
Abstract
Pogostemon Desf. includes a wide range of taxa found in subtropical and tropical areas. Few works, however, have studied microanatomical characteristics of Pogostemon species and as yet identified features of Pogostemon auricularius (L.) Hassk. Thus, in this paper, we examined the taxonomic implications of root, stem and leaf morphology for species P. auricularius collected from Quang Tri Province. Light microscopy was mainly used in our study. Qualitative characters like stem quadrangular, hirsute; leaves opposite, ovate, margin serrulate; calyx campanulate; corolla small with separate equal lobes have been found in P. auricularius. Epidermal anatomy on the aerial parts of the species like epidermal cell shape, anticlinal walls, trichomes types, stomata types and calcium oxalate crystals types were examined as well. Quantitative characters like the length and width of leaf blade and inflorescence; the size of oil droplet, stomata and calcium oxalate crystals measured provided taxonomic significance. Based on identifying morphological characteristics of P. auricularius, we aimed to contribute to the taxonomic investigation into the genus Pogostemon and give relative morphological and microanatomical features compared with other taxa.
Introduction
Pogostemon Desf. is the most ecologically and economically large genus of the family Lamiaceae, with roughly 94 species. 1 It is mainly distributed in Southern and Southeast Asia, whilst a few may be found in China, Africa and Australia. 2 Pogostemon species were found in diverse habitats, but mainly focused on aquatic/marshland or terrestrial regions. In Asian countries, Pogostemon has been widely used in traditional and folk medicine to treat abdominal pain, diarrhea, stomachaches, rheumatism, colds, headaches, fevers, and cutaneous wounds. 3 Pogostemon has had a complicated evolutionary background, bringing in difficulties in taxonomy identification. According to the latest study, the infrageneric classification split Pogostemon into three subgenuses: Allopogostemon Bhatti & Ingr., Pogostemon sensu Bhatti & Ingr. and Dysophyllus (Blume) Bhatti & Ingr., based on inflorescence morphology. However, the phylogenetic distribution of taxa formed on secondary calyx venation and calyx size is similar to that formed on habitats. 2 In particular, Allopogostemon possesses inflorescence with one lateral spike, calyx more than 4 mm long, calyx 10-veined. In contrast, the inflorescence of Pogostemon has more than two lateral spikes, calyx more than 4 mm long, calyx 5-veined and multiple secondary veins. Dysophyllus has calyx less than 4 mm long, broadly conical and 5-veined. 4 Microscopic techniques were used in previous studies to add new species records, plant systematics, anatomical, micro, and macro morphological examinations, and to identify differences in medicinal plant variations.5,6 Foliar epidermal features, along with types of trichomes diverse among the genera Pogostemon considered to play an important role in identifying species taxon.7–9 Specific studies of morphological features have been carried out on the genus Pogostemon.10–13
Pogostemon auricularius (L.) Hassk., also called Dysophylla auricularia (L.) Blume, Bijdr, 14 is a tropical and subtropical annual herb found in India, Sri Lanka, Bangladesh, China, and Southeast Asia15,16 P. auricularius is used in traditional medicine to treat stomachaches, kidney pain, digestive disorders, and malaria. 17 In addition, some studies have shown that extracts of P. auricularius have antibacterial, antifungal, analgesic, anti-inflammatory, anti-diarrheal, and α-amylase enzyme-inhibiting effects.18,19 Our previous studies of the biological and phytochemical characteristics of P. auricularius collected in Quang Tri province reported that isolated meroterpenoids, triterpenes, and phloroglucinol derivatives demonstrate moderate or strong inhibition of certain cancer cell lines such as the human colon cancer cell line (SW480), epidermal carcinoma cell line (KB), gastric cancer cell line (AGS), liver tumor cell line (HepG2), and human lung cancer cell line (LU-1).20–24
In Vietnam, P. auricularius morphology is described in the Flora of Vietnam as having the following characteristics: stem 0.4–2 m tall, prostrate basally; apex branching; petiole rarely to 1.2 cm; leaf blade oblong, yellow strigose, with scattered impressed glands, base rounded; flower continuous or occasionally interrupted at base; calyx long as corolla; stamens exserted sections bearded; nutlets brown, subglobose.15–17 However, little has been anatomically and micromorphologically done on the species P. auricularius except a previous study on the structure and function of glandular trichomes. Peltate trichomes and short-stalked capitate trichomes were found on leaves, leaf stalks, and stems of P. auricularius, and they all came from a single epidermal cell, similar to glandular trichomes in other Lamiaceae species. The histochemical findings revealed terpenes, polysaccharides, flavones, and lipids in the gland's head region, recommending secondary metabolic biosynthesis and the secretory process. 10 Therefore, this present study was designed with aims of providing relative information on morphological and micromorphological features by light microscopy techniques in order to clarify P. auricularius and to strengthen the systematics of Pogostemon.
Materials and methods
Materials
The aerial parts of P. auricularius were collected in Quang Tri Province, Viet Nam (N16°44′38.9″ E107°14′51.1″) in May 2017.
Methods
Scientific name identification
A voucher specimen (PA-01) was deposited at the Faculty of Pharmacy, Hue University of Medicine and Pharmacy, Viet Nam. The plant was verified by analyzing morphological features and reproductive parts and comparing those characteristics to those identified in a plant library template and relevant literature, including Takhtajan's classification, Flora of Vietnam, and Flora of China.16,17,25 Plant identification was confirmed by Dr Nguyen The Cuong, Institute of Ecology and Biological Resources, VAST, Viet Nam.
Plant identification characteristics
Powder characteristics
Leaves of the plant were washed, dried, and finely ground into a flour that was sifted using the appropriate size mesh. The powder was placed on slides with several drops of distilled water. The sections were covered with glass, and the leaf characteristics were viewed under an optical microscope (Eclipse E100, Nikon, Japan) and photographed with camera (Nikon, D5100).26–28
Micro-morphology
Fresh or dried leaves were softened and cut with a razor blade or microsurgical instrument to provide thin sections. The sections were soaked in 5% sodium hypochlorite for approximately 20 min to remove specific cell substances. Several times, sections were washed with water, immersed in 1% acetic acid for two minutes, and rinsed with water. The sections were then dyed with a 1% methylene blue solution for approximately 15–30 s and quickly rinsed with water. Sections were stained with 10% carmine red for approximately 30 min and washed several times with water. The sections were placed on a drop of 10% glycerol on glass slides, and a cover glass was applied. The sections were observed using light microscopy to photograph, and the micro-morphology characteristics were recorded.13,29–31
Botanical characteristics of samples were described and compared to plant taxonomic documents and stored templates at the Institute of Ecology and Biological Resources, VAST, with the assistance of scientific botany specialists. Cutting, bleaching, and double staining methods were used in conjunction with the electron microscope and digital microscope to observe, photograph, and study the microscopic characteristics of P. auricularius.
Results and discussion
Morphological characteristics
The plant is an annual herb that crawls and probably grows straight to a height of 40–100 cm. Stems are frequently quadrangular and have a hairy surface. Petiole 5–12 mm long, higher stem leaves subsessile; leaf blade ovate-oblong, 2.5–7 × 1.5–3 cm, pilose on both sides, especially near veins; leaf base rounded or obtuse, sometimes wedge-shaped, border serrated, apex obtuse to acute, five or six paired lateral veins, noticeable abaxially (Figure 1). Thyrsoid inflorescence apex, approximately 3–12 cm long and in bloom, approximately 1 cm in diameter. Bracts ovate, approximately 2 × 1–1.2 mm, hairy. Flowers are nearly subsessile. Calyx campanulate is 1 mm long, upper split into lobes, pentafid, margin pilose. Corolla white or purplish, approximately 3 mm long, glabrous or hairy externally, apex split into four equal lobes. Stamens four, attached to the corolla, approximately 3 mm long, lower pilose, upper glabrous. Glamorous ovary and style, approximately 4 mm, bifid (Figure 2).

Morphological features of the vegetative organs of P. auricularius. (A) Flowering branch; (B) Stem with trichomes; (C) Transverse section of the stem; (D) Leaves; (E) The abaxial surface; (F) The adaxial surface.

Morphological features of the reproductive organs of P. auricularius. (A) Inflorescence; (B) Inflorescence in part; (C) Floret; (D) Calyx; (E) Corolla; (F) Stamen; (G) Carpel.
Light microscopy
Root anatomy
The root of P. auricularius (Figure 3) had a circular cross-section, and the layers from outer to inner including the phellem (arrow 1) that consisted of 2–3 layers of rectangular cells arranged adjacently. The cortex (arrow 3) comprised several layers of parenchyma polygonal, large-sized, thin-walled, and loosely arranged; with spherical oil droplets inside (arrow 2). Secondary phloem (arrow 5) and xylem occupied the largest area, showing cells oriented radially in concentric circles. The outer layer of phloem had smaller cells and darker colors than parenchyma cells, along with the presence of blue-stained, thick-walled bast fibers (arrow 4) that were spatially separated or arranged in clusters. The secondary xylem (arrow 6) included blue-stained, large-sized, thick-walled xylem vessels interspersed with xylem parenchyma and small-sized, thick-walled, clustered fibers (arrow 7).
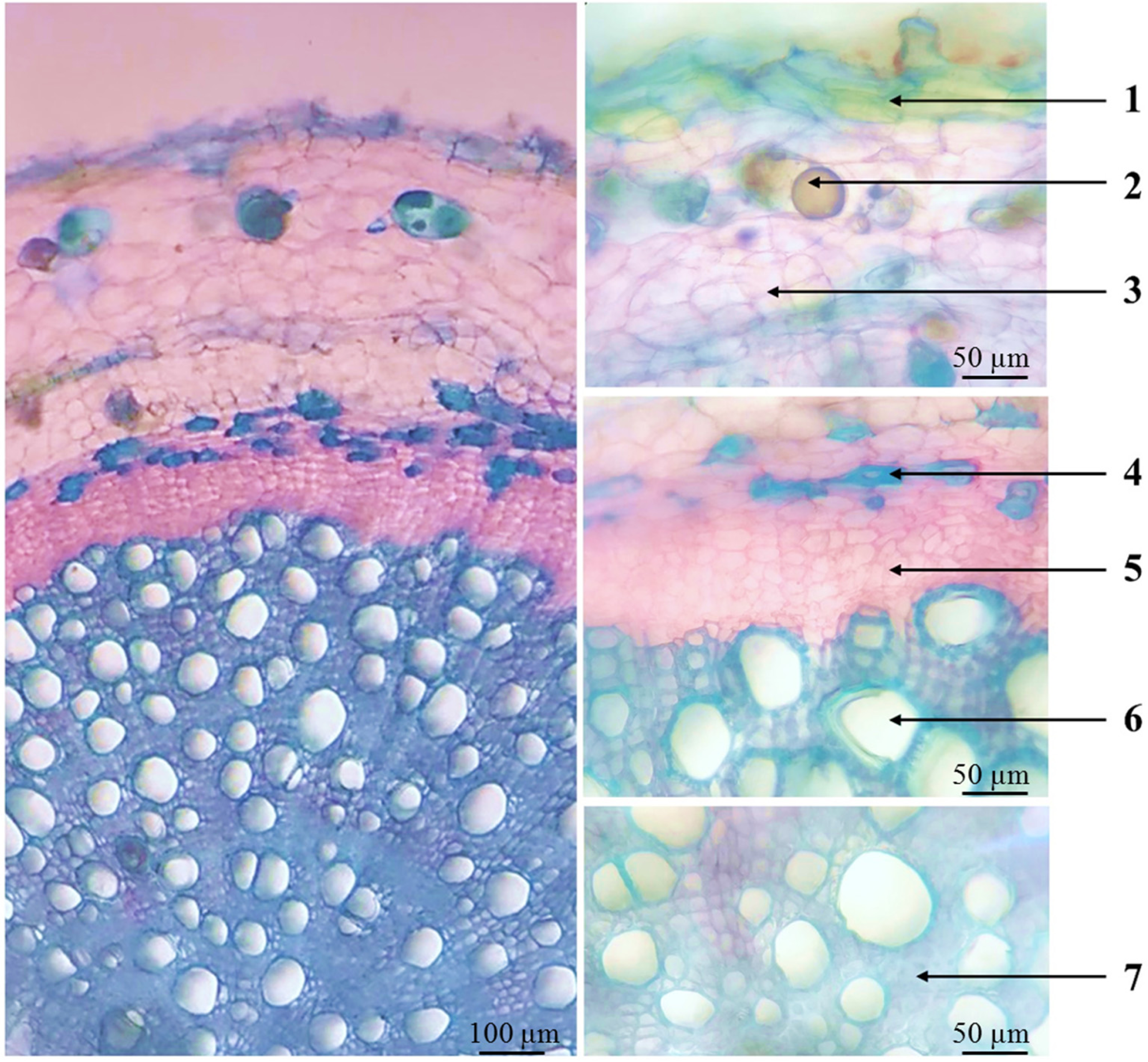
Microscopic characteristic of roots cross-sections of P. auricularius. Note: 1. Phellem; 2. Oil droplet; 3 Cortex; 4. Phloem fibers; 5. Secondary phloem; 6. Secondary xylem; 7. Xylem fibers.
Stem anatomy
The transverse section of the stem of P. auricularius (Figure 4) was square-shaped, with layers from outer to inner including epidermis (arrow 1), that was unlayered and composed of small, pentagonal-shaped to hexagonal shaped, adjacent cells; a thick outermost wall having uniseriate multicellular non-glandular trichomes with apical cone-shaped cell (arrow 2). Capitate glandular trichomes with multicellular spherical heads (arrow 3) were scattered on the surface. Within the secretory head of the trichome, oil droplets may be seen. Collenchyma (arrow 4) adjacent to the epidermis consisted of polygonal cells in 6–8 layers, with thickened walls at the corners, magenta-colored. Cortical parenchyma (arrow 5) comprised multiple layers of thin-walled, randomly organized polygonal cells larger than collenchyma cells. Secondary phloem and xylem were not divided into bundles but were annular-shaped. The secondary phloem (arrow 7) located externally consisted of polygonal cells, smaller in size and stained darker pink than parenchyma cells. The outer phloem contained phloem fibers (arrow 6), oval-shaped, thick-walled, blue-stained, either isolated or clustered, appearing in four corners of the microanatomical specimen. The secondary xylem (8) inward consisted of large xylem vessels arranged in a radial pattern; at the end, primary xylem vessels (arrow 9) were gradually smaller in size. Pith parenchyma (arrow 10) comprised many polygonal cells, thin-walled, large-sized, randomly arranged, occupying most of the micro-anatomical cross-section.

Microscopic characteristic of stems cross-sections of P. auricularius. Note: 1. Epidermis; 2. Non-glandular trichomes; 3. Glandular trichomes; 4. Collenchyma; 5. Cortical parenchyma; 6. Phloem fibers; 7. Secondary phloem; 8. Secondary xylem; 9. Primary xylem; 10. Pith parenchyma.
Leaf anatomy
The midrib was biconvex in cross section, with convexity on both adaxial and abaxial side. The lower epidermis (arrow 2) separated into oval lobes consisted of one layer of small, isodiametric cells that were organized adjacently. The anticlinal cell walls were straight to curved. The outermost wall was thickened and covered with long, large multicellular non-glandular trichomes (arrow 1) in a cross-section of the midrib. Lower collenchyma (arrow 3) consisted of 3–4 layers of cells isodiametric, large, wall-thickened and magenta-stained. More inferior parenchyma (arrow 4) comprised polygonal cells in 5–7 layers, large-sized, thin-walled, and arranged randomly. The central phloem–xylem was divided into two small bundles, including downward phloem (arrow 5) composed of 3–4 layers of the small, dark pink cell. Xylem (arrow 6), located upward that consisted of large and lined-up hollow cells, with blue-colored walls thickened and lignified. The upper parenchyma (arrow 7), upper collenchyma (arrow 8), and upper epidermis (arrow 9) had a similar structure to the upper. The surface of the upper epidermis showed short-stalked capitate glandular trichomes with oil droplet inside (arrow 10) and non-glandular trichomes partly unicellular (arrow 11).
In the leaf blade section, the epidermis cells on both sides had straight and thin anticlinal walls (arrow a, e). It presented 2–3 layers of spongy parenchyma (arrow c) and palisade parenchyma (arrow d) as well. The palisade and spongy layers were clearly separated in the mesophyll. The peltate trichome was partially sunken into the lower epidermis (arrow b) (Figure 5).

Microscopic characteristic of leaves cross-sections of P. auricularius. Note: A. Leaf midrib: 1. Multicellular non-glandular trichomes; 2. Lower epidermis; 3. Lower collenchyma; 4. Lower parenchyma; 5. Phloem; 6. Xylem; 7. Upper parenchyma; 8. Upper collenchyma; 9. Upper epidermis; 10. Oil droplet; 11. Non-glandular trichome. B. Leaf blade: a. Lower epidermis; b. Peltate trichome c. Spongy parenchyma; d. Palisade parenchyma; e. Upper epidermis.
Powder microscopy
Microscopic characteristics of powdered root
The brown powder (Figure 6) had brown pieces of phellem (1a, 1b) that were large-sized, rectangular cells in a straight line. Fragmented parenchyma consisted of polygonal cells, thin-walled, loosely arranged, with many small starch granules inside. Calcium oxalate crystals were styloid (3a, 3b), small-sized in the range of 15–25 µm. Long, thick-walled fibers adjacently form bundles (4a, 4b). Fragments of pitted vessels (5a, 5b) appeared in the microscopic area. Spherical drops of essential oil (6) with a diameter of approximately 25–35 µm can be observed occasionally.

Microscopic features of root powder. Note: 1a, 1b. Fragments of phellem; 2a, 2b. Fragments of parenchyma; 3a, 3b. Calcium oxalate crystals; 4a. Fiber bundle in vertical section; 4b. Fiber bundle in transverse section; 5a, 5b. Fragments of pitted vessels; 6. Oil droplet.
Microscopic characteristics of powdered stem
When observed under a microscope, the grayish-green powdered stem had multicellular non-glandular trichome (1a, 1b) that were large and appeared densely in a microscopic area. The short-stalked capitate glandular trichome (2) occurred separately. Pieces of pitted vessels (3a, 3b) and spiral vessels (4a, 4b) were present. Calcium oxalate crystals were styloid, including elongated square dipyramids (5a) or very elongated square dipyramids (5b), and approximately 15–25 µm long. Oil droplets (6a, 6b) were spherical, yellow colored, and 25–35 µm long. Fibers (7) were oblong, thick-walled, with small cytoplasm. Fragmented parenchyma (8) consisted of large polygonal cells, with thin walls, in a random arrangement (Figure 7).
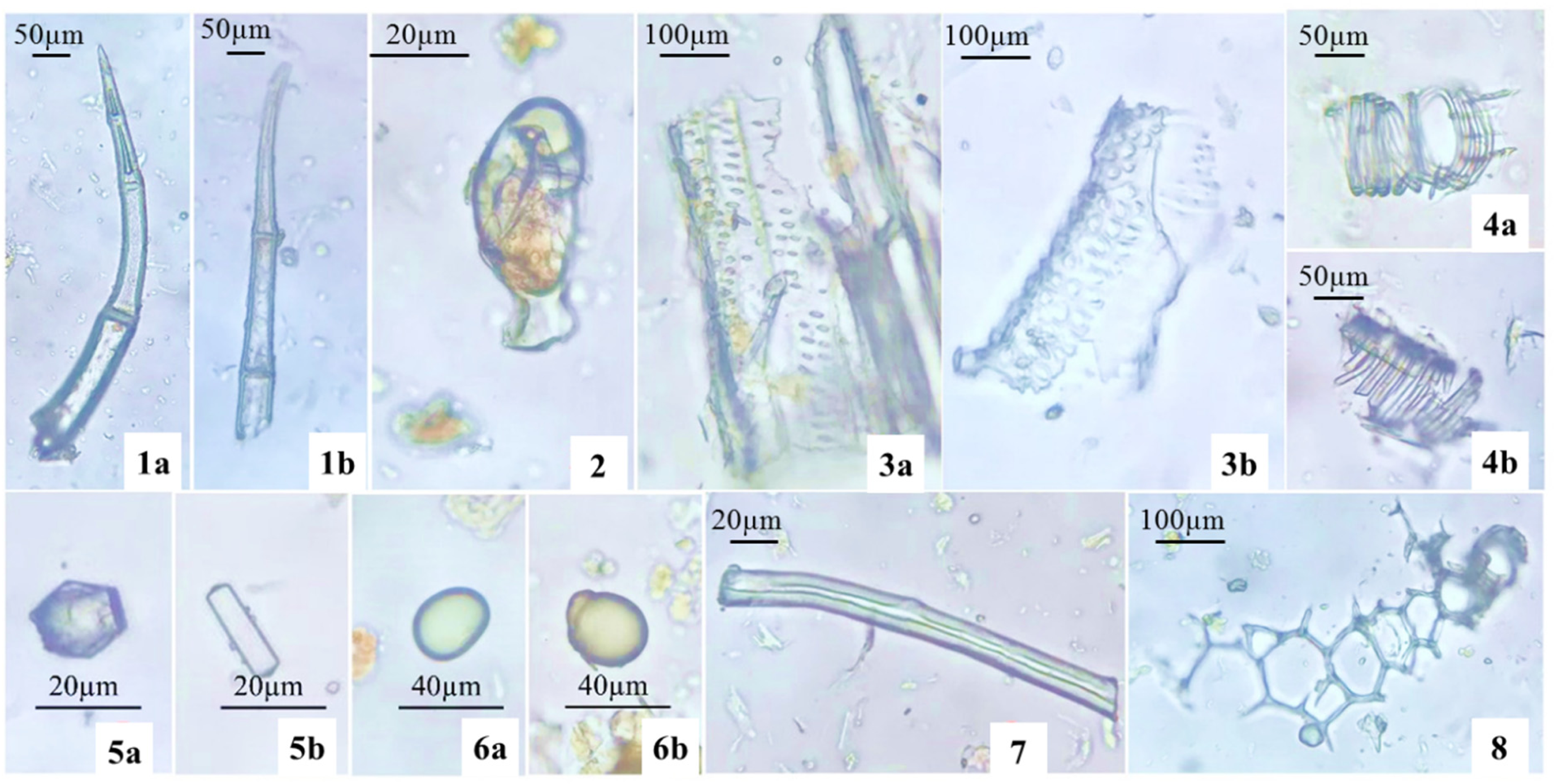
Microscopic features of stem powder. Note: 1a, 1b. Multicellular non-glandular trichome; 2. Capitate glandular trichome; 3a, 3b. Fragments of pitted vessels; 4a, 4b. Spiral vessels; 5a, 5b. Calcium oxalate crystals; 6a. 6b. Oil droplets; 7. Fiber; 8. Fragment of parenchyma.
Microscopic characteristics of powdered leaf
The dark greenish powdered leaf had several microscopic characteristics: Large multicellular non-glandular trichome (1) frequently appeared under microscopy. Capitate glandular trichome (2a) had three parts: a multicellular stalk, a short neck cell and a unicellular head. Peltate glandular trichome bearing one basal cell, a short-stalked cell and a spherical head (2b) was found on leaf microscopy. Fragments of the epidermis containing stomata (3) were rarely observed; small stomata in the range of 15 × 20 µm consisted of 2 smaller guard cells. Starch granules (4a, 4b) were circular, with well-defined umbilical points, approximately 10–25 µm in diameter, isolated or clustered in 2–3 granules. Fragmented collenchyma (5) comprised thick-walled, adjacent cells, thickening walls at the corners. Spiral vessel pieces were scattered (6a, 6b). Calcium oxalate crystals came in druse and styloid (7) that were approximately 15–25 µm long (Figure 8).

Microscopic features of leaf powder. Note: 1. Multicellular non-glandular trichome; 2a. Capitate glandular trichome 2b. Peltate glandular trichome; 3. Stomata on epidermis; 4a, 4b. Starch granules; 5. Fragment of collenchyma; 6a, 6b. Fragment of the spiral vessel; 7. Calcium oxalate crystals.
Discussion
Pogostemon plant morphologies are various, ranging from terrestrial subshrubs to aquatic herbs, and the presence of extended stamens with moniliform hairs is a frequent feature of this genus. 32 Previously, numerous taxonomists divided the genus into two divisions based on habitat,33,34 with Pogostemon s.s. being the terrestrial group and Dysophylla El Gazzar & L. Watson ex Airy Shaw being the aquatic/marshland group. 14 The terrestrial group usually has opposing petiolate leaves with broad leaves that are coated in hairs, whereas the aquatic/marshland group usually has verticillate sessile leaves with a linear to filiform lamina shape and are normally glabrous. 14 The relationship between Pogostemon and Dysophylla Blume (1826: 826) has been questioned in the past; P. auricularius (D. auricularia Blume), in instance, has opposite leaves and appears to be intermediate between the two genera.4,35 Pogostemon is strongly supported to be monophyletic, according to a recent molecular phylogenetic analysis, and all Dysophylla species sampled are nested within Pogostemon. 36 Hasskarl (1842) was the first to combine Dysophylla with Pogostemon, owing to their hairy stamens in common. 37 Following that, Bhatti and Ingrouille (1997) revised the infrageneric classification of Pogostemon Delf. and divided the genus into three subgenera: Allopogostemon Bhatti & Ingr., Dysophyllus (Blume) Bhatti & Ingr., and Pogostemon sensu Bhatti & Ingr., using a total of 135 morphological character states.2,38 P. auricularius possessed morphological traits were featured as opposite leaves with blade ovate-oblong and having pilose on both sides; flowers are nearly subsessile (calyx campanulate is 1 mm long, corolla white or purplish, approximately 3 mm long).13,39 It had stamens four that was approximately 3 mm long and lower pilose. According to the infrageneric taxonomy, P. auricularius appeared to belong to the subgenus. Dysophylla, sect. Dysophylla and have similar characters to those observed in studies on Pogostemon glabratus, Pogostemon quadrifolius, Pogostemon myosuroides, Pogostemon micangensis.4,32 In Viet Nam, there are 17 species has been found that belong to the Pogostemon. The results of morphological characters of P. auricularius has been described in specific detail in order to prevent harvesting and buying counterfeit herbal medicines.
Leaf microscopy of P. auricularius was used to explore the leaf micromorphological characteristics which are typical of the Lamiaceae such as the curved outward line of the midrib, the presence of capitate and peltate trichomes and the phloem-xylem bundles covered up around the parenchyma. 13 In particular, foliar epidermal cells were adaxially and abaxially isodiametric, and having anticlinal cell walls straight to curved and not undulated. Previous work has proved that habitats of species contributed to the undulation of anticlinal cell walls. 40 Epidermal cells with straight or curved anticlinal cell walls were identified in abundance in species that grew in arid environments, which were reasonable for P. auricularius collected from montane forests and coniferous forests. 41 Both non-glandular and glandular trichomes were observed on the surface of stem and leaf. In this study, non-glandular trichomes were identified as uni- or multicellular uniseriate trichomes. Simple unicellular non-glandular trichomes had smooth cell wall surfaces, whereas cuticular micro-papillae covered the wall surfaces of simple multicellular non-glandular trichomes. 42 Glandular trichomes seen in morphology and powder microscopy were classified into two main types: peltate and capitate. According to a previous study on P. auricularius, a basal cell was embedded between the epidermal cells, one short unicellular stalk cell, and a big head cell made up peltate trichomes. 10 The essential oils, which were the major component in this species,22–24 were produced mostly by the peltate trichomes.43,44 Capitate trichomes had two subtypes: short-stalked capitate and long-stalked capitate which were consistent with the result of previous finding on P. auricularius. 10 The stalk length, the neck cell, and the shape of the glandular head are the variations between these trichomes. The capitate trichomes with those characteristics have also been seen in various investigations on Pogostemon cablin, Leonotis leonorus and Plectranthus ornatus.12,45,46 Capitate trichomes may have diverse traits, reflecting different secretory mechanisms, and would most likely serve different purposes. Compared to the peltate trichomes, the capitate trichomes were observed abundantly and densely. Lamiaceae was known for having short-stalked capitate trichomes. 46 At the generic level, however, unicellular peltate trichomes were taxonomically unique in Pogostemon. 47 The stomatal occurrence was hypostomatic. Diacytic stomatal types were commonly found. In addition, calcium oxalate crystals belonged to two types of shape and structure, druse and styloid in particular which were also found in other species of Lamiaceae, including Oxera robusta, Ballota nigra, Leucas abyssinica, Meriandra dianthera and Hemiandra pungens. 48
Conclusion
This study provides the first detailed description of the morphological, microanatomical, and powder microscopic characteristics of P. auricularius. This result is a valuable database contributing to the taxonomic identification of P. auricularius. However, glandular trichomes, stomata and crystals are typical of the delimitation of the genus Pogostemon which were not presented in detail and completeness using light microscopy. Further study suggested that analysing secretory mechanism, in-depth micromorphology of stomata and crystal of P. auricularius compared with other Pogostemon species by using other techniques like scanning electron microscopy (SEM) and transmission electron microscopy (TEM).
Footnotes
Acknowledgements
This research was supported by Hue University of Medicine and Pharmacy, Hue University.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.