The real-time PCR was performed with an initial denaturation at 95°C for 5 min, followed by 40 cycles of 95°C for 20 s, and 60°C for 60 s, using the Roche LightCycler® 96 System (Roche Molecular Systems, Inc.). The relative expression level of targeted genes was calculated using the 2−△△Ct method with UBQ3 gene was selected as a reference gene. 31 The primer sequences were listed in the Supp. Table S1. This work was carried out with three biological replicates for each sample. The relative expression level was visualized using a bar graph by Graphpad Prism 9.0.
Results
In silico identification of the SlHSP70 in tomato
To identify the SlHSP70 genes, we searched the S. lycopersicum ITAG2.4 genome, using “HSP70” as the keyword, and resulted in 25 candidate genes named from SlHSP70-1 to SlHSP70-25, based on their locus ID (Supp. Table S2), after screening the presence of HSP70 motif in their amino acid sequences (Supp. Table S3). The genomic sequences, coding sequences (CDS), and amino acid sequences were downloaded (Supp. Data 1, 2, 3, respectively). The SlHSP70 gene structure was shown in Supp. Fig. S1, and protein conserved domains were identified as in Supp. Fig. S2. The size of SlHSP70 protein ranged from 80 amino acid (aa) to 890 aa, corresponding to molecular weights in the range of 8.68–93.88 kilo Dalton (kDa). The theoretical isoelectric point values of SlHSP70 proteins varied from 4.32–9.3, but most of them below 6.0 indicated that they were considered acidic proteins. Subcellular localization prediction revealed that the SlHSP70 protein was in multi-a subcellular localization, in which two proteins could be found in the chloroplast, two proteins in mitochondria, four in the endoplasmic reticulum (ER), one in the nucleus, 12 in the cytoplasm, and four other proteins could be located in more than one organelle (Supp. Table 2).
The full-length protein sequences of SlHSP70 genes were utilized for a phylogenetic analysis into five sub-families A, B, C, D, and E, respectively (Figure 1(a)). Sub-family A was the largest group with 10 members mostly localized in the cytoplasm, followed by sub-families B (five genes localized in ER), sub-families C (four genes in mitochondrial and chloroplast), sub-families D (four genes might localize in nucleus, ER, or cytoplasm) and finally, sub-family E contained only two genes localized in cytoplasm and chloroplast.

Phylogenetic analysis of SlHSP70 based on their amino acid sequences. (a) The analysis within the SlHSP70 gene family. (b) The comparative analysis of HSP70s between tomato and potato, Arabidopsis and rice.
For further comparative genomics of the HSP70 among different plant species, another phylogenetic tree was constructed, using the HSP70 protein sequences of tomato, Arabidopsis, potato, and rice (Supp. Data 4, 5, and 6, respectively). The SlHSP70 protein was also clustered into five groups together with the HSP70 protein of potato, Arabidopsis, and rice, except group III only containing the rice HSP70s (Figure 1(b)). As expected, the SlHSP70 were closest gathered to those of potato, followed by Arabidopsis, and finally, rice, suggesting the evolutionary relationship of the HSP70s between tomato and potato (in the Solanaceae family), Arabidopsis (dicot plants) and the segregated to rice (monocot plant).
Gene duplication and evolutionary analysis
The gene duplication analysis showed that 12 duplicate events were detected between SlHSP70 genes (Figure 2). Eleven events were determined as segments while only a pair of SlHSP70-11/SlHSP70-13 was considered as tandem, suggesting segmental duplication might play a major role in the tomato SlHSP70 evolution. The most frequent duplication occurred with the gene SlHSP70-13, which was duplicated with five genes (SlHSP70-10, SlHSP70-11, SlHSP70-17, SlHSP70-19, SlHSP70-21), followed by SlHSP70-10 and SlHSP70-19 duplicated with three and two other genes, respectively. Those genes SlHSP70-12, SlHSP70-16, SlHSP70-21, and SlHSP70-23 were duplicated with only one gene. Interestingly, the duplicated genes were only found in the sub-family A and B for cytoplastic and endoplasmic reticulum proteins, suggesting the overlapped or associated biological functions of these genes in the cytoplasm and endoplasmic reticulum.
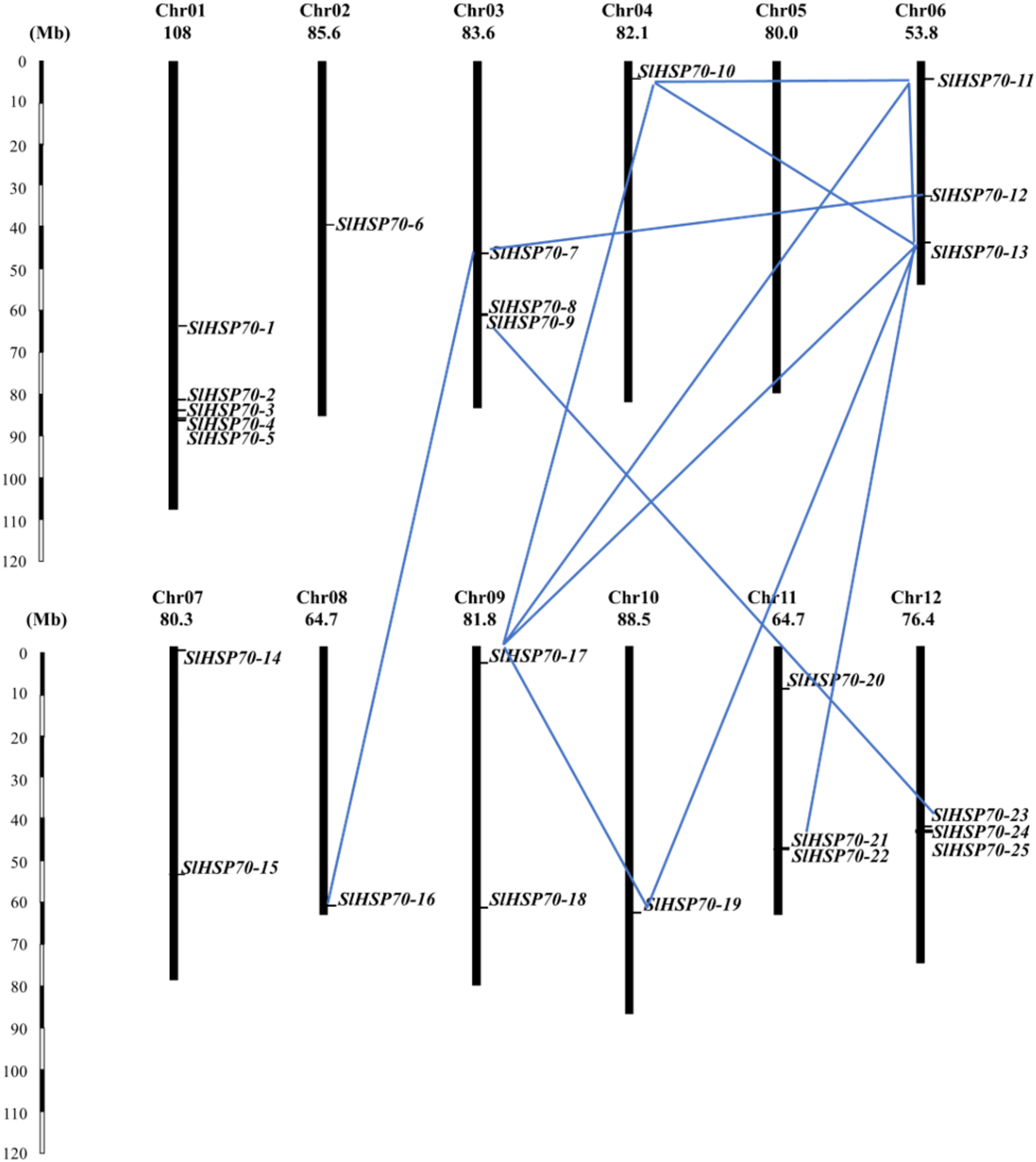
The distribution and duplication of the SlHSP70 gene family. The blue lines indicate the SlHSP70 gene duplication pairs.
The gene duplications did not occur evenly on all chromosomes. In particular, chromosome 1 carries the most member of the SlHSP70 family (five genes) however, no duplication event could be found, which was also recognized in chromosomes 2 and 7. In contrast, gene duplications were detected on chromosomes 3, 4, and on chromosomes 6 to 12, of which 2 out of 3 genes located on chromosomes 3 and 6 duplicated with other genes. The role of gene duplicated distribution and frequency might be involved in specific functions of distinct genes in the family. The Ks/Ks ratio was less than 1, implying that all segmentally duplicated genes might evolve in the stabilizing selection, relating to the high conservation of this gene family (Table 1).
Prediction of duplication events among the identified SlHSP70 genes in tomato.
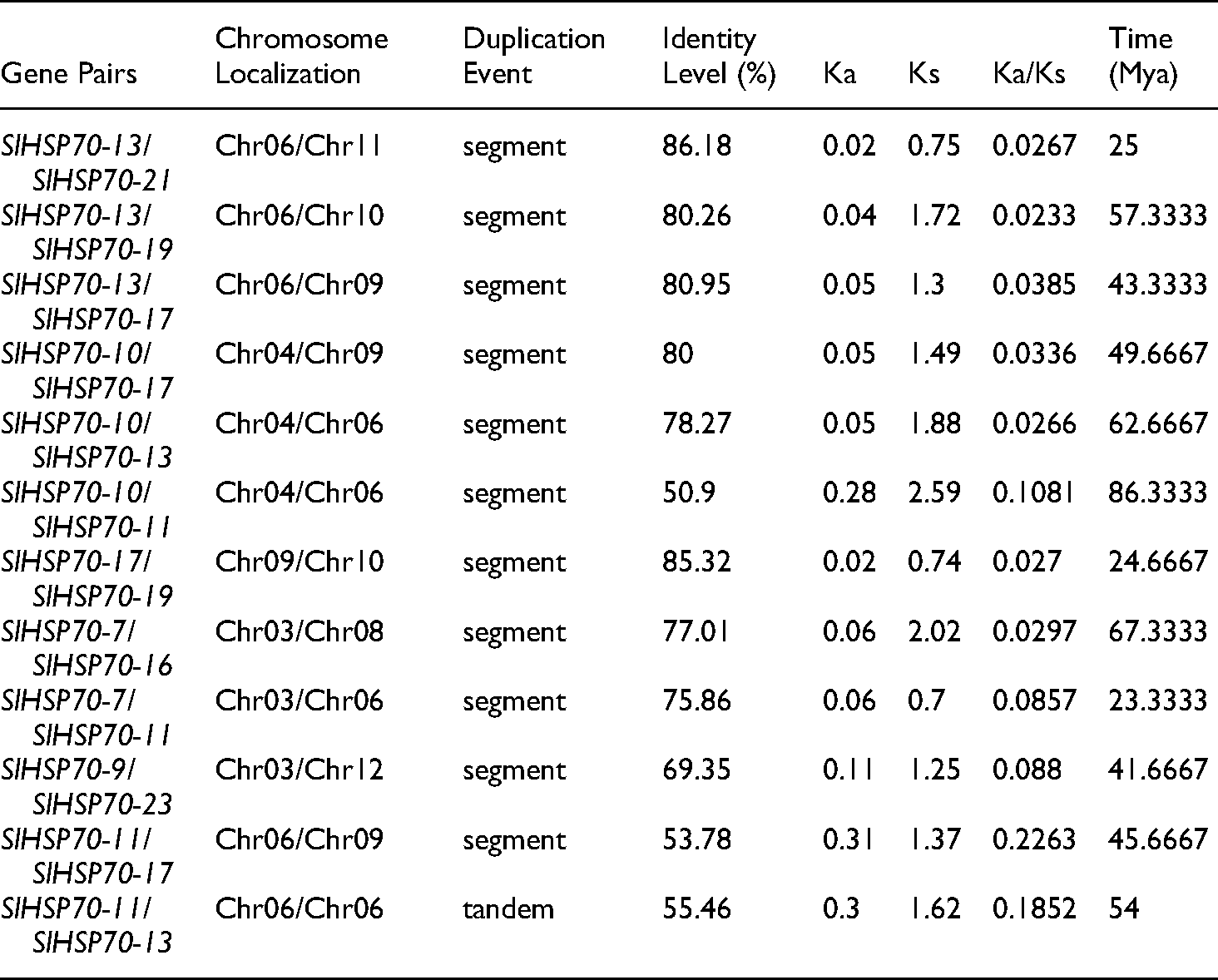
To assess the derivative analysis of gene duplication, the approximate time of each duplication event was estimated from 23.33 million to 86.33 million years ago (Mya) (Table 1). Among them, the pair of SlHSP70-10/SlHSP70-11 has the earliest duplicated event (86.3333 Mya) followed by the pair of SlHSP70-7/SlHSP70-16 (67.33333 Mya) and SlHSP70-10/SlHSP70-13 (62.66667 Mya). In contrast, the latest duplicated events were estimated from the pairs SlHSP70-7/SlHSP70-11 (23.33333 Mya), SlHSP70-17/SlHSP70-19 (24.66667 Mya) and SlHSP70-1/SlHSP70-21 (25 Mya). The time for duplication events might relate to the separation of gene functions in pairs of duplicated genes. The latest duplicated gene pairs have the highest proportion of homologous genes, which may lead to the closeness in gene functions. The earlier the duplicate gene pairs occurred, the lower the similarity rate in the genomic sequence and the greater degree of gene function segregation.
Cis-element analysis
To analyze the possible function of the SlHSP70 gene's response to abiotic and biotic stresses, 1500 bp upstream sequences of the SlHSP70 promoter regions (Suppl. Data 7) were analyzed by PlantCARE online. All potential cis-elements were identified, in which, seven cis-elements relating to abiotic stress response (LTR, HSE, ARE, STRE, DRE core, MBS, and TC-rich repeats) and other eleven relating to hormone response (ABRE, ERE, SARE, AuxRR-core, CGTCA-motif, TGACG-motif, GARE-motif, P-box, TATC-box, TCA-element, and TGA-element) were subjected and listed in Supp. Table S4. The detailed information of each cis-elements was characterized in Supp. Table S5. The total number of cis-elements in each SlHSP70 promoter ranged from 5-18, suggesting the widely involved of SlHSP70 genes in different stresses response in tomato (Figure 3).

PlantCARE analysis of different cis-elements presented in the promoter regions of the SlHSP70 genes.
The presence of cis-elements related to heat stress (HSE, LTR), and drought (MBS, DRE Core) have been found, in addition to various stress-related elements such as TC-rich repeats or STRE. The hormone-related cis-elements linked with abiotic stresses such as ABRE, which was involved in various plant tolerance to drought, high salinity, and osmotic stress, have also been documented the appearance with high frequency along with TC-rich repeats, STRE. The diverse occurrence of cis-elements in the promoter regions of all SlHSP70 genes suggested a potential feedback of SlHSP70 to a broad range of stress treatments.
Expression patterns of the SlHSP70 genes in different tissues
To comprehend the expression pattern of the SlHSP70 genes in various tomato organs, their expression levels in eight different tissues were surveyed using the public RNA-seq data (Supp. Table S6). 30 The SlHSP70-17 gene showed the highest expression levels in most tissues, followed by SlHSP70-21, SlHSP70-3, SlHSP70-24, and SlHSP70-25; while three other genes, SlHSP70-11, SlHSP70-12, and SlHSP70-23 have low expression pattern at the same organ. Gene SlHSP70-1 and SlHSP70-18 have no expression in any tissues. The expression levels of SlHSP70-4, SlHSP70-7, SlHSP70-10, SlHSP70-20, and SlHSP70-22 genes in productive tissues were higher than those in vegetative tissues and gradually increased during fruit development (Figure 4). The differentiation in the SlHSP70 genes expression might be related to their functions in various tissues. In contrast, several members expressed together in a tissue indicated their sharing biological functions might be derived from gene duplication or redundancy.

The express pattern of SlHSP70s in various tissues of tomato in normal condition. Heat map represented the log2 of FPKM value for RNA-seq data of SlHSP70 expressions from the tomato eFP browser database.
RNA-seq analysis of SlHSP70 genes expression under abiotic treatments
To understand the expression profiles of SlHSP70 genes under heat, drought, and salinity stresses, we surveyed their expressions based on reported RNA-seq data (Supp. Table S7, 8, 9). The results showed that almost SlHSP70 genes reacted to the stress treatments (Figure 5(a)–(c)). In general, the genes SlHSP70-3 and SlHSP70-17 remained highly expressed, while genes SlHSP70-1, SlHSP70-2, SlHSP70-11, SlHSP70-14, and SlHSP70-23 still kept a quite low expression under all treatments. These genes seemed to be no significant change before and after stress. The remained genes were observed to have a remarked response during stress treatment (Figure 5(a)). In detail, many genes suddenly increased after 2 h of heat treatment and then gradually decreased, such as genes SlHSP70-3, SlHSP70-9, SlHSP70-10, SlHSP70-13, SlHSP70-20, SlHSP70-21, SlHSP70-22, and SlHSP70-24 indicated that these genes tend to respond immediately to heat conditions. This trend was also observed in genes SlHSP70-7, SlHSP70-10, SlHSP70-12, SlHSP70-13, SlHSP70-19, SlHSP70-21, SlHSP70-22 under salt stress conditions. On the other hand, some other genes tended to gradually increase expression by the time of treatment, which was highlighted under drought conditions with eight genes observed (SlHSP70-4, SlHSP70-10, SlHSP70-13, SlHSP70-19, SlHSP70-20, SlHSP70-22, SlHSP70-24, and SlHSP70-25). This tendency was also present in the genes SlHSP70-7 and SlHSP70-16 under heat treatment, but absent under salt stress treatment (Figure 5(b) and (c)).

RNA-seq analysis of SlHSP70 gene family under heat, drought and salt treatments. Heat map represented the log2 of normalized FPKM value for RNA-seq data of SlHSP70s expression under heat, drought (from GEO database, accession no. GSE151277) and salt treatments (accession no. GSE148353).
In short, the gene expression analysis revealed two distinct ways the SlHSP70 genes responded to stressful conditions, responding instantaneously to heat and salt stresses, and gradually by accumulating mRNA under heat and drought treatments. Thus, the activity of SlHSP70 genes under heat and drought was more abundant than that under salt stress treatment. Several genes may play a potentially important role in abiotic stress resistance such as SlHSP70-7, SlHSP70-10, SlHSP70-13, SlHSP70-19, SlHSP70-20, SlHSP70-21, SlHSP70-22 and SlHSP70-21 due to their presence in at least two types of stress. Meanwhile, the high expression of genes SlHSP70-3 and SlHSP70-17 might be considered for their biological functions in tomato growth and stress tolerance.
It is noteworthy that pairs of duplicated genes often show a similar expression tendency under the same stress conditions. Based on RNA-seq data, pairs of SlHSP70-13/SlHSP70-21, SlHSP70-13/SlHSP70-10, SlHSP70-7/SlHSP70-16 under heat stress, SlHSP70-17/SlHSP70-19, SlHSP70-7/SlHSP70-16 under drought and pairs of SlHSP70-13/SlHSP70-21, SlHSP70-13/SlHSP70-19, SlHSP70-10/SlHSP70-17 under salt stress were found to have the same behaviour to the treatments. The similarity in the expression patterns of the duplicated genes again suggested the overlapped functions due to gene redundancy or co-expression for enhancing the gene expression of SlHSP70s and their paralogs in stress tolerance.
Expression analysis of SlHSP70 genes under heat, drought, and salt stress treatment
The real-time PCR analysis was conducted to validate the expression profiling of SlHSP70 under heat, drought, and salt stresses showed that almost SlHSP70 family members responded to abiotic stresses with different levels (Figure 6(a)–(c)). First of all, the SlHSP70 genes had strong responses to heat stress (Figure 6(a)). There were 20 members of the family that responded to heat stress while only five genes (SlHSP70-7, SlHSP70- 8, SlHSP70-9, SlHSP70-13, and SlHSP70-21) have insignificantly changed expression levels. Among responsive SlHSP70 genes, 18 members had an increase from double to 240 times expression levels. Many genes have very high levels of expression after 2 h in heat stress, such as SlHSP70-6, SlHSP70-HSP70-11, SlHSP70-17, SlHSP70-18, SlHSP70-18, SlHSP70-19, SlHSP70-20, SlHSP70-22, and SlHSP70-23, suggested their potential roles in immediate response to thermal stress. Some other genes elevated their expression level by time of heat treatment, such as SlHSP70-1, SlHSP70-3, SlHSP70-14, and SlHSP70-15 indicating their mRNA accumulation during heat stress. The elevated expression of the SlHSP70 genes suggested that these genes have a potential role to participate in the plant cells’ protective function under heat stress (Figure 6(a)).

Real-time PCR validation of SlHSP70s expression under heat (a), drought (b) and salt (c) stresses. The Bar graph with the error bar represented the Mean ± SEM of the relative expression level of targeted genes from three biological replications using UBQ3 as a reference gene. The mean expression level of SlHSP70 genes under stresses was compared to control using Student's t-test with * for P < 0.05, and ** for P < 0.01.
Contrary to the tendency of high expression for heat stress, SlHSP70 genes did not show considerable changes in expression under drought stress conditions (Figure 6(b)). Only 10 genes had exceeded double expression levels after 2 days and 5 days in drought conditions. The genes SlHSP70-1, SlHSP70-2, SlHSP70-7, and SlHSP70-9 increased expression after 2 days of drought, while SlHSP70- 13, SlHSP70-15, and SlHSP70-22 only upregulated after 5 days of drought. Three genes were keeping high expression levels after 2 days and 5 days in drought, namely SlHSP70-4, SlHSP70-5, and SlHSP70-6. Interestingly, the genes SlHSP70-12 and SlHSP70-14 downregulated during drought, suggesting that these genes may be involved in cellular physiological functions. The remaining member of the family gene did have notable changes under drought stress conditions (Figure 6(b)).
Similar to drought stress, the expression levels of SlHSP70 genes under salt stress were more stable than those under heat stress. Some genes with elevated expression such as SlHSP70-6, SlHSP70-14, SlHSP70-15, SlHSP70-16, SlHSP70-18, SlHSP70-19, SlHSP70-20, SlHSP70-22, SlHSP70-23 genes may play a role in plant resistance to salinity stress. Notably, the number of genes with reduced expression under saline stress was more than that of heat and drought stresses. Specifically, there were six genes that had remarkedly reduced expression, including genes SlHSP70-5, SlHSP70-7, SlHSP70-9, SlHSP70-10, SlHSP70-12, SlHSP70-13, and SlHSP70-21 along with two other genes SlHSP70-10 and SlHSP70-11 also tend to decreased expression. The declined expression of SlHSP70 genes may be related to their roles in tomato plant physiology under salt stress (Figure 6(c)).
Overall, among the genes with significant expression changes under heat, drought, and salt stress conditions, there were four genes that responded only to heat stress, including genes SlHSP70-1, SlHSP70-11, SlHSP70-25 with increased expression, and gene SlHSP70-10 expression decreased. Furthermore, there were 11 genes behaved simultaneously in two stress conditions, in which three genes (SlHSP70-3, SlHSP70-4, and SlHSP70-17) responded to both heat and drought conditions with elevated expression and five genes overexpressed in both heat and salinity stress, comprising SlHSP70-2, SlHSP70-16, SlHSP70-19, SlHSP70-20, and SlHSP70-23. There were three genes responded to drought and salt stress conditions, including SlHSP70-7, SlHSP70-9, and SlHSP70-13 with their up-regulated in drought and down-regulated in salt stress. Finally, there were eight genes that changed their expression patterns in all of heat, drought and salt stresses, including SlHSP70-5, SlHSP70-6, SlHSP70-12, SlHSP70-14, SlHSP70-15, SlHSP70-18, SlHSP70-22, and SlHSP70-24. Of which, SlHSP70-6, SlHSP70-15, SlHSP70-18, and SlHSP70-22 only increased, SlHSP70-12 only decreased under all conditions, while gene SlHSP70-5 decreased expressed in salt stress and gen SlHSP70-14 decreased in drought. Overall, based on the RNA-seq analysis, the genes SlHSP70-13, SlHSP70-19, SlHSP70-20, SlHSP70-22, and SlHSP70-24 might have potential functions for abiotic resistance. The different exhibitions of the SlHSP70 family members in tomato plants to heat, drought, and salinity stress conditions, suggest different roles for these genes in different treatments.
Discussion
The HSP70 gene family has been characterized in Arabidopsis, moss, rice, populus, soybeans, and other plants.32–36 However, their roles in tomato still have not been examined. Therefore, a comprehensive study of tomato SlHSP70 was performed using a genome-wide approach for their evolution analysis through gene duplication, promoter analysis, and gene expression profiling under abiotic (heat, drought, salt) stresses based on public data. In this study, the size of the tomato HSP70 family (25 genes) was larger than that in Spinach (12 genes), Arabidopsis (18 genes), and moss (21 genes), but it was smaller than that in rice (32 OsHSP70 members) or cotton.34–39 In the Solanaceae, the SlHSP70 family size was also bigger than that in potato (20 genes) and pepper (21 genes) but smaller than the 117 genes size of the eggplant HSP70 gene family.40–42 The large difference in the number of HSP70 genes in monocotyledonous and dicotyledonous plants might be derived from gene redundancy; and their functions may be overlapped or distinct, depending on their abundant interactions with specific co-chaperones.43,44
The distribution of HSP70 genes in different tissues was found previously. 45 In our study, the SlHSP70 proteins were predicted to be localized in all organelles, including the nucleus, cytoplasm, and mitochondrial membranes. The presence of SlHSP70 in all subcellular organelles might suggest their essential roles in cellular defense against stress conditions due to the direct link between protein localization and its proposed biological function. 46
The duplications might derive from a part of the gene, or take up the whole gene, with various mechanisms, expression, regulation, and biological function, potentially played important role in the evolution of plant genome structures. 47 In this study, the tandem and segments 48 were investigated, in which, segmental events were determined in most of the predicted duplications, indicating that this type of event was critical for expansion of the SlHSP70 gene family. The duplicated genes clustered together in the phylogeny tree indicated their closed protein sequences. Similarities in gene functions can also be inferred by their structural homology because of the intertwining between protein structures and their biological roles. 49 The alteration in gene structure might change gene function in the gene evolution over time under any selective pressure. 50 The direction of evolution and the selective pressures were evaluated by the ratio (Ka/Ks) of duplicated gene pairs. 27 In this study, the Ka/Ks ratios of all duplication events were below 1, strongly suggesting how essential functions the SlHSP70s needed to maintain stress resistance.
The role of the HSP70 genes in response to heat, drought, and salinity has been suggested in previous studies. 51 The enhanced transcription of HSP70s under salt and drought stress indicated the involvement of specific HSP70s to counteract abiotic stresses. 52 In cotton, at least one HSP70 gene was found to respond to cold, salt, and heat stress, and its transcription level was positively correlated with drought tolerance. 53 In Arabidopsis, the individual HSP70 not only participated in the response to abiotic stresses of plants, but also performed both redundant and specific functions with the other family members. 54
In tomato, the HSP70 played critical roles in maintaining the cellular homeostasis of plants for drought adaptation and plant growth under normal conditions.55,56 The increased transcriptional level of the HSP70 gene was also found in the overexpression of SIZ1 SUMO E3 (SlSIZ1), a post-translational modification enzyme in tomato at high temperatures to enhance tolerance to heat stress, although the role of the HSP70 in this study has not been elucidated. 57 Hahn et al. (2011) proposed a model with flexible crosstalk between HSP90, and HSP70 and in controlling heat stress factors (Hsfs) for heat stress response, but which specific HSP70 member joined in the HSP90 interaction was still unknown. 58 In this study, we found that not only one but several members of the SlHSP70 family are involved in abiotic stresses. Specifically, at least five members of the SlHSP70 gene family were mentioned because of their responsiveness to the stress response shown by RNA-seq and real-time PCR analysis. The diverse occurrence of cis-elements might take part in the response of these genes to stress, as suggested in previous studies.59,60 Gene duplication might play an important role in genome expansion during evolution, probably as a key factor in enhancing the response of genes to stress factors. 61
Conclusion
In this study, a total of 25 putative SlHSP70 genes were identified, and their structural features and distribution in the genome were characterized, using bioinformatic methods. The gene duplications in the SlHSP70 family were found, in which segmental events were determined to have critical contributions to the family gene expansion. The expression pattern of SlHSP70s in various tissues revealed their participation in tomato growth and development. Their expression profile eight genes in the SlHSP70 gene family are responsive to all heat, drought and salt treatments, as well as 18 members reacted to two stresses demonstrates the wide involvement of SlHSP70 genes in abiotics stress responses. Five SlHSP70 genes were recommended as potential candidates for playing key roles in abiotic stress resistance.
Our research has provided basic understandings of SlHSP70 characteristics and their involvement in heat, drought, and salt stress treatments. These findings might establish the fundamentals for further analysis of evolutionary as well as elucidation of potential biological functions of the SlHSP70 genes in abiotic stress resistance.
Supplemental Material
sj-xlsx-1-sci-10.1177_00368504221148843 - Supplemental material for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum)
Supplemental material, sj-xlsx-1-sci-10.1177_00368504221148843 for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum) by Nam Tuan Vu, Ngoc Bich Thi Nguyen, Hanh Hong Ha, Linh Nhat Nguyen, Ly Han Luu, Ha Quang Dao, Trinh Thi Vu, Hue Thu Thi Huynh and Hien Thu Thi Le in Science Progress
Supplemental Material
sj-pptx-2-sci-10.1177_00368504221148843 - Supplemental material for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum)
Supplemental material, sj-pptx-2-sci-10.1177_00368504221148843 for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum) by Nam Tuan Vu, Ngoc Bich Thi Nguyen, Hanh Hong Ha, Linh Nhat Nguyen, Ly Han Luu, Ha Quang Dao, Trinh Thi Vu, Hue Thu Thi Huynh and Hien Thu Thi Le in Science Progress
Supplemental Material
sj-docx-3-sci-10.1177_00368504221148843 - Supplemental material for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum)
Supplemental material, sj-docx-3-sci-10.1177_00368504221148843 for Evolutionary analysis and expression profiling of the HSP70 gene family in response to abiotic stresses in tomato (Solanum lycopersicum) by Nam Tuan Vu, Ngoc Bich Thi Nguyen, Hanh Hong Ha, Linh Nhat Nguyen, Ly Han Luu, Ha Quang Dao, Trinh Thi Vu, Hue Thu Thi Huynh and Hien Thu Thi Le in Science Progress
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship,and/or publication of this article: This research is funded by the Graduate University of Science and Technology under grant number GUST.STS.ĐT2020-SH08.
Supplemental material
Supplemental material for this article is available online.
Author biographies
Nam Tuan Vu is a researcher at the Vietnam Academy of Science and Technology (VAST). He obtained a doctorate in Agriculture from the Faculty of Life Sciences, University of Tsukuba (Japan) in 2019, and then did a post-doctoral fellow at Seoul National University (South Korea) from 2020. His research interests include applying molecular biology for plant genetic resources conservation, improving crop yields and quality; using genetic engineering for purification of active compounds from plants for pharmaceutical applications, increasing the content, and enhancing the activity of potential medicinal and pharmaceutical applications compounds.
Ngoc Bich Thi Nguyen is currently working as a research assistant at Institute of Genome Research (VAST). She has just got a Bachelor in Biotechnology from Vietnam National University of Agriculture. Her research interest is plant genetics.
Hanh Hong Ha is currently a researcher at Institute of Genome Research (VAST). She obtained a PhD in Genetics from Institute of Biotechnology (VAST) in 2016. Her research field is plant genetics.
Linh Nhat Nguyen holds a MSc in Experimental Biology from Hanoi University of Science in 2018. His area of research is molecular biology, plant barcoding, and plant genetics.
Ly Han Luu holds a MSc in Experimental Biology from Hanoi University of Science in 2016. Her area of research is molecular biology, plant cell culture, and plant genetics.
Ha Quang Dao is a research assistants at Institute of Genome Research (VAST). He is currently master students at Graduate University of Science and Technology. His area of research is plant genetics.
Trinh Thi Vu is a research assistants at Institute of Genome Research (VAST). She is currently master students at Graduate University of Science and Technology. Her area of research is plant genetics.
Hue Thu Thi Huynh is a researcher at Institute of Genome Research (VAST) and a visiting lecturer at the Graduate University of Science and Technology. She obtained a PhD in Biochemistry from Institute of Biotechnology (VAST) in 2012. Her research field is plant genetics.
Hien Thu Thi Le is currently a senior researcher and Head of Genome Diversity Laboratory, Institute of Genome Research (VAST). She also teaches courses at the Graduate University of Science and Technology. She earned her PhD in Plant biology from the Institute of Biotechnology in 2003. Her research field is Plant genetic, conservation and sustainable uses of plant genetic resources.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.